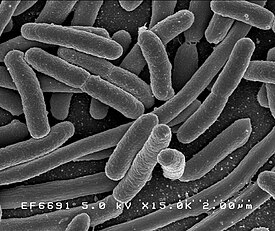
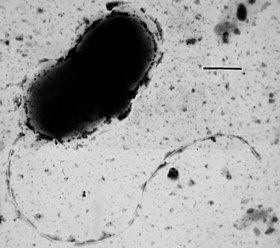
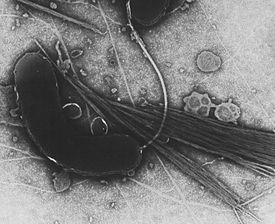
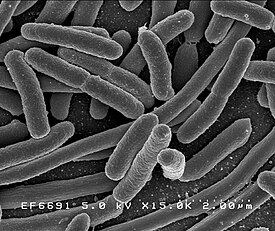
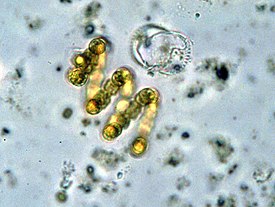
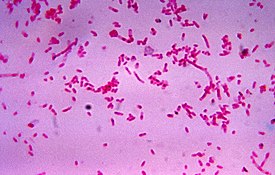
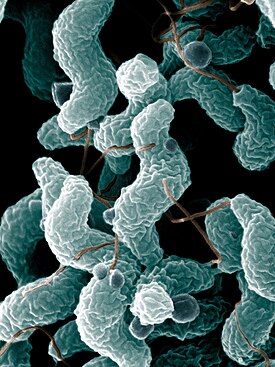
Грамположительные бактерии
- 1 year ago
- 0
- 0
Бакте́рии ( лат. Bacteria , от др.-греч. βακτήριον — «палочка») — домен прокариотических микроорганизмов . Бактерии обычно достигают нескольких микрометров в длину, их клетки могут иметь разнообразную форму: от шарообразной до палочковидной и спиралевидной. Бактерии — одна из первых форм жизни на Земле и встречаются почти во всех земных местообитаниях. Они населяют почву , пресные и морские водоёмы, кислые горячие источники , радиоактивные отходы и глубинные слои земной коры . Бактерии часто являются симбионтами и паразитами растений и животных . Большинство бактерий к настоящему времени не описано, и представители лишь половины отделов бактерий могут быть выращены в лаборатории . Бактерии изучает наука бактериология — раздел микробиологии .
Один грамм почвы в среднем содержит 40 миллионов бактериальных клеток, а в миллилитре свежей воды можно найти миллион клеток бактерий. На Земле насчитывается около 5⋅10 30 бактерий , и их биомасса превышает суммарную биомассу животных и растений . Они играют важную роль в , например, именно бактерии осуществляют фиксацию атмосферного азота . Они также разлагают останки животных и растений посредством гниения . Экстремофильные бактерии, обитающие рядом с и горячими гидротермальными источниками , вырабатывают энергию из нерастворимых соединений, таких как сероводород и метан . Предполагается, что бактерии живут и в Марианской впадине , имеющей глубину 11 километров . Имеются сообщения о бактериях, обитающих в каменистых породах на 580 метров глубже морского дна на глубине 2,6 км около северо-востока США .
Человеческую микрофлору составляют 39 триллионов бактериальных клеток (само тело человека состоит из около 30 триллионов клеток) . Наиболее многочисленна кишечная микрофлора . Кожа также заселена многими бактериями . Большинство бактерий, обитающих в человеческом теле, безвредно за счёт сдерживающего действия иммунной системы или приносят пользу ( микрофлора человека ). Ряд бактерий патогенен для человека. Такие инфекционные болезни , как холера , сифилис , сибирская язва , проказа и бубонная чума , вызываются бактериями. Наибольшее число смертей вызвано бактериальными , и один лишь туберкулёз ежегодно убивает 2 миллиона человек (преимущественно в Африке южнее Сахары ) . В развитых странах антибиотики используются не только для лечения заболеваний человека, но и в животноводстве , из-за чего проблема устойчивости к антибиотикам становится всё более актуальной. В промышленности бактерии используют в очистке сточных вод , для ликвидации разливов нефти , при получении сыра и йогурта , восстановлении золота , палладия , меди и других металлов из руд , а также в биотехнологии , для получения антибиотиков и других соединений .
Первоначально бактерии поместили в царство растений в составе класса Schizomycetes . Сейчас известно, что бактерии, в отличие от растений и других эукариот , не имеют оформленного ядра и, как правило, мембранных органелл . Традиционно бактериями называли всех прокариот, однако в 1970-х годах было показано, что прокариоты представлены двумя независимыми доменами — бактериями и археями (эукариоты составляют третий домен) .
Слово «бактерия» происходит от лат. bacterium , производного от др.-греч. βακτήριον , что означает «палочка», так как первые описанные бактерии были палочковидными .
Предки современных бактерий были одноклеточными микроорганизмами, которые стали одной из первых форм жизни на Земле, появившись около 4 миллиардов лет назад. Почти три миллиарда лет вся жизнь на Земле была микроскопической . Хотя для бактерий известны ископаемые останки (например, строматолиты ), их морфология очень однообразна, что не позволяет идентифицировать отдельные виды . Однако для реконструкции филогении бактерий можно использовать последовательности генов , и именно с их помощью было показано, что бактерии отделились раньше архей и эукариот . Ближайший общий предок бактерий и архей, вероятнее всего, был гипертермофилом , который жил 3—2,5 млрд лет назад .
Бактерии сыграли важнейшую роль в появлении эукариот. Считается, что эукариотическая клетка возникла, когда бактерии стали эндосимбионтами одноклеточных организмов, вероятно, близких к современным археям . Иными словами, прото-эукариотическая клетка проглотила клетку α-протеобактерии , которая дала начало митохондриям и гидрогеносомам . На данный момент неизвестны эукариоты, лишённые и митохондрий, и гидрогеносом, хотя иногда эти органеллы сильно редуцированы. Впоследствии некоторые из эукариот, уже имеющих митохондрии, проглотили клетки цианобактерий , которые стали пластидами растений и водорослей .
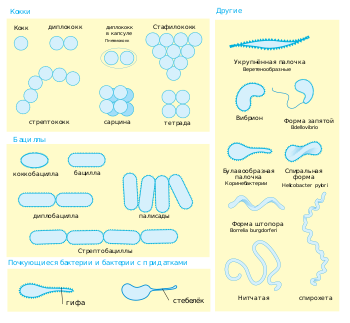
Бактериальные клетки имеют чрезвычайно разнообразную морфологию (то есть форму и размер). Как правило, бактериальные клетки в десять раз мельче эукариотических и достигают 0,5—5 мкм в длину. При этом есть и бактерии, видимые невооружённым глазом: так, Thiomargarita namibiensis достигает половины миллиметра в длину , а длина Epulopiscium fishelsoni может составлять 0,7 мм . Один из видов бактерий вырастает до 2 см в длину, отличается необычно сложной клеткой, фактически стирая границу между прокариотами и эукариотами и, по одному из предположений, даже является недостающим звеном в эволюции живого от одноклеточных организмов . К числу самых мелких бактерий можно отнести представителей рода Mycoplasma , длина клеток которых не превышает 0,3 мкм , что сравнимо по размерам с вирионами некоторых вирусов и лишь немного превышает предел разрешающей способности оптического микроскопа . Существуют ещё более мелкие бактерии ( ), однако они плохо изучены .
Большинство бактерий имеют шарообразную ( кокки ) или палочковидную ( бациллы ) форму . Некоторые бактерии, называемые , выглядят как слегка закрученные палочки или запятые; спириллы имеют спиральную форму, а спирохеты имеют длинные плотно закрученные клетки. Описаны и бактерии с другими необычными формами клеток, например, клетками в форме звезды . Разнообразие форм бактериальных клеток обусловлено особенностями их клеточных стенок и цитоскелета . Форма бактериальной клетки обусловливает их способность поглощать питательные вещества , прикрепляться к поверхностям, плавать в жидкостях и ускользать от питающихся бактериями организмов .

Многие виды бактерий существуют в виде одиночных клеток, однако у некоторых видов клетки образуют характерные скопления: например, клетки Neisseria объединены в пары, у Streptococcus — в цепочки, у Staphylococcus — в скопления в виде грозди винограда. Некоторые бактерии могут формировать более сложные многоклеточные структуры. Так, Actinobacteria формируют длинные филаменты (внутриклеточные нитевидные образования), Myxococcales образуют плодовые тела , а Streptomyces образуют ветвящиеся нити . Иногда такие сложные структуры появляются только при некоторых условиях. Например, при нехватке аминокислот клетки Myxococcales определяют расположение соседних клеток того же вида с помощью чувства кворума , движутся навстречу друг другу и формируют плодовые тела до 500 мкм длиной, состоящие из около 100 тысяч бактериальных клеток . Бактериальные клетки в составе плодовых тел выполняют различные функции: десятая часть всех клеток мигрирует к верхней части плодового тела и превращается в особую покоящуюся форму, называемую миксоспорой, которая более устойчива к высыханию и другим неблагоприятным условиям внешней среды .
Бактерии часто прикрепляются к какой-либо поверхности и формируют плотные скопления, известные как биоплёнки , или более крупные скопления — бактериальные маты . Толщина биоплёнок и матов может составлять от нескольких микрометров до полу метра , в их состав могут входить бактерии разных видов, а также археи и протисты . В биоплёнках наблюдается сложное расположение клеток и внеклеточных компонентов, которые формируют вторичные структуры, известные как микроколонии, через которые проходит сеть каналов, обеспечивающая лучшую диффузию питательных веществ . В таких местообитаниях, как почва и поверхность растений, большинство бактерий, прикреплённых к поверхностям, входят в состав биоплёнок . Биоплёнки имеют важное значение для медицины, потому что они часто образуются при хронических бактериальных инфекциях или инфекциях , связанных с инородными имплантатами. Более того, бактерии в составе биоплёнок гораздо сложнее убить, чем отдельные бактериальные клетки .

Бактериальная клетка окружена мембраной , состоящей в основном из фосфолипидов . Мембрана окружает всё содержимое клетки и выступает в роли барьера для удержания в клетке питательных веществ, белков и других компонентов цитоплазмы . В отличие от клеток эукариот, у бактерий, как правило, отсутствуют крупные мембранные органеллы, такие как ядро, митохондрии, хлоропласты . Однако у некоторых бактерий имеются органеллы с белковой оболочкой, в которых протекают определённые метаболические процессы , например, карбоксисомы . Кроме того, у бактерий имеется многокомпонентный цитоскелет, который контролирует локализацию нуклеиновых кислот и белков внутри клетки и управляет клеточным делением .
Многие важные биохимические реакции , такие как образование АТФ , происходят за счёт градиента концентрации определённых ионов по разные стороны мембраны, что создаёт разность потенциалов , как в батарейке . Поскольку у бактерий нет мембранных органелл, такие реакции (например, перенос электронов ) протекают при участии мембраны бактериальной клетки, обращённой во внешнюю среду в случае грамположительных бактерий или в периплазматическое пространство в случае грамотрицательных бактерий . Однако у многих фотосинтезирующих бактерий мембрана образует многочисленные складки, которые заполняют почти всё внутреннее пространство клетки . На этих складках располагаются светопоглощающие комплексы, однако у некоторых бактерий, например, зелёных серных бактерий , светопоглощающие комплексы находятся внутри особых мембранных пузырьков — хлоросом .
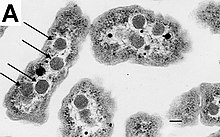
У большинства бактерий нет ядра, окружённого мембранами, и их генетический материал , в большинстве случаев представленный единственной , находится в цитоплазме в составе нуклеоида , имеющего неправильную форму . Нуклеоид содержит не только геномную ДНК, но также взаимодействующие с ней белки и РНК . Как все живые организмы, бактерии имеют рибосомы , которые обеспечивают синтез белков , однако размеры и структура рибосом бактерий отличаются от таковой у рибосом архей и эукариот .
У некоторых бактерий в цитоплазме имеются гранулы, запасающие питательные вещества, такие как гликоген , полифосфат , сера или полигидроксиалканоаты . Ряд бактерий, например, фотосинтезирующие цианобактерии, имеют газовые вакуоли , с помощью которых они регулируют свою плавучесть, благодаря чему могут перемещаться между слоями воды с разным содержанием питательных веществ и уровнем освещённости .
Поверх мембраны бактериальной клетки располагается клеточная стенка. Клеточная стенка бактерий состоит из пептидогликана , также известного как муреин, который состоит из полисахаридных цепочек, связанных пептидными линкерами из D -аминокислот . По химическому составу бактериальная клеточная стенка отличается от клеточной стенки растений и грибов , у которых она состоит из целлюлозы и хитина соответственно . Клеточная стенка архей также не содержит пептидогликана. Клеточная стенка жизненно необходима для многих видов бактерий, и некоторые антибиотики, такие как пенициллин , подавляют биосинтез пептидогликана и тем самым убивают бактерию .
В широком смысле по составу клеточной стенки бактерий принято делить на грамположительные и грамотрицательные . Название этих типов связано с их дифференциальной окраской по методу Грама , который долгое время используется для классификации бактерий . У грамположительных бактерий имеется толстая клеточная стенка, состоящая из многих слоёв пептидогликана и тейхоевых кислот . У грамотрицательных бактерий, напротив, клеточная стенка значительно тоньше и включает всего лишь несколько слоёв пептидогликана, а поверх неё залегает вторая мембрана , содержащая липополисахариды и липопротеины . Большинство бактерий грамотрицательны, и только фирмикуты и актинобактерии грамположительны (ранее они были известны как грамположительные бактерии с низким GC-составом и грамположительные бактерии с высоким GC-составом соответственно) . Различия между грамположительными и грамотрицательными бактериями могут обусловливать различную чувствительность к антибиотикам. Например, ванкомицин эффективен только против грамположительных бактерий и не действует на грамотрицательные бактерии . У некоторых бактерий строение клеточной стенки не соответствует в строгом смысле ни грамположительному, ни грамотрицательному типу. Например, у микобактерий имеется толстый слой пептидогликана, как у грамположительных бактерий, который покрыт внешней мембраной, как у грамотрицательных бактерий .
У многих бактерий клетка покрыта так называемым S-слоем , состоящим из плотно уложенных молекул белков . S-слой обеспечивает химическую и физическую защиту клетки и может выступать в роли макромолекулярного диффузионного барьера . Функции S-слоя разнообразны, но плохо изучены, однако известно, что у Campylobacter он выступает фактором вирулентности , а у он содержит поверхностные ферменты .

У многих бактерий имеются жгутики , представляющие собой плотные белковые структуры около 20 нм в диаметре и до 20 мкм в длину. Они обеспечивают подвижность клеток и по строению и механизму работы не имеют ничего общего с эукариотическими жгутиками. Движение жгутиков бактерий происходит за счёт энергии, которая высвобождается при движении ионов по электрохимическому градиенту через клеточную мембрану .
Нередко клетки бактерий покрыты фимбриями , которые представляют собой белковые филаменты, достигающие 2—10 нм в диаметре и до нескольких мкм в длину. Они покрывают всю поверхность бактериальной клетки и в электронный микроскоп выглядят как волоски. Предполагается, что фимбрии участвуют в прикреплении клеток бактерий к различным поверхностям и друг к другу, а у многих патогенных бактерий они являются факторами вирулентности . Пили — это клеточные белковые придатки, более толстые, чем фимбрии, которые обеспечивают перенос генетического материала от одной бактериальной клетки к другой в ходе (половые пили) . Кроме того, пили IV типа участвуют в движении .
Многие бактериальные клетки выделяют покрывающий их гликокаликс различной сложности строения: от тонкого неструктурированного слоя до высоко структурированной капсулы . Гликокаликс может защищать бактерию от поглощения эукариотическими клетками, например, макрофагами , входящими в состав иммунной системы . Он также может выступать в роли антигена , который используется для распознавания бактериальных клеток иммунной системой, а также участвовать в формировании биоплёнок и прикреплении бактериальных клеток к поверхностям .
Образование внеклеточных структур бактериальной клетки обеспечивается бактериальными системами секреции . Они транспортируют белки из цитоплазмы в периплазматическое пространство или во внешнюю среду. Известно несколько типов бактериальных систем секреции, кроме того, бактериальные системы секреции нередко выступают в роли факторов вирулентности .

Представители нескольких родов грамположительных бактерий, таких как , Clostridium , , и , образуют покоящиеся структуры, обладающие повышенной устойчивостью к неблагоприятным факторам внешней среды и называемые эндоспорами . Эндоспоры образуются в цитоплазме клетки, и, как правило, в одной клетке может сформироваться только одна эндоспора. Каждая эндоспора содержит ДНК и рибосомы, окружённые поверхностным слоем цитоплазмы, поверх которого залегает плотная многослойная оболочка, состоящая из пептидогликана и разнообразных белков .
Внутри эндоспор не протекают метаболические процессы, и они могут выживать при сильнейших неблагоприятных физических и химических воздействиях, таких как интенсивное УФ-излучение , γ-излучение , детергенты , дезинфицирующие агенты, замораживание, давление и . Эндоспоры могут сохранять жизнеспособность в течение миллионов лет , и с их помощью бактерии могут оставаться живыми даже в условиях вакуума и космического излучения . Некоторые бактерии, формирующие эндоспоры, патогенны. Так, сибирская язва развивается после вдыхания спор грамположительной бактерии Bacillus anthracis , а попадание эндоспор Clostridium tetani в глубокие открытые раны может привести к столбняку .
У бактерий наблюдается колоссальное разнообразие видов метаболизма . Традиционно таксономия бактерий строилась на основе их метаболических особенностей, однако она во многом не совпадает с современной классификацией, построенной на геномных последовательностях . Бактерии делятся на три типа питания в зависимости от ключевых черт метаболизма: источника энергии, донора электронов и источника углерода .
Бактерии получают энергию двумя способами: поглощая свет в ходе фотосинтеза или окисляя химические соединения ( хемосинтез ) . Хемотрофы используют в качестве источника энергии химические вещества, перенося электроны с имеющегося донора на конечный акцептор электронов в ходе окислительно-восстановительной реакции . Высвобождающаяся при этой реакции энергия далее используется для нужд метаболизма. В зависимости от того, какое вещество используется как донор электронов, хемотрофы подразделяются ещё на несколько групп. Бактерии, использующие неорганические вещества , такие как водород , угарный газ или аммиак , называются литотрофами , а бактерии, окисляющие органические соединения , называются органотрофами . Бактерий также классифицируют в зависимости от веществ, выступающих акцепторами электронов. У аэробов акцептором электронов выступает кислород , а анаэробы используют для этого другие соединения, такие как нитрат , сульфат и углекислый газ .
Многие бактерии удовлетворяют свои потребности в углероде за счёт органических соединений; такие бактерии называются гетеротрофами . Другие бактерии, например, цианобактерии и некоторые пурпурные бактерии , являются автотрофами , то есть получают углерод, фиксируя углекислый газ . В некоторых условиях метанотрофные бактерии используют метан и как источник электронов, и как источник углерода .
| Тип питания | Источник энергии | Источник углерода | Примеры |
|---|---|---|---|
| Фототрофы | Солнечный свет | Органические вещества ( фотогетеротрофы ) или фиксированный углекислый газ (фотоавтотрофы) | Цианобактерии, зелёные серные бактерии, , пурпурные бактерии |
| Литотрофы | Неорганические соединения | Органические вещества (литогетеротрофы) или фиксированный углекислый газ ( литоавтотрофы ) | , , |
| Органотрофы | Органические соединения | Органические вещества (хемогетеротрофы) или фиксированный углекислый газ (хемоавтотрофы) | Bacillus , Clostridium , Enterobacteriaceae |
Метаболизм бактерий имеет огромное значение для экологической стабильности и деятельности человека. Например, некоторые бактерии являются единственными фиксаторами атмосферного азота (с помощью фермента нитрогеназы ) . Другими важными для окружающей среды химическими процессами, осуществляемыми бактериями, являются денитрификация , восстановление сульфата и ацетогенез . Метаболические процессы бактерий также могут служить источниками загрязнения. Так, сульфатредуцирующие бактерии образуют высокотоксичные соединения ртути ( метил - и диметилртуть ) . Ряд анаэробных бактерий осуществляет брожение для получения энергии, и его побочные продукты (например, этанол при спиртовом брожении ) попадают в окружающую среду. Факультативные анаэробы могут переключаться между получением энергии с помощью брожения и получением её с помощью дыхания с различными акцепторами электронов в зависимости от условий окружающей среды .

В отличие от многоклеточных организмов , у одноклеточных организмов, в том числе бактерий, увеличение клетки в размерах и размножение путём деления клеток тесно связаны . Бактериальные клетки достигают определённого размера и после этого делятся бинарным делением. В оптимальных условиях бактерии растут и делятся очень быстро, описан пример морской псевдомонады , популяция которой может удваиваться каждые 9,8 минуты . При бинарном делении образуются две дочерние клетки, идентичные материнской. Некоторые бактерии, хотя и размножаются простым делением, образуют более сложные структуры, предназначенные для распространения дочерних клеток. Примером могут служить плодовые тела миксобактерий и воздушные гифы стрептомицетов . Некоторые бактерии способны к почкованию , когда дочерняя клетка образует вырост на материнской, который впоследствии отделяется и переходит к самостоятельной жизни .

В лаборатории бактерий выращивают на твёрдых или жидких средах . Твёрдые среды, такие как агар , используются для чистых культур бактериальных штаммов . Жидкие среды используются, когда необходимо измерять скорость роста или получить большое количество клеток. При выращивании бактерий в жидкой среде с перемешиванием получаются однородные клеточные культуры, однако сложно заметить загрязнение другими бактериями. Для идентификации отдельных бактерий используются селективные среды, содержащие антибиотики, специфические питательные вещества или, наоборот, лишённые каких-то соединений .
Для большинства лабораторных методов выращивания бактерий необходимы большие количества питательных веществ, чтобы обеспечить быстрое получение больших объёмов клеток. Однако в естественных условиях питательные вещества ограничены, и бактерии не могут размножаться бесконечно. Из-за ограниченного количества питательных веществ в ходе эволюции появились различные стратегии роста. Некоторые виды растут чрезвычайно быстро, когда питательные вещества доступны, например, цианобактерии нередко вызывают цветение водоёмов , насыщенных органикой . Другие организмы адаптированы к жёстким условиям окружающей среды, например, бактерии рода Streptomyces выделяют антибиотики, которые подавляют рост конкурирующих бактерий . В природе многие виды бактерий живут сообществами (например, в виде биоплёнок ), которые обеспечивают каждую клетку необходимым питанием и защищают от неблагоприятных условий . Некоторые организмы и группы организмов растут только в составе сообществ и не могут быть выделены в чистую культуру .
Динамику роста бактериальной популяции можно подразделить на четыре фазы. Когда популяция бактерий попадает в среду, богатую питательными веществами, клетки начинают адаптироваться к новым условиям. Первая фаза роста называется лаг-фазой, это период медленного роста, когда клетки адаптируются к среде, богатой питательными веществами, и готовятся к быстрому росту. Во время лаг-фазы происходит интенсивный синтез белков . За лаг-фазой следует логарифмическая , или экспоненциальная фаза, во время которой происходит быстрый экспоненциальный рост . Скорость, с которой клетки растут во время этой фазы, называют скоростью роста, а время, которое необходимо для удвоения клеточной популяции, называется временем генерации. В ходе лог-фазы питательные вещества потребляются с максимальной скоростью, до тех пор пока одно из необходимых соединений не кончится и не начнёт подавлять рост. Третья фаза роста называется стационарной, она начинается при нехватке питательных веществ для быстрого роста. Скорость метаболизма падает, и клетки начинают расщеплять белки, не являющиеся строго необходимыми. Во время стационарной фазы экспрессируются гены, белковые продукты которых участвуют в репарации ДНК , метаболизме антиоксидантов и транспорте питательных веществ . Финальная фаза роста — фаза смерти, при которой запас питательных веществ исчерпывается и бактерии погибают .
У большинства бактерий геном представлен единственной кольцевой молекулой ДНК (её иногда называют хромосомой ), а размер генома варьирует от 160 тысяч пар оснований (п. о.) у эндосимбиотической бактерии до примерно 13 миллионов п. о. у почвенной бактерии . Впрочем, у ряда представителей родов Streptomyces и Borrelia геном представлен единственной линейной хромосомой , а у некоторых видов рода Vibrio имеется более одной хромосомы . Многие бактерии также содержат плазмиды — маленькие внехромосомные молекулы ДНК, которые содержат несколько генов, обеспечивающих своих обладателей разными полезными свойствами: устойчивостью к антибиотикам, новыми метаболическими реакциями и разнообразными факторами вирулентности .
В бактериальных геномах, как правило, содержится от нескольких сотен до нескольких тысяч генов. Бактериальные гены, за редкими исключениями, лишены интронов , а если они и есть, то очень короткие .
При делении бактериальной клетки дочерние клетки наследуют идентичные копии её генома и, по сути, являются её клонами . Однако в геномах бактерий постоянно происходят мутации , лучшие из которых поддерживаются отбором , кроме того, иногда у бактерий происходит и генетическая рекомбинация . Мутации возникают из-за ошибок ферментов, удваивающих ДНК , а также под действием мутагенов . Скорость мутирования значительно различается не только у бактерий разных видов, но даже и у разных клонов, относящихся к одному виду . Изменения в бактериальных геномах происходят также благодаря случайным мутациям и так называемым стресс-направленным мутациям (то есть гены, отвечающие за какой-либо процесс, сдерживающий рост, мутируют особенно часто) .
У некоторых бактерий клетки могут обмениваться друг с другом генетическим материалом. Существует три способа обмена генетической информацией между бактериями. Во-первых, бактериальные клетки могут поглощать экзогенную ДНК из окружающей среды в ходе процесса, называемого трансформацией . Некоторые бактерии поглощают ДНК извне в нормальном состоянии, а другие начинают захватывать чужеродную ДНК после химического воздействия, то есть они должны прежде стать . В природе компетентность развивается при стрессовых условиях и выступает в роли адаптивного механизма, потому что захваченный извне фрагмент ДНК может оказаться полезным для переживания неблагоприятных условий . Во-вторых, бактерия может получить чужеродную ДНК в ходе трансдукции , когда бактериофаг не только сам вставляется в бактериальную хромосому, но и приносит с собой фрагмент генома другой бактерии. Существует множество бактериофагов, но для всех них существует два типа жизненного цикла: литический цикл , когда клетка-хозяин разрушается вскоре после заражения, высвобождая наружу новые вирусные частицы, и лизогенный цикл , когда бактериофаг встраивается в геном бактерии и до некоторого момента мирно сосуществует с ней . У бактерий существует ряд механизмов защиты от бактериофагов, в частности, система рестрикции-модификации и система CRISPR /Cas. Система CRISPR/Cas, по сути, играет роль адаптивного иммунитета , потому что фрагменты геномов фагов, заразивших клетку, встраиваются в локус CRISPR, и при повторном заражении их репликация подавляется за счёт РНК-интерференции . В-третьих, бактериальные клетки обмениваются генетическим материалом в ходе конъюгации, когда ДНК передаётся из одной клетки в другую при непосредственном контакте. Обычно передача генетического материала любым из трёх механизмов подразумевает участие бактерий одного вида, однако в некоторых случаях ДНК обмениваются бактерии разных видов. Например, гены устойчивости к антибиотикам передаются от одних видов бактерий к другим . Передача генетического материала между разными особями носит название горизонтального переноса генов (в противовес вертикальному переносу генов, то есть от родителей к потомкам) .

Многие бактерии подвижны и перемещаются за счёт разных механизмов. Чаще всего для движения используются жгутики — длинные филаменты, которые вращаются, подобно пропеллеру , за счёт особого мотора у их основания. Движущей силой для мотора является электрохимический градиент клеточной мембраны . В состав бактериального жгутика входят около 20 белков, а ещё 30 белков необходимы для регуляции его работы и сборки .

С помощью жгутиков бактерии могут двигаться по-разному. Многие бактерии, например, кишечная палочка Escherichia coli , могут двигаться вперёд или кувыркаться. За счёт кувыркания клетка меняет направление своего движения, которое представляет собой случайное блуждание в трёхмерном пространстве . Количество и расположение жгутиков различно у разных видов. У некоторых бактерий есть всего один жгутик (монотрихи), у других два жгутика располагаются на двух противоположных концах клетки (амфитрихи), у третьих на полюсах клетки находятся пучки жгутиков (лофотрихи), а у четвёртых жгутики покрывают всю поверхность клетки (перитрихи). У спирохет жгутик находится в периплазматическом пространстве между двумя мембранами. Клетки спирохет имеют характерную извитую форму, которая меняется при движении .
Некоторые бактерии способны к так называемым ( англ. twitching ) за счёт пилей IV типа , а также скольжению . При подёргивании на клетке имеется палочковидный пиль, который связывается с субстратом и сокращается, проталкивая клетку вперёд .
Для подвижных бактерий характерно движение, направленное навстречу какому-либо стимулу или, напротив, от него — таксис . К числу таких поведенческих программ относятся хемотаксис , фототаксис , энергетический таксис и магнитотаксис . Клетки бактерий могут кооперироваться с образованием единого скопления за счёт чувства кворума, как, например, миксобактерии при образовании плодовых тел. Некоторые виды родов Listeria и Shigella движутся внутри клетки-хозяина, используя её цитоскелет , который обычно используется для перемещения клеточных органелл. Стимулируя полимеризацию актина у одного из полюсов своих клеток, эти бактерии формируют своего рода актиновый хвост, который проталкивает их вперёд .
У некоторых бактерий имеются химические системы, испускающие свет. Способность к биолюминесценции часто имеется у бактерий, живущих в симбиозе с глубоководными рыбами , и свет, производимый бактериями, привлекает рыб друг к другу или более крупных животных к рыбам .
Бактерии часто формируют многоклеточные скопления, известные как биоплёнки, обмениваясь разнообразными химическими сигналами, за счёт которых их движение становится координированным . Формирование многоклеточных скоплений даёт бактериям ряд преимуществ: в них наблюдаются разделение труда между клетками и появление различных функциональных типов клеток, питательные вещества усваиваются более эффективно, обеспечивается более надёжная защита от естественных врагов. Например, бактерии в составе биоплёнок в 500 раз более устойчивы к антибиотикам, чем одиночные планктонные клетки того же вида .
Координированное поведение клеток одного и того же вида бактерий часто осуществляется за счёт особых химических веществ. На основе локальной концентрации этих веществ бактерия определяет плотность клеток-сородичей вокруг себя (чувство кворума). За счёт чувства кворума бактерии могут координировать экспрессию генов и начинают выделять и улавливать или феромоны , концентрация которых повышается по мере роста популяции .

Бактерий можно классифицировать на основе строения клетки, метаболизма, а также различий в химическом составе клеток (наличия или отсутствия некоторых жирных кислот , пигментов , антигенов, хинонов ) . В то время как перечисленные характеристики подходят для выделения штаммов, непонятно, можно ли их использовать для разделения видов бактерий. Дело в том, что у большинства бактерий нет отличительных структур, а из-за широко распространённого горизонтального переноса генов родственные виды могут сильно отличаться по морфологии и метаболизму . В связи с этим в настоящее время современная классификация базируется на молекулярной филогенетике . К числу её методов относят определение GC-состава генома, гибридизация геномов, а также секвенирование генов, которые не подверглись интенсивному горизонтальному переносу, такие как гены рРНК . Релевантная классификация бактерий публикуется «Международным журналом систематической бактериологии» ( англ. International Journal of Systematic Bacteriology ) и руководством по систематической бактериологии Берджи ( англ. Bergey's Manual of Systematic Bacteriology ). ( англ. International Committee on Systematics of Prokaryotes ) регулирует международные правила именования таксонов бактерий и определение их рангов согласно правилам ( англ. International Code of Nomenclature of Prokaryotes ) .
Термин «бактерии» традиционно применяли по отношению к микроскопическим одноклеточным прокариотам. Однако данные молекулярной филогенетики свидетельствуют о том, что в действительности прокариоты подразделяются на два независимых домена, которые первоначально получили названия эубактерии ( лат. Eubacteria ) и архебактерии ( лат. Archaebacteria ), но в настоящее время называются бактерии и археи . Эти два домена, наряду с доменом эукариоты, составляют основу трёхдоменной системы , которая является наиболее популярной системой классификации живых организмов . Археи и эукариоты состоят в более близком родстве, чем каждый из этих доменов к бактериям. Впрочем, высказывается мнение, что археи и эукариоты произошли от грамположительных бактерий . Поскольку количество отсеквенированных последовательностей бактериальных геномов очень быстро растёт, классификация бактерий постоянно меняется .
В медицине идентификация бактерий имеет огромное значение, поскольку от неё зависит схема лечения. По этой причине ещё до эры молекулярной биологии учёные активно разрабатывали методы, позволяющие быстро идентифицировать патогенные бактерии . В 1884 году Ганс Кристиан Грам предложил метод дифференциального окрашивания бактерий на основе строения их клеточной стенки . При окрашивании по Граму грамположительные бактерии с толстым слоем пептидогликана имеют фиолетовый цвет, а грамотрицательные бактерии с тонким слоем пептидогликана окрашены в розовый. Комбинируя окрашивание по Граму и морфотипы , выделяют четыре основные группы бактерий: грамположительные кокки, грамположительные бациллы, грамотрицательные кокки, грамотрицательные бациллы. Однако для идентификации некоторых бактерий больше подходят другие методы окрашивания. Например, микобактерии и бактерии рода Nocardia не после окрашивания по Цилю — Нильсену . Некоторых бактерий можно идентифицировать по их росту на специфических средах и при помощи других методов, например, серологии .
Методы разработаны так, чтобы способствовать росту определённых бактерий, но подавлять рост других бактерий из того же образца. Часто эти методы разрабатываются специально для определённых образцов, откуда берутся микробы. Например, для идентификации возбудителя пневмонии для дальнейшего культивирования берут образец мокроты , для идентификации возбудителя диареи для выращивания на селективной среде берут образец стула , причём во всех случаях рост непатогенных бактерий будет подавляться. Образцы, которые в норме стерильны (например, кровь , моча , спинномозговая жидкость ), культивируются в условиях, подходящих для роста любых микроорганизмов . После изоляции патогенного микроорганизма можно изучать его морфологию, особенности роста (например, аэробный или анаэробный рост), характер , а также окрашивать его разными методами.
Как и для классификации бактерий, молекулярные методы всё чаще применяют и для их идентификации. Диагностика, использующая такие молекулярные методы, как полимеразная цепная реакция (ПЦР), набирает всё большую популярность благодаря своей скорости и специфичности . С помощью этих методов можно обнаруживать и идентифицировать бактерии, которые, хотя и сохраняют метаболическую активность, не делятся и поэтому не могут быть выращены в культуре . Однако даже с помощью молекулярных методов точно определить или хотя бы примерно оценить число существующих видов бактерий невозможно. По состоянию на 2018, год описано несколько тысяч видов бактерий, но лишь около 250 из них являются патогенами человека . Общее число видов бактерий, по разным оценкам, составляет от 10 7 до 10 9 , но даже эти оценки могут быть на порядки меньше настоящего количества видов .
Однозначная и точная концепция вида бактерий так и не сформулирована. Это связано с невероятным разнообразием бактерий, широким распространением горизонтального переноса генов , невозможностью культивирования большинства бактерий и рядом других причин. Введение ПЦР и методов секвенирования в микробиологию позволило выделять виды бактерий на основании степени их сходства с геномами уже известных бактерий, однако и этот подход зачастую оказывается неэффективен из-за огромного разнообразия бактерий .
Помимо видов, при классификации бактерий иногда используют другие категории. К названию не до конца подтверждённых, а только предполагаемых видов добавляют слово Candidatus . Многие виды подразделяются на так называемые штаммы — морфологические или генетические варианты (подтипы) бактерий в пределах одного вида. Однако ряд специалистов считает категорию «штамм» искусственной .

Несмотря на видимую простоту, бактерии могут вступать в сложные взаимоотношения с другими организмами. Такие симбиотические отношения можно подразделить на паразитизм , мутуализм и комменсализм , а также хищничество. Из-за небольших размеров бактерии-комменсалы распространены повсеместно и обитают на всевозможных поверхностях, в том числе на растениях и животных. Рост бактерий на теле человека ускоряется от тепла и пота , и их большие популяции придают .
Некоторые бактерии убивают и поглощают другие микроорганизмы. К числу таких хищных бактерий относится Myxococcus xanthus , формирующая скопления, которые убивают и переваривают любую попавшую на них бактерию . Хищная бактерия прикрепляется к своей добыче, после чего постепенно переваривает её и всасывает высвобождающиеся питательные вещества . Daptobacter проникает внутрь других бактериальных клеток и размножается в их цитозоле . Вероятно, хищные бактерии произошли от сапрофагов , питающихся мёртвыми микроорганизмами, после того как приобрели приспособления для ловли и убийства других микробов .
Некоторые виды бактерий образуют скопления, которые необходимы для их выживания. Одна из таких мутуалистических ассоциаций, известная как межвидовая передача водорода, формируется между кластерами анаэробных бактерий, которые поглощают органические кислоты , такие как масляная и пропионовая кислоты , и выделяют водород, и метаногенными археями, которые используют водород. Бактерии из этой ассоциации не могут поглощать органические кислоты сами по себе, так как в ходе этой реакции образуется водород, накапливающийся вокруг. Только благодаря метаногенным археям концентрация водорода поддерживается достаточно низкой, чтобы позволить бактериям расти .
Многие бактерии являются симбионтами людей и других организмов. У человека от бактерий полностью свободны только кровь и лимфа . Например, более тысячи видов бактерий, входящих в состав нормальной кишечной микрофлоры человека, участвуют в работе иммунитета, синтезируют витамины (например, фолиевую кислоту , витамин K и биотин ), превращают сахара в молочную кислоту , а также сбраживают сложные неперевариваемые углеводы . Кроме того, кишечная микрофлора подавляет размножение патогенных организмов за счёт конкурентного исключения . Полезные микроорганизмы кишечной микрофлоры часто продают в виде пробиотических пищевых добавок .
Бактерии вступают в сложные мутуалистические отношения с самыми разными животными. Например, в губок обитает множество бактерий, причём все исследованные к настоящему времени виды губок имеют симбиотические ассоциации с одним или более видами бактериальных симбионтов . Многие моллюски имеют особые светящиеся органы, которые светятся благодаря обитающим в них бактериям. Бактерии получают надёжную защиту и благоприятные условия для питания, а моллюскам свечение помогает в привлечении полового партнёра . Асцидии вступают в симбиотические отношения с цианобактериями рода , который фиксирует CO 2 , а животное обеспечивает ему защищённое местообитание .
У жвачных животных в сложно устроенном желудочно-кишечном тракте обитает множество микроорганизмов, благодаря которым животные могут питаться почти что безбелковой пищей. Разрушать целлюлозу способны лишь некоторые бактерии, в результате деятельности которых образуются органические кислоты ( муравьиная , уксусная , пропионовая , масляная ), которые и усваиваются животными. Выделяющиеся углекислый газ и водород обитающие тут же метаногены превращают в метан . В одной из секций сложного желудка жвачных, рубце , обитают не только бактерии, разрушающие целлюлозу, но также бактерии, расщепляющие крахмал , пектин , полисахариды и пептиды , сбраживающие разнообразные сахара , спирты , аминокислоты и жирные кислоты . Целлюлозоразрушающие бактерии также населяют заднюю кишку термитов , образуя ацетат , который и усваивается насекомым .
В почве бактерии, входящие в состав ризосферы , осуществляют фиксацию азота, превращая его в различные азотсодержащие соединения . Они являются единственной усваиваемой формой азота для многих растений, которые сами не могут фиксировать азот. Множество бактерий обнаруживается на поверхности и внутри семян .

Бактерии, паразитирующие на других организмах, называют патогенами. Патогенные бактерии являются причиной множества человеческих смертей и вызывают такие инфекции, как столбняк , брюшной тиф , дифтерия , сифилис , холера , пищевые отравления , проказа и туберкулёз . Патоген, вызывающий заболевание, может быть описан много лет спустя после описания самой болезни, как, например, произошло с Helicobacter pylori и язвенной болезнью желудка . Бактерии ответственны за многие болезни культурных растений ( бактериозы ), в числе которых пятнистость листьев , ожог плодовых культур и увядание . Бактериальную основу имеют такие заболевания домашнего скота , как паратуберкулёз , мастит , сальмонеллёз и сибирская язва .
Каждый патоген характеризуется особыми взаимодействиями с организмом хозяина. Некоторые возбудители, такие как виды родов Staphylococcus и Streptococcus , вызывают кожные инфекции, пневмонию, менингит и даже сепсис , системный воспалительный ответ, переходящий в шок , массивную вазодилатацию сосудов и заканчивающийся смертью . При этом те же самые микроорганизмы входят в состав нормальной микрофлоры человека и зачастую обитают на коже и внутри полости носа , не вызывая никакого заболевания. Другие бактерии всегда вызывают болезнь, например, риккетсии , которые являются облигатными внутриклеточными паразитами и могут размножаться только внутри клеток организма-хозяина. Один вид риккетсий вызывает сыпной тиф , другой является возбудителем пятнистой лихорадки Скалистых гор . Другой род облигатных внутриклеточных паразитов, Chlamydia , включает возбудителей пневмонии, инфекций мочевыводящих путей и коронарной недостаточности . Некоторые бактерии, такие как Pseudomonas aeruginosa , и Mycobacterium avium , являются оппортунистическими патогенами и вызывают заболевания преимущественно у людей, страдающих от иммунодефицита или муковисцидоза .
Бактериальные инфекции можно лечить антибиотиками, в числе которых выделяют бактерицидные препараты, которые убивают бактерии, и , только подавляющие их рост. Существует несколько классов антибиотиков, которые действуют на процессы, которые есть у бактерии-патогена, но нет у организма-хозяина. Так, антибиотики хлорамфеникол и пуромицин подавляют работу бактериальной рибосомы, но не действуют на эукариотические рибосомы . Антибиотики используются не только в медицине, но и в животноводстве для стимуляции роста животных, что стало причиной повсеместного распространения устойчивости к антибиотикам в популяциях бактерий .
Некоторые бактерии, например, молочнокислые бактерии ( Lactobacillus и ), наряду с дрожжами и плесневыми грибками в течение нескольких тысяч лет использовались людьми для приготовления продуктов брожения, в числе которых сыры, квашеная капуста , соевый соус , уксус , вино и йогурт .
Способность бактерий разлагать разнообразные органические соединения находит применение в переработке отходов и биоремедиации . Бактерии, способные разрушать углеводороды нефти , часто используются для устранения разливов нефти . После выброса нефти из танкера «Эксон Валдез» в 1989 году в проливе Принца Вильгельма на некоторых из близлежащих пляжей были разложены удобрения, чтобы способствовать росту бактерий, разрушающих углеводороды нефти. Эта мера оказалась эффективной для тех пляжей, на которых слой нефти был не слишком толстым. Бактерии также используют для биоремедиации промышленных . В химической промышленности бактерии играют наиболее важную роль в производстве чистых энантиомеров химических соединений, которые используются в фармацевтике или агрохимии .
Бактерии можно использовать вместо пестицидов для биологической защиты растений . Наиболее часто для этих целей используется грамположительная почвенная бактерия Bacillus thuringiensis . Подвиды этой бактерии входят в состав инсектицидов , эффективных против чешуекрылых насекомых и известных под торговыми названиями Dipel и Thuricide . Благодаря узкой специфичности такие пестициды считаются экологичными и безопасными для человека , диких животных , опылителей и других .
Благодаря способности к быстрому росту и лёгкости манипуляций бактерии стали настоящими «рабочими лошадками» молекулярной биологии, генетики и биохимии . Внося мутации в геномы бактерий и изучая получившиеся фенотипы , учёные могут определять функции генов, ферментов и метаболических путей у бактерий и далее экстраполировать полученные данные на более сложные организмы . Для хорошо изученных бактерий, таких как E. coli , даже получены математические модели их метаболизма . Благодаря знаниям о метаболизме и генетике бактерий они могут использоваться в биотехнологии и биоинженерии для получения терапевтически важных белков, таких как инсулин , факторы роста и антитела .

Первым бактерии увидел голландский микроскопист Антони ван Левенгук в 1676 году с помощью микроскопа с одной линзой собственного изобретения . Он описал свои наблюдения в серии писем Лондонскому королевскому обществу . Размеры бактерий были почти на пределе разрешающей способности простых линз Левенгука. В течение почти сотни лет после открытия Левенгука бактерий никто не видел . Левенгук также описал протистов , которых назвал , и интерес к его открытиям появился в свете развития клеточной теории .
Термин «бактерии» предложил Христиан Готфрид Эренберг в 1828 году . Описанный им род на самом деле оказался сборной группой неспорообразующих палочковидных бактерий в противовес спорообразующим палочкам рода Bacillus , который Эренберг описал в 1835 году .
В 1859 году Луи Пастер показал, что в основе брожения лежит рост микроорганизмов, причём рост бактерий не был результатом их спонтанного самозарождения . Наряду со своим современником Робертом Кохом , Пастер стал первым приверженцем микробной теории болезней .
Роберт Кох , пионер медицинской микробиологии, работал с возбудителями холеры, сибирской язвы и туберкулёза. Коху удалось показать микробную основу туберкулёза, за что в 1905 году ему была присуждена Нобелевская премия по физиологии и медицине . Так называемые постулаты Коха , которые он ввёл как критерии для установления связи между микробом и болезнью, используются и по сей день .
Основателем бактериологии считается Фердинанд Кон , занимавшийся изучением бактерий с 1870 года. Он стал автором классификации бактерий на основе их морфологии .
Хотя в XIX веке стало известно, что многие болезни вызываются бактериями, эффективных антибактериальных препаратов всё ещё не существовало . В 1910 году Пауль Эрлих получил первый антибиотик, превратив красители для специфического окрашивания возбудителя сифилиса Treponema pallidum в соединения, которые селективно убивали бактерию . В 1908 году он был удостоен Нобелевской премии за свои работы в области иммунологии , однако он также стал пионером использования красителей для идентификации бактерий .
Положение бактерий в системе живого мира неоднократно менялось. С самого момента открытия их отнесли к растениям как класс Schizomycetes , а ныне выделяемые в отдельный тип цианобактерий (тогда известные как синезелёные водоросли) получили название Schizophyceae . Вместе с Schizomycetes они были выделены в отдел растений под названием Schizophyta . В 1866 году Эрнст Геккель выделил бактерий в тип дробянки ( Monera ), который рассматривал в составе царства протисты . Термин «прокариоты» для обозначения организмов, не имеющих клеточного ядра, был предложен Эдуардом Шаттоном в 1925 году наряду с термином «эукариоты». Шаттон также поместил цианобактерий в группу прокариоты из группы настоящих водорослей , которые являются эукариотами . В 1938 году Херберт Коупленд выделил дробянок из царства протистов, которое переименовал в . В 1969 году Роберт Уиттекер предложил деление живого мира на пять царств: дробянки, протисты, растения, грибы и животные . Колоссальный шаг вперёд в понимании биологии бактерий был совершён в 1977 году, когда Карл Вёзе разделил прокариот (которых по-прежнему нередко называли дробянками) на бактерий и архей . Филогенетическая таксономия , построенная на основе последовательностей гена 16S рРНК , легла в основу предложенной им трёхдоменной системы живого мира , которая закрепилась в 1990 году .
| Патогенные бактерии | |
|---|---|
|
Микрофлора
человека |
|
|
Субстратная
специфичность |
|
| Дыхание | |
|
Генетика и
размножение |
|
| Покоящиеся формы | |
| См. также | |
|
|
Некоторые
внешние ссылки
в этой статье
ведут на сайты, занесённые в
спам-лист
|