Гены старения
- 1 year ago
- 0
- 0

Гомеозисные гены (или гомеотические гены ) — гены , определяющие процессы роста и дифференцировки в организме. Гомеозисные гены кодируют транскрипционные факторы , контролирующие программы формирования органов и тканей .
Мутации в гомеозисных генах могут вызвать превращение одной части тела в другую. Гомеозисными мутантами называются такие организмы, у которых на месте органа развивается орган другого типа. Например, у дрозофилы при мутации antennapedia формируется конечность на месте антенны .
Гомеозисные гены контролируют работу других генов и определяют превращение внешне неразличимых участков зародыша или определённого органа (ткани, участка тела). В частности, гомеозисные гены контролируют появление различий сегментов многоклеточных животных в раннем эмбриональном развитии. У насекомых гомеозисные гены играют ключевую роль в определении особенностей строения эмбриональных сегментов и структур на них (ноги, антенны, крылья, глаза).
Гомеозисные гены животных относятся к семейству * . Однако не все гены этого семейства являются гомеозисными. Так, у дрозофилы к Hox-генам комплекса Antennapedia относятся гены zerknullt, zerknullt2, bicoid и fushi tarazu, которые не являются гомеозисными.
Есть ещё семейство генов . В геноме человека есть шесть генов ParaHox (GSX1, GSX2, , , , ), из которых три гена (GSX1, PDX1 (=IPF1), CDX2) образуют кластер на хромосоме 13 .
У растений также известны процессы, которые контролируются гомеозисными генами: филлотаксис, развитие цветков и соцветий.
Гомеозисные гены содержат гомеобокс — последовательность из 180 пар нуклеотидов ДНК , образующую в кодируемом белке гомеодомен .
Гомео домен впервые был обнаружен в составе генов, контролирующих развитие, и, в частности, в составе гомео зисных генов, у дрозофилы. Однако многие гены, содержащие гомеобокс, не являются гомеозисными. Таким образом, гомеобокс — это особая последовательность нуклеотидов, в то время как гомеозисность — это потенциальная возможность образования гомеозисной мутации.
Последовательность нуклеотидов в гомеобоксе высоко консервативна. Функциональная равнозначность гомеозисных белков может быть доказана тем фактом, что развитие мухи с соответствующими гомеозисными генами курицы протекает нормально. Несмотря на то, что общий предок курицы и мухи существовал около 670 миллионов лет назад, гомеозисные гены куриц сходны с аналогичными генами мух до такой степени, что могут заменить друг друга.
Из-за вырожденности генетического кода последовательность остатков аминокислот в составе белков более консервативна, чем последовательность нуклеотидов в ДНК, так как одну аминокислоту могут кодировать разные кодоны . Единственная мутация в ДНК гомеозисных генов может привести к поразительным изменениям организма (см. ).
Белковые продукты гомеозисных генов принадлежат к особому классу белков — транскрипционным факторам , которые связываются с ДНК и регулируют транскрипцию генов . Последовательность состоит из 60 аминокислотных остатков. У дрозофилы белковый продукт гомеозисного гена Antennapedia активирует гены, которые определяют структуру второго грудного сегмента, содержащего ноги и крылья, и репрессирует гены, вовлеченные в формирование глаз и антенн. Гены, которые регулируются белками, содержащими гомеобокс, называют реализаторными генами, и они являются белковыми продуктами генов полярности сегментов, которые кодируют ткане- и органо-специфичные белки.
Последовательность ДНК, с которой связывается гомеодомен, содержит последовательность нуклеотидов
TAAT
на
5'
-конце, причем
T
наиболее важна для связывания.
Данная последовательность нуклеотидов консервативна практически во всех сайтах связывания
гомеодомена
. Так как многие белки, содержащие гомеодомен, имеют одинаковые сайты узнавания, пары оснований, следующие за этой инициаторной последовательностью, используются для того, чтобы различать эти белки. Например, последовательность нуклеотидов после
TAAT
распознается девятой аминокислотой белка, содержащего гомеодомен. Белок, кодируемый геном с материнским эффектом,
Bicoid
, содержит остаток
лизина
в данном положении, который служит для узнавания и связывания
гуанина
. В белке
Antennapedia
в данном положении находится
глутамин
, который распознает
аденин
и связывается с ним. Если остаток лизина в белке Bicoid заменить на глутамин, измененный белок будет распознавать Antennapedia-специфичные энхансерные сайты.
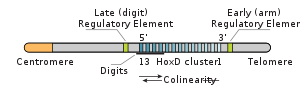
Hox-гены располагаются на одной или нескольких (до четырёх) хромосомах, обычно тесными группами (кластерами), внутри которых сохраняется более или менее строгий порядок: «головные» гены впереди, «хвостовые» — сзади. У более примитивных представителей многоклеточных, таких как гребневики (Ctenophora) и кишечнополостные (Cnidaria), этих эмбриональных регуляторных генов только четыре, у млекопитающих их уже 48.
Семейство Hox-генов подразделяется на 14 классов. Считается, что эти 14 классов возникали путём дупликации одного или немногих исходных генов, реплики затем мутировали и обретали новые функции. У примитивных кишечнополостных и гребневиков имеется всего 4 класса Hox-генов, у предполагаемого общего предка двустороннесимметричных животных их должно было быть по крайней мере 8, у млекопитающих присутствуют все 14 классов. Принцип работы этих генов одинаков. Их продукты являются транскрипционными факторами, функция которых состоит во «включении» или «выключении» других генов. В результате работы Hox-факторов запускается каскад реакций, приводящий к появлению в клетке нужных белков.
За последнее десятилетие расшифрованы ДНК -последовательности Hox-генов у многих групп животных: аннелид, плоских червей, иглокожих, нематод, членистоногих, оболочников, ланцетников, не говоря уже о млекопитающих.
Гомеозисные гены регулируют работу реализаторных генов, и, в свою очередь, регулируются генами и , которые находятся под контролем белков-морфогенов ряда генов с материнским эффектом . В результате этого образуется каскад транскрипционных факторов : гены материнского эффекта включают гены gap и pair-rule; гены gap и pair-rule включают гомеозисные гены; наконец, гомеозисные гены включают реализаторные гены, которые приводят к сегментации и дифференцировке зародыша.
Такая регуляция осуществляется градиентами концентрации белков-морфогенов. Высокая концентрация одного из материнских белков и низкая — других включает определенный набор генов gap и pair-rule. У мух вторая полоска экспрессии гена Even-skipped эмбриона активируется материнскими белками Bicoid и Hunchback и репрессируется белками gap Giant и .
Молекулы микроРНК в hox-кластерах сильнее ингибируют передние гомеозисные гены, вероятно, для более точной регуляции их экспрессии.
В кластерах гомеозисных генов широко распространены некодирующие РНК (ncRNA). Один из генов некодирующих РНК у человека, HOTAIR, снижает уровень транскрипции гомеозисных генов (транскрибируется с кластера HOXC и ингибирует поздние гены HOXD), связываясь с белками группы Polycomb (PRC2).
Структура хроматина необходима для транскрипции , но также требуется выпетливание хромосомных территорий, на которых располагается кластер. Количественная ПЦР показала некоторые закономерности коллинеарности: система находится в равновесии и общее количество транскриптов зависит от количества генов, представленных в линейной последовательности.
Ошибки экспрессии гомеозисных генов приводят к крупным изменениям в морфологии индивида. Гомеозисные мутации впервые были описаны в 1894 году Уильямом Бэтсоном , который описал появление тычинок на месте лепестков.
В конце 1940-х годов на модельном объекте Drosophila melanogaster Эдвард Льюис изучал гомеозисные мутации, которые приводили к формированию причудливых органов. Мутации в генах, участвующих в развитии конечности, могут приводить к уродствам или даже к смерти. Например, мутации в гене Antennapedia приводят к образованию конечностей на голове мухи на месте антенн.
Другим известным примером у дрозофилы является мутация в гомеозисном гене , который определяет развитие третьего грудного сегмента. Обычно на этом сегменте есть пара ног и пара жужжалец (редуцированных крыльев). У мутантных особей, которые не имеют функционального белка Ultrabithorax, на третьем сегменте образуются такие же структуры, как на втором грудном сегменте, который несёт пару конечностей и пару полностью развитых крыльев. Такие мутанты иногда встречаются в диких популяциях дрозофил, и изучение таких мутантов привело к открытию гомеозисных генов животных.

Гомеозисные гены в хромосомах у многих животных располагаются очень близко друг к другу, образуя при этом кластеры. При этом у дрозофилы наблюдается коллинеарность — последовательность расположения генов на хромосоме соответствует последовательности их экспрессии вдоль передне-задней оси тела.
В различных таксонах гомеозисным генам были даны разные названия, что привело к путанице в номенклатуре. В случае некоторых первичноротых ( Ecdysozoa — членистоногих, нематод) гомеозисные гены составляют два кластера Antennapedia и , которые вместе называют HOM-C (Гомеозисный комплекс, Homeotic Complex). В случае вторичноротых (иглокожих, хордовых) гомеозисные гены называют Hox -генами и выделяют четыре кластера: Hoxa, Hoxb, Hoxc и Hoxd. У первичноротых гомезисные гены также часто называют Hox-генами, хотя это и не вполне верно.
У Ecdysozoa существует около десяти гомеозисных генов. Позвоночные имеют четыре набора паралогов этих десяти генов Hoxa, Hoxb, Hoxc, и Hoxd. Эти кластеры паралогов образовались в результате двух дупликаций геномов позвоночных.
Обе дупликации произошли после того, как от общего ствола с позвоночными отделились предки ланцетников и оболочников, и до того, как разделились эволюционные линии млекопитающих и хрящевых рыб. Вероятнее всего, первая дупликация произошла незадолго до разделения линий бесчелюстных и челюстноротых, а второе — вскоре после этого (разделение этих линий, вероятно, произошло около 530 млн лет назад).
Хотя гомеозисные гены позвоночных являются копиями генов Ecdysozoa , эти копии не идентичны. В результате накопления мутаций в течение долгого времени белки выполняют различные функции. У разных групп позвоночных некоторые гены утрачены или дуплицированы.
Hoxa и Hoxd определяют развитие конечности. Экспрессия Hox в конечности имеет две стадии — на первой развивается собственно конечность, на более поздней работают Hoxd 8 — 13 и образуются пальцы, при этом задействована отдельная регуляторная область на 5’-конце гена Hoxd 13, который не встречается у Teleostei .
На важное значение мутаций в гомеозисных генах для развития теории наследственности впервые указал автор этого термина Уильям Бэтсон в 1894 году. В 1920-е годы изучением гомеозисных мутаций (в том числе на дрозофиле ) занималась ученица С. С. Четверикова . Балкашина описала мутацию aristopedia у дрозофилы и установила параллелизм явлений гомеозиса при регенерации и при мутировании гомеозисных генов, а также картировала три известных в то время гомеозисных гена дрозофилы.
Эдвард Льюис в 1948 году начал систематическое изучение гомеозисных генов, управляющих развитием имагинальных дисков личинки в органы имаго . Льюис обнаружил коллинеарность в пространстве между порядком расположения генов комплекса в хромосоме и порядком расположения имагинальных дисков (сегментов), за развитие которых они отвечают, вдоль передне-задней оси тела.
Кристиана Нюсляйн-Фольхард и Эрик Вишаус классифицировали 15 генов, определяющих строение тела и образование сегментов у Drosophila melanogaster . В 1995 году исследователи получили Нобелевскую премию по медицине.
Испанские ученые в январе 2013 года провели эксперимент по внедрению в генотип рыбы данио-рерио гена hoxd13, отвечающего за развитие конечностей для передвижения по суше, заимствованного у мышей. Подобный ген есть и у самих рыб, но не проявляет достаточной активности для развития лап. В результате эксперимента рыбы вместо плавников получили зачатки конечностей, которые могли бы обеспечить передвижение по земле.
Экспрессия генов, регулирующих развитие растений, находится под контролем внутренних и внешних факторов. К внутренним факторам, влияющим на их активность, относят гормоны , сахарозу и некоторые минеральные элементы, к внешним — температуру и свет. В регуляции процессов дифференцировки и развития важная роль принадлежит генам, которые содержат промоторы , чувствительные и специфичные к фитогормонам и к таким факторам внешней среды, как свет и температура. В составе промоторов очень многих генов, активность которых регулируется фитогормонами, выявлены транскрипционные элементы, определяющие гормональную специфичность ростовых реакций растений.
В настоящее время идентифицированы ключевые гены, которые контролируют эмбриогенез , старение и фотоморфогенез, регулируют функционирование апикальных, латеральных и флоральных меристем , отвечают за формирование корня, листьев и сосудов. Наиболее хорошо изучена экспрессия генов, регулирующих развитие цветков. На основе имеющейся в настоящее время генетической информации, математического аппарата и компьютерных программ стало возможным построение так называемых генетических регуляторных сетей, которые позволяют оценить весь спектр взаимодействий между различными регуляторными генами в процессе дифференцировки клеток и формирования органов растения. Отдельные элементы этих сетей способны на разных этапах развития контролировать несколько процессов. Поэтому мутации, затрагивающие разные участки одного регуляторного гена, могут отличаться своим фенотипическим проявлением.
У высших растений наиболее хорошо изучено функционирование двух типов генов-регуляторов развития: гомеобокссодержащих и генов с MADS-боксом .
Гены, содержащие гомеобокс , определяются по наличию характерной последовательности ДНК из приблизительно 180 пар нуклеотидов (гомеобокса), кодирующей гомеодомен — консервативный участок ряда транскрипционных факторов. Эта нуклеотидная последовательность типична для генов каскадного типа регуляции развития.
Первым клонированным геном растений, кодирующим гомеодоменсодержащий белок, был KNOTTED1 (KN1) кукурузы. Мутация knotted 1 приводит к тому, что ген KN1 начинает экспрессироваться в несоответствующее время и не в том месте. У мутантов kn1 вокруг уже дифференцированных клеток листа появляются группы клеток, которые ещё продолжают делиться. Группы делящихся клеток, расположенные вдоль сосудистых элементов по всей листовой пластинке, образуют так называемые узлы (knots). Позднее было обнаружено целое семейство генов, подобных KN1, названное KNOX (KNOTTED1-like HOMEOBOX). Сверхэкспрессия генов семейства KNOX также искажает развитие листа.
Среди KNOX-генов растений наиболее детально исследована большая группа, участвующая в регуляции деятельности апикальной меристемы побегов и в развитии листьев: KN1 и RS1 у кукурузы, KNAT1, KNAT2 и STM у Arabidopsis thaliana , HvKNOX3 у ячменя и OSH1 у риса. Гены KN1, STM и их функциональные аналоги отвечают за поддержание деления клеток меристем, репрессируя их дальнейшую дифференцировку. Эти гены экспрессируются в апикальных меристемах побегов, а также во флоральных меристемах.
Термин «MADS-бокс» образован начальными буквами четырёх генов: MCM1 дрожжей, AG арабидопсиса, DEF львиного зева и SRF млекопитающих. К генам, содержащим MADS-бокс, относятся, в частности, AG ( AGAMOUS ), DEF (DEFICIENCE), AP1 (APETALA1) и AP3 (APETALA3), TFL1 (TERMINAL FLOWER), PI (PISTILLATA). Гены этого типа регулируют флоригенез и определяют судьбу клеток в семяпочке; их экспрессия выявлена в зародыше, корнях и листьях. К MADS-бокс-генам относится большинство гомеозисных генов растений, в частности гены идентичности органов цветка. Предполагается, что возникновение новых органов в процессе прогрессивной эволюции растений, например семяпочек и семян, сопровождалось появлением новых подсемейств именно MADS-бокс-генов.
Непосредственный контроль за развитием органов и тканей растения осуществляется транскрипционными факторами (ТФ) — белками, которые после перемещения в ядро клетки регулируют транскрипцию, специфически взаимодействуя с ДНК либо с другими белками, которые могут образовывать комплекс белок-ДНК.
|
|
|
|---|