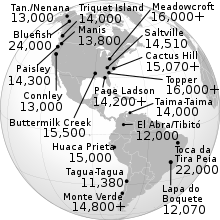
Древние контакты между Китаем и Средиземноморьем
- 1 year ago
- 0
- 0


Щелевы́е конта́кты ( англ. gap junctions ) — межклеточные контакты , обеспечивающие прямой перенос ионов и небольших молекул между соседними клетками . Щелевые контакты способны образовывать почти все клетки животных . Каналы щелевых контактов имеют цилиндрическую форму и состоят из двух половин — коннексонов , или полуканалов. Каждый коннексон состоит из шести белковых субъединиц — . Проницаемость щелевых контактов регулируется путём открытия и закрытия ворот канала (гейтинг). Гейтинг, в свою очередь, контролируется изменением клеточного pH , концентрацией ионов кальция или непосредственным фосфорилированием коннексинов. Помимо коннексинов, известны и другие семейства белков щелевых контактов, поэтому можно предположить, что щелевые контакты неоднократно возникали в ходе эволюции животных .
Щелевые контакты могут содержать от нескольких десятков до многих тысяч каналов, проходящих сквозь плазматические мембраны смежных клеток. Каждый канал и состоит из двух половин, которые известны как коннексоны, или полуканалы. Эти половины стыкуются в узкой щели шириной 2—3 нм , разделяющей соседние клетки. Каждый коннексон состоит из 6 белковых субъединиц — коннексинов. Коннексон имеет цилиндрическую форму и представляет собой гидрофильный канал длиной 17 нм и диаметром 7 нм в наиболее широкой части и 3 нм в наиболее узкой. Субъединица коннексина содержит 4 α-спирали , связанных внеклеточными петлями. По всей видимости, внеклеточные петли противоположных коннексинов связываются друг с другом посредством антипараллельных β-слоёв , образуя β-бочонок .
В геноме человека закодировано не менее 20 различных коннексинов, и во многих клетках экспрессируется более одного типа коннексинов. В связи с этим различают гомоолигомерные коннексоны, состоящие из одинаковых коннексинов, и гетероолигомерные коннексоны, которые содержат субъединицы разных типов. В одной бляшке щелевого контакта могут находиться коннексоны с разным составом коннексинов, причём в пределах бляшки коннексоны или располагаются гомогенно, или пространственно разобщены в соответствии с составом коннексинов. В коннексинах идентифицированы домены , необходимые для стыковки двух коннексонов, для узнавания других коннексинов, для образования олигомеров и сочетаемости коннексиновых субъединиц . В большинстве клеток коннексины быстро деградируют, и время их полужизни составляет около 15 часов .
Известны ещё два семейства белков щелевых контактов. обнаружены только у беспозвоночных животных , однако они не являются гомологами коннексинов. Тем не менее, они формируют межклеточные контакты, которые по строению и по функциям похожи на щелевые контакты позвоночных . Другое семейство представлено паннексинами , которые имеются как у позвоночных, так и у беспозвоночных. По структуре они отличаются и от коннексинов, и от иннексинов. Паннексины обнаруживаются почти исключительно в нейронах и, вероятно, играют важную роль в их функционировании и развитии даже у животных с примитивной нервной системой . В геноме человека и мыши к настоящему времени идентифицированы 3 гена паннексинов . У иглокожих и некоторых других групп животных есть щелевые контакты, но нет генов ни одного из вышеназванных семейств. Это означает, что существуют ещё не открытые семейства белков щелевых контактов . Таким образом, можно заключить, что щелевые контакты появлялись в ходе эволюции животных не менее двух раз, поэтому они представляют собой продукт конвергентной эволюции .

Сборка новых коннексонов из коннексинов происходит внутри секреторных везикул . Новые коннексоны добавляются на периферии бляшек щелевых контактов, а старые коннексоны удаляются из их центральной части .
Проницаемость щелевых контактов может регулироваться открытием и закрытием каналов (этот процесс известен как гейтинг, или воротный механизм). Гейтинг каналов находится под влиянием таких факторов, как клеточный pH, мембранный потенциал , концентрация ионов кальция и фосфорилирование коннексинов. Показано, что при увеличении концентрации ионов кальция от 10 -7 М до 10 -5 М проницаемость каналов уменьшается, и при концентрациях выше 10 -5 М каналы полностью закрываются. Вероятно, закрытие каналов щелевых контактов в ответ на возрастание концентрации ионов Ca 2+ служит механизмом защиты от апоптоза , так как при апоптозе внутриклеточная концентрация ионов кальция резко увеличивается, и, если щелевые контакты не будут закрыты, апоптоз произойдёт и в соседних клетках . На динамику щелевых контактов влияют некоторые соединения. Например, олеамид ( амид жирной кислоты , который образуется в головном мозге ) блокирует щелевые контакты и вызывает сон . Органические спирты ( и октанол ) и общие анестетики ( галотан ) также могут вызывать обратимое закрытие щелевых контактов, однако их действие не ограничивается щелевыми контактами. Коннексины могут фосфорилироваться несколькими протеинкиназами . Например, в течение нескольких секунд цАМФ активирует протеинкиназу A , которая фосфорилирует C-концевые участки коннексинов, снижая или увеличивая долю открытых каналов в зависимости от изоформы коннексина и типа клетки. В долговременной перспективе (несколько часов) цАМФ также способствует сборке щелевых контактов .
Щелевые контакты служат для перемещения ионов и малых молекул между соседними клетками. Через щелевой контакт могут проходить молекулы массой до 1,2 кДа , а молекулы с массой 2 кДа задерживаются. Клетки могут обмениваться такими молекулами, как сахара , нуклеотиды , вторичные посредники (цАМФ или цГМФ ), небольшие пептиды и РНК . Щелевые контакты особенно важны, когда большому количеству клеток необходимо выдать быстрый, хорошо скоординированный ответ. Так, щелевые контакты составляют основу очень быстрых электрических синапсов , которые можно найти, например, в нейронах головного мозга и в клетках миокарда ( кардиомиоцитах ) .
Щелевые контакты встречаются практически во всех тканях . Одним из исключений является поперечно-полосатая мускулатура , где клеткам не требуется электрическая связь, поскольку клетки слиты в симпласт (однако щелевые контакты встречаются в сосудах, питающих мышцы ). Также щелевые контакты не обнаруживаются у эритроцитов и зрелых сперматозоидов . Как правило, большая часть каналов в щелевых контактах закрыта: так, доля открытых каналов в кардиомиоцитов составляет около 0,2, а в нейронах — 0,01 .
Точечные мутации в генах, кодирующих коннексины, у человека приводят к очень специфическим дефектам, из чего можно заключить, что большая часть коннексинов экспрессируется лишь в нескольких тканях. Рецессивные мутации в гене являются наиболее частой причиной наследственной глухоты . Коннексин-26 участвует в транспорте ионов калия в клетках эпителия , поддерживающего чувствительные волосковые клетки в ухе . Люди с мутациями в гене, кодирующем , могут страдать от разрушения миелиновой оболочки аксонов ( X -связанный вариант болезни Шарко — Мари — Тута ). Возможно, стабильность миелина может зависеть от щелевых контактов между клетками разных слоёв миелиновой оболочки, и нарушения в функционировании контактов приводят к её разрушению .
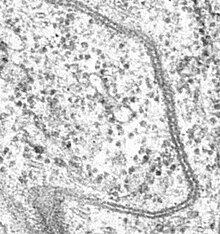
Первые свидетельства о существовании щелевых контактов появились в 1960-х годах, когда было показано, что электрический импульс передаётся между смежными клетками непосредственно, а не через разделяющую их жидкость. Кроме того, было продемонстрировано, что при введении в культуральную среду флуоресцентных молекул они проходят между соседними клетками гораздо быстрее, чем если бы они проходили насквозь через клеточные мембраны . На основании этих экспериментальных данных было высказано предположение, что клетки обмениваются ионами и малыми молекулами через каналы, которые пронизывают плазматическую мембрану и непосредственно соединяют соседние клетки. Позднее с помощью электронной микроскопии удалось получить изображения щелевых контактов, что окончательно подтвердило их существование .
|
|||||
| Запирающие контакты |
|
||||
| Контакты, формирующие каналы |
|
||||