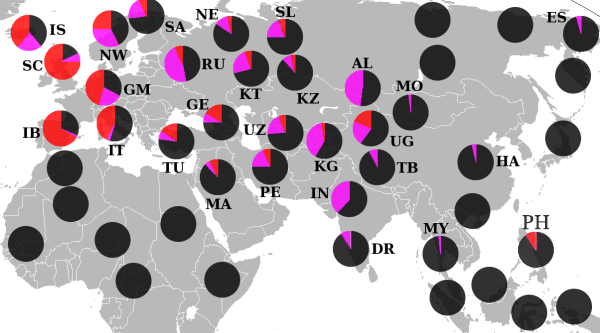
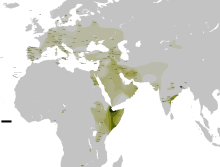
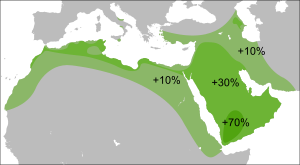
Эта гаплогруппа имеет две основных субгаплогруппы (субклада): С1-F3393/Z1426 (ранее CxC3, то есть старый C1, старый С2, старый С4, старый C5 и старый C6) и С2-M217.
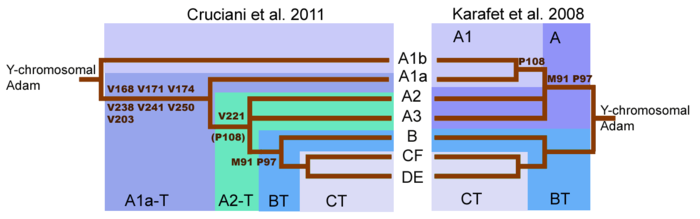
Это одна из очень древних гаплогрупп, распространившихся из вторичного Ближневосточного очага этногенеза. В период с 88 000 по 68 500 проживали потомки гаплогруппы СТ, в период 68 500 по 65 200 проживали потомки гаплогрупп DE CF, и только с этого времени начинается бурное ветвление С, F и D, E.
Определяется
полиморфизмами уникальных событий
M130/RPS4Y711, P184, P255, P260 — все они являются мутациями
SNP
. Является родственной
Y-хромосомной гаплогруппе F
, которая также является потомком более древней
Y-хромосомной гаплогруппы CF
. В отличие от других гаплогрупп, возникших в тот же период (включая потомков), все подклассы гаплогруппы CF являются неафриканскими, а гаплогруппа C интересна тем, что продвинулась на восток особенно далеко.
Содержание
Распространение
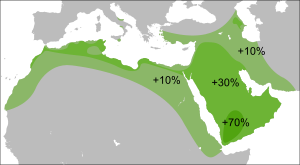
Распространение гаплогруппы C-M130 обычно ограничивается Сибирью, некоторыми частями Восточной Азии, Океании и Америки . Из-за огромного возраста она успела накопить многочисленные вторичные мутации, и было идентифицировано множество регионально важных подветвей этой гаплогруппы.
Субклад C1a2 доминировал в Западной и Центральной Европе до
неолита
, будучи основной гаплогруппой европейских кроманьонцев. В настоящее время доля C1a2 в Европе невелика, будучи вытесненная сперва неолитическими (G2, I1, I2) так и более поздними переселенцами — носителями R1a, R1b, N1c, E1b1b.
Теории миграции
Y-хромосомная гаплогруппа C-M130: происхождение, субклады, предполагаемые миграции
Маршрут миграции гаплогруппы C в Восточной и Южной Азии
Определяющая гаплогруппу
C
мутация
M130 произошла, скорее всего, в период 88 000-68 500 лет назад когда
ОНП
-мутации M168 из
Y-хромосомной гаплогруппы BT
выделилась
Y-хромосомная гаплогруппа 'CT
, а из последней — посредством мутации P143
гаплогруппа CF
. По данным компании YFull Y-хромосомная гаплогруппа C-M130 образовалась 65,9 тыс. л. н.
Последний общий предок
современных носителей гаплогруппы C-M130 жил 48,4 тыс. л. н.
образовалась 48 800 л. н. Последний общий предок современных носителей гаплогруппы C1-F3393 жил 47 200 л. н.
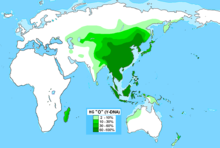
Распространение Y-хромосомной гаплогруппы C2-M217
C2-M217 образовалась 48 800 л. н. Последний общий предок современных носителей гаплогруппы
(ранее C3) жил 34 000 лет назад
.
Гаплогруппа С претерпела первоначальное быстрое расширение вскоре после 54 тыс л. н. и к 50 тыс. л. н. уже имела восемь субветвей. Линии
найдены сегодня от Европы до Восточной, Юго-Восточной, Южной Азии и в Океании, в то время как более распространённые линии
в настоящее время встречаются в Восточной и Южной Азии, а в Центральной и Западной Азии
появились относительно поздно в период экспансии монголов.
Хотя сегодня наибольшая концентрация гаплогруппы
C
наблюдается среди коренного населения
Монголии
,
Дальнего Востока России
,
Полинезии
,
аборигенов Австралии
. Гаплогруппа С доминирует у западных бурят, эвенков и юкагиров, достигая частот 61,3 %, 69,2 % и 80 % соответственно
.
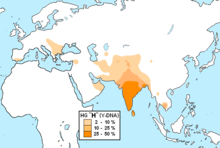
Наибольшее разнообразие данной гаплогруппы обнаружено среди населения Индии, из чего некоторые исследователи делают предположение, что она либо произошла, либо существовала наиболее длительное время в своей истории на территории побережья
Южной Азии
. Субклад C8-CTS5573 был обнаружен у японцев, отобранных в
Токио
(JPT). Японский субклад
вместе с европейской и непальской линией
образуют одну ветвь C1a-CTS11043 (C1'8-CTS824). Это поддерживает гипотезу о том, что общий предок европейской и непальской ветви C1a2-V20 жил в палеолите в
Восточной Азии
(по данным компании YFull субклад C1a2-V20 образовался 46,7 тыс. л. н., последний общий предок современных носителей субклада C1a2-V20 жил 42,5 тысячи лет назад
)
.
По мнению ряда исследователей, гаплогруппы
C
и
D
попали в Восточную Азию вместе, в рамках одной и той же популяции, которая впервые успешно колонизировала этот регион, однако в настоящее время распространение гаплогрупп
C
и
D
сильно различается. Различные субклады гаплогруппы
C
встречаются с высокой частотой среди
аборигенов Австралии
,
полинезийцев
,
микронезийцев
,
монголов
, западных
бурят
,
калмыков
,
казахов
и аборигенных народов Дальнего востока России, а также с умеренной частотой среди
корейцев
и
маньчжуров
. С частотой около 15 % субклад C2-M217 встречается у айнов
. С другой стороны,
гаплогруппа
D
встречается с высокой частотой только среди
тибетцев
,
японцев
(особенно
айнов
) и жителей
Андаманских островов
, однако не обнаружена ни в Индии, ни среди индейцев или аборигенов Океании.
По данным компании YFull гаплогруппа C1-F3393/Z1426 разделилась на ветви C1a-CTS11043 и C1b-F1370 47 300 лет назад
. Разделение гаплогрупп C1-F3393/Z1426 и С2-M217 на субклады происходило в Сибири до
Последнего ледникового максимума
.
По данным генетиков линии C1b-F1370 (ISOGG 2018) из Южной Азии, представленными субкладами C1b1a1-M356 из Индии и C1b1a2-AM00847/AMM008/B65 из Борнео (
Калимантан
), разошлись с линиями, представленными субкладами C1b2b-M347/P309 из Австралии и C1b2a-M38 из Новой Гвинеи, 54 тыс. л. н. (95%-й
доверительный интервал
: 47,8—61,4 тыс. л. н.). C1b2b аборигенов Австралии и C1b2a папуасов разошлись 50,1 тыс. л. н. (95%-й
доверительный интервал
: 44,3—56,9 тыс. л. н.)
. По данным компании YFull гаплогруппа C1b-F1370 разделилась на ветви C1b1-AM00694/K281 и C1b2-B477/Z31885 47 200 лет назад
.
В период от 15,3 до 14,3 тыс. л. н. от гаплогруппы C2-M217 образовалось 8 субветвей. В
Берингию
мигрировали 3 субклада, а Америки достиг один субклад C2b1a1a-P39 (ISOGG 2018) или C2a1a1a-P39 (ISOGG 2019)
. C2a1a1-F3918 включает в себя субклад C2a1a1a-P39, который с высокой частотой обнаруживался в выборках некоторых коренных североамериканских популяций, и субклад C2a1a1b-FGC28881, который в настоящее время встречается с различной (но в целом довольно низкой) частотой по всей
евразийской степи
от провинций Хэйлунцзян и
Цзянсу
на востоке до турецкой провинции
Гиресун
,
Подляского воеводства
и
Южно-Чешского края
на западе.
Предполагается, что в Америку гаплогруппа C2-M217 попала около 8—6 тысяч лет назад вместе с носителями языков
на-дене
и разошлась вдоль северо-западного побережья
Северной Америки
.
Структура
C* (M130/Page51/RPS4Y711, M216)
имеет два субклада:
C1
и
C2,
которые имеют свои нижестоящие субклады.
C1 (F3393)
C1a (CTS11043)
C1a1 (M8)
C1a1a (P121)
C1a1a1 (CTS9336)
C1a1a1a (CTS6678)
C1a1a1b (Z1356)
C1a1a2 (Z45460)
C1a2 (ранее C6) - (V20)
C1a2a (V182)
C1a2a1 (V222)
C1a2a2 (Z29329)
C1a2b (Z38888)
C1b (F1370)
C1b1 (K281)
C1b1a (B66/Z16458)
C1b1a1 (ранее C5) - (M356)
C1b1a2 (B65)
C1b2 (B477/Z31885)
C1b2a (ранее C2) - (M38)
C1b2a1 (M208)
C1b2a1a (P33)
C1b2a1b (P54)
C1b2b (ранее C4) - (M347)
C1b2b1 (M210)
C2 (ранее C3) - (M217)
C2a (M93)
C2b (L1373/F1396)
C2b1
C2b1a
C2b1a1
C2b1a1a (P39)
C2b1a2 (ранее C3c) - (M48)
C2c (C-F1067)
C2c1 (F2613/Z1338)
C2c1a (Z1300)
C2c1a1
C2c1a1a
C2c1a1a1 (M407)
Палеогенетика
C1 (C-F3393) определили у образцов ВВ7-240 и CC7-335 возрастом более 45 тыс. л. н. из пещеры
Бачо Киро
(
Болгария
)
C1a2 определили у
ZBC из местечка Pınarbaşı в турецкой провинции Конья, жившего 13 642 — 13 073 лет до нашей эры
.
C2a1a1-F3918>C-F3918*
(ISOGG 2019) определили у представителя
UKY001 (14 054—13 771 л. н.) из
Усть-Кяхты
в
Бурятии
C1a1a определили у представителя
азильской культуры
BAL003 (12 830—10 990 лет до настоящего времени) из Бальма де Гилания или Бальма де ла Гвинеу (Balma Guianya) в Пиренеях (испанская провинция
Льейда
)
Гаплогруппу C определили у анатолийского фермера ZMOJ из местечка Бонджуклу (Boncuklu), жившего 8300—7800 лет до нашей эры (
Анатолия
)
C2c-P53.1 определили у представителя мезолитической культуры
Лепенски-Вир
C2b (ISOGG 2018) или C2a>C-L1373 (ISOGG 2019) определили у образца CP19 (Lapa01 (Burial 1)) из
бразильского
(10 160—9600 лет до настоящего времени)
C1a2 определили образцов 2779 F.265 и 5357 F.576 (7035—6680 лет до н. э.) из
Чатал-Хююка
(Турция)
C1a2-V20 определили у неолитического образца из местонахождения
в Турции с датировкой 8500—8200 лет назад
C1a2b определена у образца LEPE48 (8012—7867 лет
до настоящего времени
) из
Лепенски-Вира
в
Сербии
и у представителя культуры линейно-ленточной керамики (LBK) Dil16 (7235—6998 лет до н. в.) из Dillingen-Steinheim в Германии
Гаплогруппу C определили у образца La368 культуры
Хоа-Бинь
(около 7888 лет назад) из Лаоса
У темнокожих и голубоглазых
мезолитических
братьев из пещеры
(
), найденных в провинции Леон (северо-запад Испании) определена Y-хромосомная гаплогруппа C1a2 (ранее обозначалась как C6)
Гаплогруппу C определили неолитического обитателя
Малык-Преславеца
(Болгария)
C1a2 определили у представителя культуры кардиальной керамики из хорватской пещеры Земуника (
Zemunica
) близ
Биско
, жившего около 7,5 тыс. лет назад
C1a2 определили у раннеолитического (~5460—5220 лет до н. э.) образца UZZ61 из
Сицилии
C1a2 (ранее обозначалась как C6) определили у представителя
культуры линейно-ленточной керамики
из венгерского местонахождения Apc-Berekalja I, жившего примерно 4950—5300 лет назад, и у представителя восточной культуры линейно-ленточной керамики (Alföld Linear Pottery) из венгерского местонахождения Kompolt-Kigyósér, жившего примерно 4990—5210 лет назад
C2b-F1396 определили у представителей
(~5000 лет до н. э.) в Приморском края
Гаплогруппу C определили у представителя культуры неолита/халколита
Хуншань
(
Китай
)
C2b1a1 (ISOGG 2018) определили у энеолитического образца NIZ001 (4445–4337 лет до н. э.) из Нижнетытескенской пещеры I в
Горном Алтае
C1a-V20>C-V86>C-V182>C-V3918>
C-Y173810>C-Y173810* определили у представителя трипольской культуры VERT103 из украинской пещеры
Вертеба
C1a2 определили у образца AID002 из позднего бронзового века Греции (
микенский
некрополь
Аидония
, мтДНК:
R0a1a
)
C2b1b/F845 и C2a1a1b1a/F3830 определили у 6 кочевников с Монгольского плато, живших в период с позднего неолита до династии Юань (~3500—700 л. н.)
C1b1a2b определили у образца AMA001 (2258 ± 30 л. н.) с острова
Моротай
(Aru Manara, провинция
Северное Малуку
, Индонезия)
C2a2-M217>MPB373/L1373>F1756 (ISOGG 2019) определили у образца bla001 (1344—1270 л. н., VII век) со стоянки Октябрьское (Oktyabr'skoe (burial 2)) в
Амурской области
C2a1a1b1b-Y10442/etc (xZ31688,Z31693) определили у раннего
аварского
военного лидера FGDper4 (Early Avar military leader, 620—660 гг.)
C2b1a3a1a-Y4580 (с отрицательной Y4541) определили у члена армии хана Джучи (образец DA28), похороненного в золотоордынском погребении XIII—XIV века в горах
Улытау
(
Казахстан
)
↑
Генофонд казахов
(неопр.)
. Web (22 января 2017). Дата обращения: 17 октября 2019.
14 января 2020 года.
Copyright 2015 by ISOGG.
(неопр.)
. www.isogg.org. Дата обращения: 30 сентября 2015.
15 августа 2021 года.
В. Н. Харьков, К. В. Хамина, О. Ф. Медведева, К. В. Симонова, Е. Р. Ерёмина, В. А. Степанов.
// ГЕНЕТИКА, 2014, том 50, № 2, стр. 203–213.
25 октября 2020 года.
Жабагин М. К.
Анализ связи полиморфизма Y-хромосомы и родоплеменной структуры в казахской популяции / О. П. Балановский. — Москва, 2017. — С. 54. — 148 с.
Жаксылык Сабитов, Батыр Даулет.
(рус.)
// The Russian Journal of Genetic Genealogy. — 2013. —
Т. 5
,
№ 1
. —
С. 48-51
. —
ISSN
.
14 июля 2020 года.
Wang, CC; Li, H.
Inferring human history in East Asia from Y chromosomes
(англ.)
// Investig Genet : journal. — 2013. —
Vol. 4
. —
P. 11
. —
doi
:
. —
. —
PMC
.
(неопр.)
. Дата обращения: 1 июня 2019.
1 июня 2019 года.
Pille Hallast, Anastasia Agdzhoyan, Oleg Balanovsky, Yali Xue, Chris Tyler-Smith
.
от 8 декабря 2019 на
Wayback Machine
, 2019
Juhasz
et al. New clustering methods for population comparison on paternal lineages. Molecular genetics and genomics 290 (2) : 767-784, (2014)
Tatiana M. Karafet
et al.
, 08 November 2018
(неопр.)
. Дата обращения: 28 сентября 2017.
1 июня 2019 года.


_2017.png)