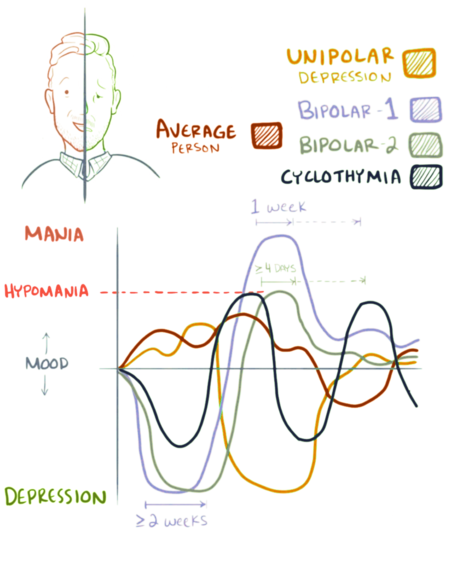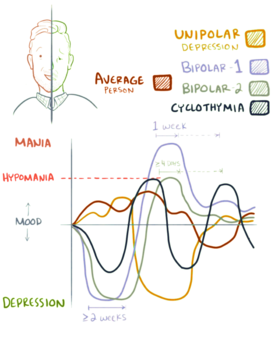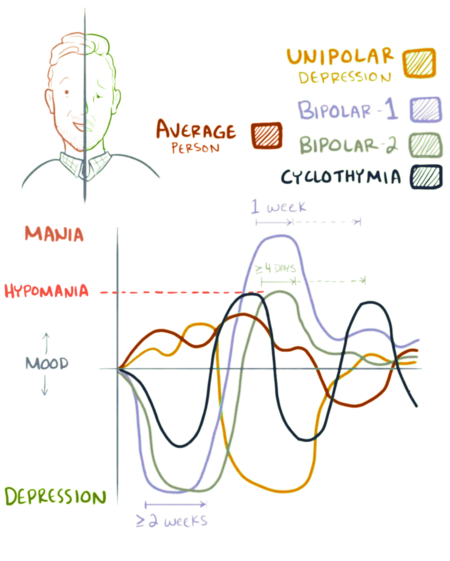
Биполярное расстройство
- 1 year ago
- 0
- 0


Полярное тельце веретена ( ПТВ ) — центр организации микротрубочек , грибной эквивалент центросомы клеток животных. В отличие от центросомы в ПТВ нет центриолей . У дрожжей S. cerevisiae под электронным микроскопом оно выглядит как электронно-плотная многослойная структура, встроенная в оболочку ядра. Помимо основной функции (центр организации микротрубочек), полярное тельце веретена опосредованно участвует в сегрегации хромосом, расположении ядер в клетке, кариогамии и ориентации веретена деления. Кроме того, оно является местом восприятия сигналов MEN ( англ. mitotic exit network ) пути, и, возможно участвует в формировании стенки сумки споры S. cerevisiae .
Структура ПТВ была тщательно изучена с помощью электронной микроскопии у ряда дрожжей ( , S. pombe , ) и мицелиального гриба Ashbya gossypii . Показано, что ПТВ представляет собой трехслойную немембранную структуру, которая в случае неделящегося ядра находится в цитоплазме, плотно примыкает к ядерной оболочке и принимает участие в формировании цитоплазматических микротрубочек, а во время деления ядра (митоз, мейоз) встраивается в ядерную оболочку и принимает участие в формировании веретена деления. ПТВ S. cerevisiae представляет собой цилиндрическую органеллу, которая состоит из трех дисков или пластин темноокрашенного материала. Выделяют следующие слои ПТВ (при движении от цитоплазмы к ядру):
С одной стороны центральная пластина полярного тельца веретена контактирует с электронноплотным районом ядерной оболочки, называемым полумостом ( англ. half-bridge ). Это место сборки нового полярного тельца, но кроме того он участвует в кариогамии. Ядерная и цитоплазматические стороны полумоста не эквивалентны. Два мембранных белка, Kar1p и Mps3p, расположенные в полумосте, необходимы для формирования и поддержания его структуры и маркируют его стороны. Оба белка связаны с Cdc31p, дрожжевым гомологом , который необходим для поддержания целостности полумоста. Ещё один компонент полумоста, Sfi1p, способен связываться с Cdc31p при помощи множества консервативных Cdc31-связывающих сайтов. Kar1p участвует в соединении полумоста с ядром ПТВ посредством взаимодействия с Bbp1p, а также играет роль в реорганизации ПТВ во время G 1 -фазы . Центральная пластинка и внутренний слой 2 в свою очередь состоят из отдельных высокоупорядоченных слоев.
Тщательный анализ размера и структуры ПТВ у S. cerevisiae показал, что ПТВ — динамичная органелла. В гаплоидных клетках ПТВ разрастаются в диаметре от 80 нм в стадии G, клеточного цикла до 110 нм в митозе, хотя высота ПТВ (расстояние от внутренней до наружной пластины) остается постоянной и составляет примерно 150 нм. Диаметр ПТВ увеличивается с увеличением содержания в клетке ДНК . В диплоидной клетке диаметр ПТВ составляет 160 нм, а в тетраплоидной — в два раза больше. Увеличение диаметра ПТВ увеличивает его способность к образованию микротрубочек, что является важным для сегрегации хромосом . Как регулируется размер ПТВ, пока неизвестно. Структура ПТВ изменяется на стадии G 1 , при подготовке к удвоению и созреванию и в процессе второго деления мейоза при инициации формирования спор.
Полярное тельце веретена является единственным местом зарождения микротрубочек у многоядерного мицелиального аскомицетного гриба , близкого родственника почкующихся дрожжей . Анализ ПТВ A. gossypii с помощью электронной микроскопии выявил многослойную структуру, сходную с полярным тельцем S. cerevisiae , но с выраженными различиями на цитоплазматической стороне. До 6 перпендикулярных и тангентальных цитоплазматических микротрубочек отходят от наружной пластинки. Перпендикулярные и тангентальные цитоплазматические микротрубочки соответствуют коротким микротрубочкам, ассоциированным с кортексом. Каждое ПТВ порождает свой собственный пучок цитоплазматических микротрубочек, и отсутствие перекрытия пучков микротрубочек соседних ядер объясняет автономную осцилляцию ядер, наблюдаемую в многоядерной гифе A. gossypii .
Молекулярная масса диплоидного ПТВ, включая микротрубочки и ассоциированные с микротрубочками белки, достигает 1-1,5 ГДа , сердцевина полярного тельца веретена составляет 0,3-0,5 ГДа. ПТВ содержит, по крайней мере, 30 разных белков. Однако, к настоящему времени идентифицировано только 17 компонентов митотического ПТВ. ПТВ образовано структурными белками, γ-тубулиноподобными белками и компонентами MEN. Делеция или мутация гена, контролирующего синтез любого из этих белков, приводит к множественным дефектам структуры и функции ПТВ: арест клеточного цикла, однополюсное веретено, дефекты сборки ПТВ, мутации регуляторов кариогамии, нарушение положения ядер и др.
Spc42 — особый спирально закрученный белок, который образует центр ПТВ. Примерно 1000 молекул Spc42 собрано в триммеры димеров Spc42, формируя гексагональную решетку, видимую на крио-ЭМ в слое IL2. Суперэкспрессия Spc42 приводит к формированию суперпластины, необходимой для действия cis- и /nms-факторов, которые задействованы в удвоении ПТВ на стадии G 1 клеточного цикла.
На N-конце белок Spc42 связан с Spc110 и Spc29, двумя другими спирально закрученными белками, которые локализованы на ядерной стороне ПТВ. С-Концы Spc110 локализованы в центральной пластинке и связаны с Spc29 и кальций-связывающим белком кальмодулином (Cmdl). У дрожжей — гипотетическая функция кальмодулина в ПТВ — регуляция связывания белка Spcll0 с Spc29. Белок Spc29 является линкером между внутренней и центральной пластинкой и связан с Spc42 и Spc110, хотя по некоторым данным Spc110 может непосредственно взаимо¬действовать с Spc42 .
N-Конец Spc110 локализован во внутренней пластинке и связан непосредственно с Spc98, одним из двух похожих у-тубулин-связанных белков, необходимых для образования микротрубочек. С-Конец Spc42 обращен в цитоплазму и связан с С-концом Cnm67. Сходно с Spc110, спирально закрученный участок Cnm67 способствует его димеризации и функционированию между внутренними слоями IL2 и IL1. N-Конец белка Cnm67 связан с наружной пластиной ПТВ и белком Nudl, необходимым для выхода из митоза. Другой спирально закрученный белок Spc72 также обнаружен в наружной пластинке. Spc72 связан с Nudl и с компонентами γ-тубулинового комплекса.
По крайней мере, 9 компонентов сигнального пути сети MEN, которые контролируют положение веретена деления и некоторые поздние события митоза , связаны с ПТВ. Локализация белков MEN в полярном тельце веретена важна для их функционирования. Белки, которые регулируют положение веретена деления, могут быть также обнаружены в ПТВ. Динеин , а также фракция кортикально-связанного белка Kar9 локализованы в ПТВ и связаны с микротрубочками.
Полярное тельце веретена не синтезируется заново, а удваивается каждый раз на стадии G 1 клеточного цикла . Этот процесс очень важен для образования биполярного веретена деления . Процесс удвоения ПТВ можно разделить на три этапа: первая происходит рано на стадии G 1 , когда на краю полумоста образуется сателлитный материал. В ходе второго этапа полумост удлиняется, а его цитоплазматическая и ядерные стороны сливаются. В то же время сателлит образует новую несущую пластинку, слоистую структуру, которая по составу схожа с цитоплазматической частью ПТВ. Эта пластинка встраивается в ядерную оболочку, и на её основе происходит достраивание нового ПТВ, сначала цитоплазматической, а затем и ядерной части. В конце G 1 в дрожжевой клетке содержатся два расположенных рядом ПТВ, соединённых мостом. Затем связывающий их мост разрывается, и два полярных тельца веретена перемещаются на противоположные стороны ядерной оболочки .
Диплоидные клетки дрожжей S. cerevisiae подвергаются двум делениям мейоза с образованием гаплоидного потомства в условиях азотного голодания. В процессе деления происходит расхождение хромосом, и потомство получает только одну копию из всех 16 хромосом. Мейотическое веретено аналогично по структуре митотическому веретену и образуется в процессе первого деления мейоза. Морфологически полярное тельце веретена деления мейоза I идентично ПТВ митотического веретена деления, но больше его в диаметре. Предположительно, все компоненты ПТВ важны для митотического ПТВ. Между делениями мейоза ПТВ должно удвоиться, так как формируются два биполярных веретена в стадии мейоза II. Любопытно, что удвоение происходит в отсутствие ядерного деления (митоз у грибов и ядерная оболочка сохраняется на протяжении всего цикла), то есть два мейотических веретена мейоза II формируются в одном ядре. Вслед за последовательным удвоением и формированием веретена мейоза II ПТВ подвергаются морфологическим изменениям. Белок наружной пластины Spc72 удаляется и заменяется на два мейотических белка ПТВ, Мрс54 и Spo21/Mpc70, которые формируют мейотическую пластину .
|
Эндомембранная система
|
|
|---|---|
| Цитоскелет | |
| Эндосимбионты | |
| Другие внутренние органеллы | |
| Внешние органеллы | |