Гаплогруппа U4a1
— гаплогруппа митохондриальной ДНК человека. U4a1 возможно, может быть прослежена до неолита в Восточной Европе и на Кавказе. Эти линии мтДНК, вероятно, были вовлечены в позднеледниковую экспансию из восточноевропейских
рефугиумов (мест выживания)
после LGM (ледниковых периодов), (примерно за 12 000–19 000 лет
до настоящего времени
)
.
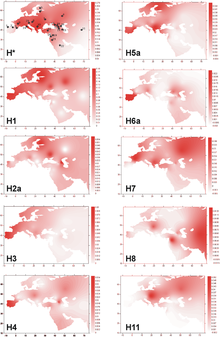
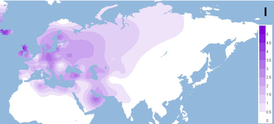
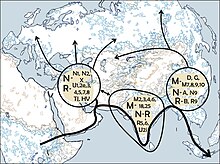
Гаплогруппа U4a1 в значительной степени характеризуется восточноевропейским и западносибирским распространением, обнаруживаемым с наибольшей частотой 7–21% в таких популяциях, как марийцы, чуваши и кеты. 4,5% в Волго-Уральском регионе (марийцы, коми-зыряне, коми-пермяки, мордвины, удмурты, чуваши, татары и башкиры); 1% в центральной и восточной Европе (русские, белорусы, украинцы, поляки и словаки); 0,9% в северной Европе (финны, карелы, эстонцы, латыши и литовцы); 0.5% в центральной и западной Европе (британцы, немцы, чехи, австрийцы, швейцарцы, боснийцы, словенцы, итальянцы, французы, испанцы и португальцы). Интересно, что среди восточных славян (русских и белорусов) выявлен единственный специфический субклад U4a1a1, тогда как у западных славян обнаружены остальные субклады. Оценка времени слияния полных геномов U4a1 составила 14 650 ± 2 400 YBP. Это значение близко к значениям (около 10 000–14 000 YBP), рассчитанным на основе данных первого гипервариабельного сегмента (HVS1) для центральноевропейского (германоязычного) населения, но не для балтийских финно-угров и поволжских народов (около 20 000–22 000 YBP)(Tambets et al. 2003)
. По результам исследований мтДНК
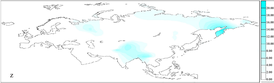
калашей
, в их генетическом составе гаплогруппа U4a1 составляет 21,6% —
Читрал (округ)
,
Хайбер-Пахтунхва
,
Пакистан
.
U4a1-a
C16134T возник 11100 лет до настоящего времени, БОП 8400 до настоящего времени
U4a1a
T961C возник 8400 лет до настоящего времени, БОП 5800 до настоящего времени
U4a1a1
A955AC * A955C возник 5800 лет до настоящего времени, БОП 2600 до настоящего времени
U4a1a1a
G12618 возник 2600 до настоящего времени, БОП 1200 до настоящего времени
U4a1a1a1
T8167C возник 1200 до настоящего времени, БОП 200 до настоящего времени
U4a1a1b
C12588T возник 2600 до настоящего времени, БОП 1350 до настоящего времени
U4a1a1c
A13986T возник 2600 до настоящего времени, БОП 550 до настоящего времени
U4a1a1d
C3576T возник 2600 до настоящего времени, БОП 900 до настоящего времени
U4a1a1e
A4562G возник 2600 до настоящего времени, БОП 900 до настоящего времени
U4a1a1f
G12308A! возник 2600 до настоящего времени, БОП 550 до настоящего времени
U4a1a1g
A7439G возник 2600 до настоящего времени, БОП 800 до настоящего времени
U4a1a1h
C14884T возник 2600 до настоящего времени, БОП 1750 до настоящего времени
U4a1a1j
A189G возник 2600 до настоящего времени, БОП 1350 до настоящего времени
U4a1a2
T5250C * G12630A возник 5800 лет до настоящего времени, БОП 2900 до настоящего времени
U4a1a2a
A3447G возник 2900 лет до настоящего времени, БОП 1000 до настоящего времени
U4a1a3
T6216C * C15544A возник 5800 лет до настоящего времени, БОП 3000 до настоящего времени
U4a1a3a
T6216C возник 3000 лет до настоящего времени, БОП 1650 до настоящего времени
U4a1a3a1
A15112G возник 1650 лет до настоящего времени, БОП 225 до настоящего времени
U4a1a4
A7052G возник 5800 лет до настоящего времени, БОП 900 до настоящего времени
U4a1a5
T8258A возник 5800 лет до настоящего времени, БОП 3900 до настоящего времени
U4a1a6
G16310A возник 5800 лет до настоящего времени, БОП 2300 до настоящего времени
U4a1a7
A479T возник 5800 лет до настоящего времени, БОП 2300 до настоящего времени
U4a1a8
A7439G возник 5800 лет до настоящего времени, БОП 2100 до настоящего времени
U4a1a8a
C16111T возник 2100 лет до настоящего времени, БОП 225 до настоящего времени
U4a1a9
T11722C возник 5800 лет до настоящего времени, БОП 3700 до настоящего времени
U4a1a9a
T16086C возник 3700 лет до настоящего времени, БОП 600 до настоящего времени
U4a1-a1
T3548C возник 8400 лет до настоящего времени, БОП 100 до настоящего времени
U4a1-a10
C3576T
U4a1-a11
A10532G возник 8400 лет до настоящего времени, БОП 6300 до настоящего времени
U4a1-a11a
C14792G возник 6300 лет до настоящего времени, БОП 5200 до настоящего времени
U4a1-a11a1
T5981A возник 1950 лет до настоящего времени, БОП 250 до настоящего времени
U4a1-a12
C16286A возник 8400 лет до настоящего времени, БОП 3700 до настоящего времени
U4a1-a14
T8167C возник 8400 лет до настоящего времени, БОП 3300 до настоящего времени
U4a1-a14a
G12618A возник 3300 лет до настоящего времени, БОП 1150 до настоящего времени
U4a1-a2
A13773G возник 8400 лет до настоящего времени, БОП 2800 до настоящего времени
U4a1-a2a
A7025T возник 2800 лет до настоящего времени, БОП 1100 до настоящего времени
U4a1-a2a1
A2792G возник 1100 лет до настоящего времени, БОП 225 до настоящего времени
U4a1-a3
T15341C возник 8400 лет до настоящего времени, БОП 3400 до настоящего времени
U4a1-a3a
T5492C возник 3400 лет до настоящего времени, БОП 550 до настоящего времени
U4a1-a4
T16311C! возник 8400 лет до настоящего времени, БОП 6300 до настоящего времени
U4a1-a4a
A7299G возник 6300 лет до настоящего времени, БОП 1850 до настоящего времени
U4a1-a4a1
C8275T возник 1850 лет до настоящего времени, БОП 425 до настоящего времени
U4a1-a4b
C8393T возник 6300 лет до настоящего времени, БОП 2100 до настоящего времени
U4a1-a5
T14635C возник 8400 лет до настоящего времени, БОП 4400 до настоящего времени
U4a1-a5a
T72C возник 4400 лет до настоящего времени, БОП 2600 до настоящего времени
U4a1-a5a1
G15773A возник 2600 лет до настоящего времени, БОП 1100 до настоящего времени
U4a1-a5a2
A12950G возник 2600 лет до настоящего времени, БОП 350 до настоящего времени
U4a1-a6
A13731G возник 8400 лет до настоящего времени, БОП 1450 до настоящего времени
U4a1-a7
C9518T возник 8400 лет до настоящего времени, БОП 5700 до настоящего времени
U4a1-a7a
G8392A возник 5700 лет до настоящего времени, БОП 2500 до настоящего времени
U4a1-a7aa
T14502C возник 2500 лет до настоящего времени, БОП 300 до настоящего времени
U4a1-a8
C15031T возник 8400 лет до настоящего времени, БОП 6000 до настоящего времени
U4a1-a8a
A13722G возник 6000 лет до настоящего времени, БОП 4500 до настоящего времени
U4a1-a9
G3531T
U4a1b
A745G * C3204T возник 8400 лет до настоящего времени, БОП 4000 до настоящего времени
U4a1b1
C296T возник 4000 лет до настоящего времени, БОП 2100 до настоящего времени
U4a1b1b
C16150A возник 2100 лет до настоящего времени, БОП 600 до настоящего времени
U4a1b1c
T11009C возник 2100 лет до настоящего времени, БОП 1200 до настоящего времени
U4a1b1c1
T14971C возник 1200 лет до настоящего времени, БОП 450 до настоящего времени
U4a1b1d
C14368T возник 2100 лет до настоящего времени, БОП 1350 до настоящего времени
U4a1b1d1
G7853A возник 1350 лет до настоящего времени, БОП 1150 до настоящего времени
U4a1b2
C16301T возник 4000 лет до настоящего времени, БОП 2100 до настоящего времени
U4a1b2a
C7816T возник 2100 лет до настоящего времени, БОП 1050 до настоящего времени
U4a1c
G8155A * A13158G * T14110C * C16234T возник 8400 лет до настоящего времени, БОП 1350 до настоящего времени
U4a1c1
C16221T возник 1350 лет до настоящего времени, БОП 600 до настоящего времени
U4a1d
C198T возник 8400 лет до настоящего времени, БОП 4800 до настоящего времени
U4a1d1
A4206G возник 4800 лет до настоящего времени, БОП 3300 до настоящего времени
U4a1d1a
C9335A возник 3300 лет до настоящего времени, БОП 1850 до настоящего времени
U4a1d1a1
T16154C возник 1850 лет до настоящего времени, БОП 500 до настоящего времени
U4a1d1a2
G1438A возник 1850 лет до настоящего времени, БОП 500 до настоящего времени
U4a1d1a3
T8022C возник 1850 лет до настоящего времени, БОП 500 до настоящего времени
U4a1g
T504C возник 11100 лет до настоящего времени, БОП 1800 до настоящего времени
Палеогенетика
Мезолит
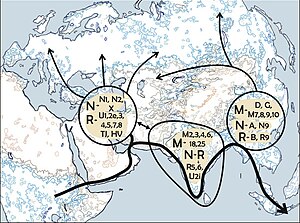
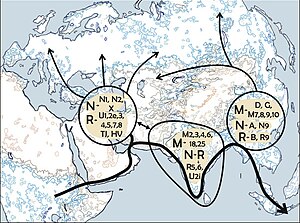
Генетическое положение кластера "Сиделькино":(
Песчаница
,
Попово
,
Сиделькино
,
Минино
,
Оленеостровcкий
, Васильевка I,
Караваиха
, Языково I) среди других древних евразийских охотников-собирателей
.
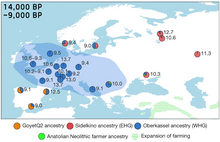
Генетическая родословная охотников-собирателей в Европе между 14 и 9 тыс. лет назад, ареалом западных охотников-собирателей (WHG) выделен синим цветом. Отдельные числа соответствуют датам калиброванных образцов.
.
Примерно, 15–13 тыс. лет назад предки
впервые появились в Центральной Европе, что совпадает с периодом
Бёллингского потепления
. Это повышает вероятность того, что формирование кластера "Сиделькино" могло быть результатом расширения и смешения популяций под влиянием резкого потепления
. Кластер "Сиделькино" (Posth 2023 et.al) включает могильники Песчаница, Попово, Сиделькино, Минино, Оленеостровcкий, Васильевка I, Караваиха, Языково I.
PES001 (10,785—10,626 лет до н. э.) Песчаница 1 у озера
Лача
,
Архангельская обл.
,
Россия
Y-хромосомная гаплогруппа R1a5-YP1301 (под R1a1b~-YP1272))
MIN11 (8,671 ± 48 лет до н. э.), MIN3 (7472±52 лет до н. э.), MIN8 (6,450 – 5,800 лет до н. э.) у озера Кубенское в деревне
Минино
,
Вологодская область
,
Россия
MN2002(ok. 8,595 лет до н. э.) Y-хромосомная гаплогруппа R1a1 M459, MNN005 (ok. 8,563 лет до н. э.), MNN003 (ok. 5,610 лет до н. э.), Y-хромосомная гаплогруппа CT у озера Кубенское в деревне
Минино
,
Вологодская область
,
Россия
I4440 (ZVEJ21) (5,220-5,039 лет до н. э.) Y-хромосомная гаплогруппа I-FTA37476 (I2a) могильник
Звейниеки
, на севере Латвии, на северном берегу озера Буртниекс в устье реки Руя.
ZV162 (4,470 ± 72 лет до н. э.) и Zv317 (3890 ± 67 лет до н. э.)
Звейниеки
,
Латвия
Примерно 8 тыс. лет назад, когда
начали распространяться по Европе, максимально увеличилось составляющая часть ANF среди секвенированный геномов охотников-собирателей этого периода
.
Kivisaare2 (4635 лет до н. э.) и Kivisaare3 (4730-4540 лет до н. э.), могильник Kivisaare, южная часть
Эстонии
в деревне Лалси, примерно в 1,5 км от реки Пылтсамаа и в 6 км от нынешнего берега озера Võrtsjärv3
.
I5892 (5301-4982 лет до н. э.) Y-хромосомная гаплогруппа R-L754 (R1b) и I5875 (5291-5060 лет до н. э.)Y-хромосомная гаплогруппа I-L703* (I2a),
Дериевка
,
Украина
.
NEO508 (ок. 5106 лет до н. э.) и NEO498 (ок. 4354 лет до н. э.)
Вовниги
,
Украина
.
I4627 (ZVEJ26) (4251-3976 лет до н. э.) Y-хромосомная гаплогруппа R-M73* (R1b) R1b1a1a(xR1b1a1a2), I4435 (ZVEJ13)(4146-3960 лет до н. э.) и I4436 (ZVEJ14)(4260-4050 лет до н. э.) могильник
Звейниеки
, на севере Латвии, на северном берегу озера Буртниекс в устье реки Руя
.
Koljala1 (4230–3810 лет до н. э.) Kõljala,
Эстония
.
NEO185 (ок. 5078 лет до н. э.) со стоянки Сахтыш IIA, (
Тейковский район
Ивановской области
) расположена на низком левом берегу небольшого озеровидного расширения старого русла р. Койки, примерно в 2 км к югу от ее истока из древнего озера Сахтыш,
Россия
.
BOO49-3 (ок. 1500 лет до н. э.) и BOO57-1 (ок. 1500 лет до н. э.),
Большой Олений Остров
на Кольском полуострове. Этот участок находится в пределах территории, в настоящее время заселенной саамами,
Россия
.
C1703у (TGB M2-2) (ок. 2250 лет до н. э.) могильник Tuoganbai
уезда Каба
в округе Алтай Синьцзян-Уйгурского автономного района,
Китай
.
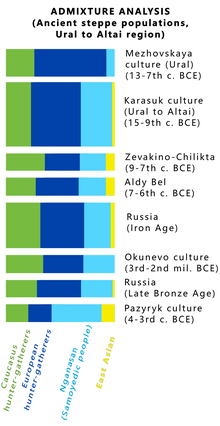
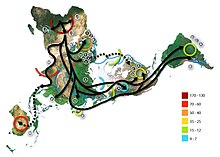
Упрощенная схема результатов анализа ADMIXTURE для древних степных популяций кочевников Евразийской степи (от Урала до Алтая), в том числе представителей Карасукской культуры. Голубым отмечен
нганасанский
компонент (
самодийцы
), синиим - европейские охотники-собиратели, желтый - восточно-азиатский компонент, зеленым - кавказские охотники-собиратели. (см. полную версию результатов
количество предковых популяций К варьируется от 2 до 15) .
I3861 (1872-1684год до н. э.), Ой-Джайляу III, Урочище Ой-Джайляу, Курдайский район Джамбулской области, в Чу-Илийских горах,
Казахстан
.
Malyarchuk; et al. (2008). .
Science
.
25
(8): 1651—1658.
DOI
: .
2013
Brandt; et al. (2013). .
Science
.
342
: 257–261.
DOI
: .
Der Sarkissian C; et al. (2013). “Ancient DNA reveals prehistoric gene-flow from siberia in the complex human population history of North East Europe”.
.
DOI
: .
Buzhilova A.P.; et al. (2016). “A reconstruction of the lifestyle of early humans by natural-science methods”.
Вестник Российской академии наук
.
86
(4): 297–304.
DOI
: .
Csősz, A., Szécsényi-Nagy, A., Csákyová, V.; et al. (2016). .
Nature
.
DOI
: .
Fu, Q., Posth, C., Hajdinjak, M.; et al. (2016). .
Nature
.
534
: 200—205.
DOI
: .
PMID
.
Unterländer, M., Palstra, F., Lazaridis, I.; et al. (2017). “Ancestry and demography and descendants of Iron Age nomads of the Eurasian Steppe”.
Nature Communications
.
8
.
DOI
: .
2018
Damgaard, P.d.B., Marchi, N., Rasmussen, S.; et al. (2018). .
Nature
.
557
: 369—374.
DOI
: .
Emery; et al. (2018). .
Journal of Archaeological Scienc
: 200—209.
DOI
: .
Günther T, Malmström H, Svensson EM, Omrak A, Sanchez-Quinto F, KKilinc GM; et al. (2018). .
PLoS Biology
.
DOI
: .
Siddiqi MH, Rakha A, Khan K, Akhtar T. (2021 July). .
Mitochondrial DNA Part B
.
6
(8): 2410–2414.
DOI
: .
PMC
.
PMID
.
Проверьте дату в
|date=
(
справка на английском
)
Kumar; et al. (2022). .
Science
.
376
(6588): 62–69.
DOI
: .
Allentoft; et al. (2022). .
bioRxiv
(022.05.04.490594).
DOI
: .
2023
Posth, C., Yu, H., Ghalichi, A; et al. (2023). “Palaeogenomics of Upper Palaeolithic to Neolithic European hunter-gatherers”.
Nature
.
615
: 117—126.
DOI
: .
Engovatova; et al. (2023). “Ancient DNA of the Bearers of the Fatyanovo and Abashevo Cultures (Concerning Migrations of the Bronze Age people in the Forest Belt on the Russian Plain)”.
.
2
: 207–228.
DOI
: .
Rodríguez-Varela, Ricardo; et al. (2023). “The genetic history of Scandinavia from the Roman Iron Age to the present”.
186,1: 32-46.e19.
DOI
: .