Лан (город)
- 1 year ago
- 0
- 0

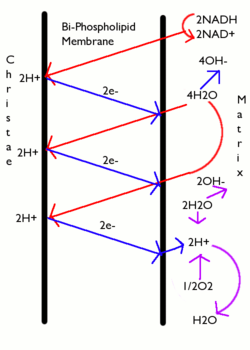
Ядерно-цитоплазматический транспорт — это материальный обмен между клеточным ядром и цитоплазмой клетки, который осуществляется через ядерные поры . Ядерно-цитоплазматический транспорт можно разделить на две категории: активный транспорт , требующий затрат энергии, а также специальных белков- рецепторов и транспортных факторов, и пассивный транспорт , протекающий путём простой диффузии молекул через канал ядерной поры.
Молекулы небольших размеров ( ионы , метаболиты , мононуклеотиды и т. д.) способны пассивно перемещаться между цитоплазмой и ядром за счёт диффузии . Проводимость ядерных пор для молекул разных размеров различна: белок массой менее 15 кДа проникает в ядро быстрее, чем белок массой более 30 кДа. Белковые молекулы массой более 40 кДа, по-видимому, вообще не могут пассивно проходить через ядерные поры. Тем не менее, даже маленькие белки, которые выполняют функции в ядре, такие как гистоны , в большинстве случаев переносятся туда в комплексе с белками-переносчиками, а не проникают пассивно .

Путём активного транспорта через ядерные поры могут проходить гораздо более крупные молекулы и целые надмолекулярные комплексы. Так, рибосомные субчастицы массой до нескольких мегадальтон транспортируются из ядра в цитоплазму через ядерные поры, и нет никаких оснований предполагать, что процесс транспорта сопровождается частичной разборкой этих субчастиц. Системы активного транспорта обеспечивают весь макромолекулярный обмен между ядром и цитоплазмой. Молекулы РНК, синтезируемые в ядре, поступают через поры в цитоплазму, а в ядро попадают белки, участвующие в ядерном метаболизме. Причём одни белки должны поступать в ядро конститутивно (например, гистоны ), а другие в ответ на определенные стимулы (например, транскрипционные факторы ).
Центральную роль в активном ядерно-цитоплазматическом транспорте белков играет малая ГТФаза Ran, относящаяся к семейству и служащая для сопряжения транспорта с гидролизом ГТФ , что придает процессу необратимость (снабжает его энергией). Ran может образовывать комплекс с ГТФ или с ГДФ . Как и другие Ras-подобные ГТФазы, Ran состоит из небольшого G-домена и содержит две поверхностных петли, switch-I и switch-II, которые меняют конформацию в зависимости от связанного нуклеотида . Собственно акт гидролиза ГТФ осуществляется непосредственно Ran. Фактор обмена нуклеотидов (англ., G TPase Е xchange F actor, GEF) для Ran, хроматин-связывающй белок RCC1, локализован исключительно в ядре, а активаторы ГТФазной активности (англ., G TPase A ctivation P rotein, GAP) RanGAP1 и некоторые другие белки — исключительно в цитоплазме. Эта асимметричная локализация приводит к формированию градиента: в ядре находится преимущественно ГТФ-связанная форма Ran, в цитоплазме, наоборот, ГДФ-связанная. Ran используется для снабжения энергией как процессов импорта, так и процессов экспорта различных субстратов, а вся схема носит название Ran-цикла (англ., Ran-cycle). Ключевыми стадиями Ran-цикла являются гидролиз ГТФ в цитоплазме и обмен ГДФ на ГТФ в ядре.
Ядерные белки содержат особые аминокислотные последовательности, отвечающие за их локализацию в ядре, — сигналы ядерной локализации ( англ. N uclear L ocalization S ignal, NLS ). Сигналы ядерной локализации — это места узнавания молекулы транспортными факторами — транспортинами (кариоферинами), которые и переносят белок-субстрат в ядро. Одним из первых был открыт так называемый классический, или основной сигнал ядерной локализации (cNLS). Он представляет собой один или два кластера положительно заряженных аминокислотных остатков ( аргинин и лизин ) и распознаётся кариоферинами-α . Транслокация белков в ядро, в отличие от транслокации в митохондрии и эндоплазматический ретикулум , не сопровождается отщеплением этой сигнальной последовательности и разворачиванием полипептидной цепи. Помимо связывания субстрата, транспортины и их комплексы способны заякориваться на ядерной поре и связывать Ran.
Рассмотрим механизм поступления субстратов в ядро на примере импорта NLS-содержащих белков. Первой стадией транспортировки является узнавание субстрата транспортинами, в данном случае комплексом импортинов-α/β (транспортины участвующие в транспорте в ядро называются импортинами, а из ядра — экспортинами ). Затем образовавшийся комплекс заякоривается на белках ядерной поры с цитоплазматической стороны и транслоцируется через канал в ядро, где с ним связывается Ran-ГТФ, что вызывает диссоциацию комплекса и высвобождение груза. После чего импортины в комплексе с Ran-ГТФ направляются обратно в цитоплазму, где Ran под действием RanGAP1 гидролизует ГТФ (ГТФ => ГДФ + PO 4 3- ). Комплекс Ran-ГДФ-импортины α/β нестабилен и диссоциирует. Ran-ГДФ поступает обратно в ядро при помощи собственного переносчика, димерного белка NTF2. В ядре под действием белка RanGEF, ГДФ в активном центре Ran заменяется на ГТФ и цикл, тем самым, замыкается.
Теперь рассмотрим механизм экспорта из ядра на примере белков, содержащих сигналы ядерного экспорта (англ., N uclear E xport S ignal, NES). Для этих сигнальных последовательностей характерно высокое содержание гидрофобных аминокислот. Первой стадией транспортировки здесь также является рецепция субстрата специфическим экспортином Crm1 (англ., C hromosome R egion M aintenance) и образование комплекса. Главным отличием механизмов экспорта является тот факт, что в состав транслоцирующегося комплекса в случае экспорта помимо субстрата и Crm1 входит и Ran-ГТФ, то есть сопряжение с циклом Ran происходит на стадии транслокации, а не на стадии реимпорта рецептора. После прохождения через ядерную пору в цитоплазму Ran расщепляет ГТФ, комплекс теряет стабильность и диссоциирует, высвобождая груз.