Interested Article - Цианобактерии

- 2020-12-01
- 1
Цианобакте́рии , или сине-зелёные во́доросли , или циане́и ( лат. Cyanobacteria , от греч. κυανός — сине-зелёный) — отдел крупных грамотрицательных бактерий , способных к фотосинтезу , сопровождающемуся выделением кислорода .
Эволюционное и систематическое положение
Цианобактерии наиболее близки к древнейшим микроорганизмам, постройки которых ( строматолиты , возраст более 3,5 млрд лет) обнаружены на Земле. Это единственные бактерии, способные к оксигенному фотосинтезу . Цианобактерии относятся к наиболее сложно организованным и морфологически дифференцированным прокариотам.
Один из видов древних цианобактерий, вступив в эндосимбиоз с предком архепластид , дал начало их хлоропластам (по молекулярным данным , хлоропласты в этой группе приобретались лишь однажды, а ближайший современный родственник их предка — цианобактерия ) . Кроме этого, известен ещё только один случай первичного появления пластид — у амёб рода (вступивших в симбиоз с другой цианобактерией и значительно позже) .
Цианобактерии являются объектом исследования как альгологов (как организмы, физиологически схожие с эукариотическими водорослями), так и бактериологов (как прокариоты ). Сравнительно крупные размеры клеток и сходство с водорослями было причиной их рассмотрения ранее в составе растений («сине-зелёные водоросли»). За это время было альгологически описано более 1000 видов в почти 175 родах. Бактериологическими методами в настоящее время подтверждено существование не более 400 видов. Биохимическое , молекулярно-генетическое и филогенетическое сходство цианобактерий с остальными бактериями в настоящее время подтверждено солидным корпусом доказательств.
Жизненные формы и экология
.jpg)
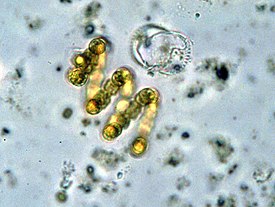
В морфологическом отношении цианопрокариоты — разнообразная и полиморфная группа. Общие черты их морфологии заключаются только в отсутствии жгутиков и наличии клеточной стенки ( гликокаликс , состоящий из пептидогликана ). Поверх слоя пептидогликана толщиной 2—200 нм имеют наружную мембрану. Ширина или диаметр клеток варьируется от 0,5 мкм до 100 мкм. Цианобактерии — одноклеточные , и колониальные микроорганизмы. Отличаются выдающейся способностью адаптировать состав к спектральному составу света, так что цвет варьируется от светло-зелёного до тёмно-синего. Некоторые азотфиксирующие цианобактерии способны к дифференцировке — формированию специализированных клеток: гетероцист и гормогониев . Гетероцисты выполняют функцию азотфиксации , в то время как другие клетки осуществляют фотосинтез.
Морские и пресноводные, почвенные виды, участники симбиозов (например, в лишайнике ). Составляют значительную долю океанического фитопланктона . Способны к формированию толстых бактериальных матов . Некоторые виды токсичны (выделяют такие токсины , как anatoxin-a, anatoxin-as, аплизиатоксин , цилиндроспермопсин, домоевую кислоту , микроцистин, нодулярин, неосакситоксин, сакситоксин ) и условно-патогенны (например, Anabaena ). Главные участники цветения воды , которое вызывает массовые заморы рыбы и отравления животных и людей. Уникальное экологическое положение обусловлено наличием двух трудносочетаемых способностей: к фотосинтетической продукции кислорода и фиксации атмосферного азота (у 2/3 изученных видов).
Деление бинарное в одной или нескольких плоскостях, . Жизненный цикл у одноклеточных форм при оптимальных условиях роста — 6—12 часов.
Биохимия и физиология
Цианобактерии обладают полноценным , характерным для кислородвыделяющих фотосинтетиков. Фотосинтетическая электронтранспортная цепь включает фотосистему (ФС) II , b 6 f-цитохромный комплекс и ФСI . Конечным акцептором электронов служит ферредоксин , донором электронов — вода , расщепляемая в , аналогичной таковой высших растений. Светособирающие комплексы представлены особыми пигментами — фикобилинами , собранными (как и у красных водорослей ) в фикобилисомы . При отключении ФСII способны к использованию других, нежели вода, экзогенных доноров электронов: восстановленных соединений серы , органических соединений в рамках с участием ФСI. Однако эффективность такого пути фотосинтеза невелика, и он используется преимущественно для переживания неблагоприятных условий.
Цианобактерии отличает чрезвычайно развитая система внутриклеточных выпячиваний цитоплазматической мембраны (ЦПМ) — тилакоидов ; высказаны предположения о возможном существовании у них системы тилакоидов, не связанных с ЦПМ, что до сих пор считалось невозможным у прокариот . Накопленная в результате фотосинтеза энергия используется в темновых процессах фотосинтеза для производства органических веществ из атмосферного CO 2 .
Большинство цианобактерий — облигатные фототрофы , которые, однако, способны к непродолжительному существованию за счёт расщепления накопленного на свету гликогена в окислительном пентозофосфатном цикле и в процессе гликолиза (достаточность одного гликолиза для поддержания жизнедеятельности подвергается сомнению).
Азотфиксация обеспечивается ферментом нитрогеназой , который отличается высокой чувствительностью к молекулярному кислороду. Поскольку кислород выделяется при фотосинтезе, в эволюции цианобактерий реализованы две стратегии: пространственного и временного разобщения этих процессов. У одноклеточных цианобактерий пик фотосинтетической активности наблюдается в светлое, а пик нитрогеназной активности — в тёмное время суток. Процесс регулируется генетически на уровне транскрипции ; цианобактерии являются единственными прокариотами, у которых доказано существование циркадных ритмов (причём продолжительность суточного цикла может превышать продолжительность жизненного цикла). У нитчатых цианобактерий процесс азотфиксации локализован в специализированных терминально-дифференцированных клетках — гетероцистах , отличающихся толстыми покровами, которые препятствуют проникновению кислорода. При недостатке связанного азота в питательной среде в колонии насчитывается 5—15 % гетероцист. ФСII в гетероцистах редуцирована. Гетероцисты получают органические вещества от фотосинтезирующих членов колонии. Накопленный связанный азот накапливается в гранулах или экспортируется в виде глутаминовой кислоты .
Значение
Цианобактерии, по общепринятой версии, явились «творцами» современной кислородсодержащей атмосферы на Земле, что привело к « кислородной катастрофе » — глобальному изменению состава атмосферы Земли, произошедшему в самом начале протерозоя (около 2,4 млрд лет назад), которое привело к последующей перестройке биосферы и глобальному гуронскому оледенению .
В настоящее время, являясь значительной составляющей океанического планктона, цианобактерии стоят в начале большей части пищевых цепей и производят значительную часть кислорода (вклад точно не определён: наиболее вероятные оценки колеблются от 20 % до 40 %).
Однако многие цианобактерии являются токсичными для зоопланктона , поэтому при массовом цветении вызывают гибель многих его представителей, нанося вред питающимся зоопланктоном организмам, о чём свидетельствует выявление мёртвого планктона в пробах с помощью красителя, например, анилинового .
Наиболее многочисленными оксигенными фототрофными организмами в океане являются представители родов Prochlorococcus и .
Цианобактерия стала первым фотосинтезирующим организмом, чей геном был полностью расшифрован.
В настоящее время цианобактерии служат важнейшими модельными объектами исследований в биологии. В Южной Америке и Китае бактерии родов спирулина и носток из-за недостатка других видов продовольствия используют в пищу: их высушивают, а затем готовят муку. Рассматривается возможное применение цианобактерий в создании замкнутых циклов жизнеобеспечения.
Классификация
Исторически существовало несколько систем классификации высших уровней цианобактерий.
В соответствии с « Справочником Берджи по бактериологической систематике », цианобактерии распределены по морфологии на 5 порядков. Хроококковые ( Chroococcales ) и ( ) объединяют одиночные или колониальные сравнительно простые формы, в порядки ( ), ( ), ( ) входят нитчатые формы. Порядок Oscillatoriales включает в себя нитчатые безгетероцистные виды. Нитчатые формы, имеющие гетероцисты, делятся на виды с настоящим ветвлением Stigonematales , неветвящиеся и виды с ложным ветвлением Nostocales . «Высокоорганизованные» порядки содержат нитчатые формы, разница между ними — в наличии или отсутствии истинного ветвления и в наличии или отсутствии дифференцированных клеток (гетероцист и гормогониев). Внесистематической группой цианобактерий считаются « прохлорофиты » — цианобактерии, содержащие помимо хлорофилла a какой-либо другой хлорофилл (b, с или d) . Некоторые из них не имеют фикобилипротеинов (хотя это — один из основных признаков цианобактерий). Родство установлено по гомологии 16S rDNA и генов фотосинтетического аппарата (psbA, psbB).
По данным сайта AlgaeBase , на январь 2018 года отдел делят на следующие таксоны рангом по порядок включительно :
-
Класс
- Роды incertae sedis
-
Подкласс
- Порядок
- Порядок
-
Подкласс
- Порядок —
-
Подкласс
- Порядок Chroococcales — Хроококковые
- Порядок
- Порядок —
- Порядок —
- Порядок
-
Подкласс
- Порядок
- Порядок
По данным сайта NCBI , на январь 2018 года в отдел включают следующие порядки :
- Роды incertae sedis
- Порядок
- Порядок
- Порядок —
- Порядок —
- Порядок
- Порядок
-
Подкласс
- Порядок Chroococcales — Хроококковые
- Порядок —
-
Класс
- Порядок
Ранее выделявшийся порядок Prochlorales понижен в ранге до семейства порядка Synechococcales , а порядок Стигонемовые ( Stigonematales ) синонимизирован с ностоковыми.
См. также
Примечания
- Комментарии
- Время разделения эволюционных линий Gloeomargarita и предка хлоропластов оценивают в 2,1 млрд лет назад, а время существования последнего общего предка архепластид — в 1,9 млрд лет назад .
- То есть в результате симбиоза непосредственно с цианобактерией, а не с уже содержащей пластиду эукариотической клеткой.
- Источники
- ↑ Биологический энциклопедический словарь / Гл. ред. М. С. Гиляров ; Редкол.: А. А. Баев , Г. Г. Винберг , Г. А. Заварзин и др. — М. : Сов. энциклопедия , 1986. — С. 576. — 831 с. — 100 000 экз.
- ↑ Синезелёные водоросли / Ю. Е. Петров // Сафлор — Соан. — М. : Советская энциклопедия, 1976. — ( Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 23).
- Исторически названия таксонов синезелёных водорослей регулируются МКН , поэтому высший ранг группы — отдел, а не тип, как у остальных бактерий.
- ↑ Sánchez-Baracaldo P., Raven J. A., Pisani D., Knoll A. H. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2017. — Vol. 114 , no. 37 . — doi : . — . 23 января 2022 года.
- Анна Семёнова, канд. биол. наук. «Инопланетянин» с болота Целау // Наука и жизнь . — 2022. — № 10 . — С. 3,4 .
- Daniella Mella-Flores, Christophe Six, Morgane Ratin, Frédéric Partensky, Christophe Boutte. Prochlorococcus and Synechococcus have Evolved Different Adaptive Mechanisms to Cope with Light and UV Stress (англ.) // Frontiers in Microbiology. — 2012. — Т. 3 . — ISSN . — doi : .
- : [ англ. ] // AlgaeBase. (Дата обращения: 12 февраля 2018) .
- : [ англ. ] // NCBI . (Дата обращения: 12 февраля 2018) .
Литература
- Гусев М. В. , Никитина А. А. . — М. : Наука , 1979. — 228 с. — 950 экз.
- Судьина Е. Г., , Костлан Н. В. и др. / Отв. ред. К. М. Сытник . — Киев: Наукова думка , 1978. — 264 с. — 1000 экз.
Ссылки
- Gillian Cribbs (1997) Nature’s Superfood, the Blue-Green Algae Revolution . Newleaf. ISBN 0-7522-0569-2
- , (1992, 1994) . Little, Brown. ISBN 0-316-77163-5
- Dyer, Betsey D. A Field Guide to Bacteria. Ithaca: Comstock Publishing Associates, 2003.
- Введение в биологию цианобактерий Беркли . 03 Feb. 2006.
- Gomont M. (1892): Monographie des Oscillatoriées (Nostocacées homocystées). — Ann. Sci. Nat. Bot. Ser. 7, 15: 263—368, 16: 91—264.
|
Галобактерии
( Бактериородопсин ) |
|||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
( Хлорофилл ) |
|||||||||||||||||
|
Пурпурные бактерии
( Бактериохлорофиллы a и b ) |
|
||||||||||||||||
|
Зелёные бактерии
( Бактериохлорофиллы c и d ) |
|
||||||||||||||||
|
Гелиобактерии
( Бактериохлорофилл g ) |
|
||||||||||||||||
|
( Бактериохлорофиллы a ) |
|
||||||||||||||||
| См. также | |||||||||||||||||
- 2020-12-01
- 1