Быстрая коммутация пакетов
- 1 year ago
- 0
- 0


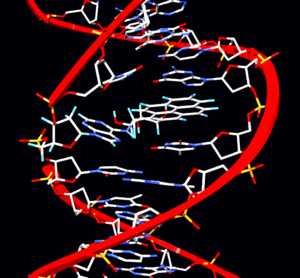
Мута́ция ( лат. «изменение») — стойкое (то есть такое, которое может быть унаследовано потомками данной клетки или организма ) изменение генома . Термин предложен Хуго де Фризом в 1901 году .
Мутагенез — процесс возникновения мутаций.
Мутации делятся на спонтанные и индуцированные .
Спонтанные мутации возникают самопроизвольно на протяжении всей жизни организма в нормальных для него условиях окружающей среды с частотой около — на нуклеотид за клеточную генерацию организма.
Индуцированными мутациями называют наследуемые изменения генома , возникающие в результате тех или иных мутагенных воздействий в искусственных (экспериментальных) условиях или при неблагоприятных воздействиях окружающей среды .
Мутации появляются постоянно в ходе процессов, происходящих в живой клетке. Основные процессы, приводящие к возникновению мутаций — репликация ДНК , нарушения репарации ДНК , транскрипции и генетическая рекомбинация .
Многие спонтанные химические изменения нуклеотидов приводят к мутациям, которые возникают при репликации . Например, из-за дезаминирования цитозина напротив гуанина в цепь ДНК может включаться урацил (образуется пара У-Г вместо канонической пары Ц-Г). При репликации ДНК напротив урацила в новую цепь включается аденин , образуется пара У-А, а при следующей репликации она заменяется на пару Т-А, то есть происходит транзиция (точечная замена пиримидина на другой пиримидин или пурина — на другой пурин).
Из процессов, связанных с рекомбинацией , наиболее часто приводит к мутациям неравный кроссинговер . Он происходит обычно в тех случаях, когда в хромосоме имеется несколько дуплицированных копий исходного гена, сохранивших похожую последовательность нуклеотидов. В результате неравного кроссинговера в одной из рекомбинантных хромосом происходит дупликация , а в другой — делеция .
Спонтанные повреждения ДНК встречаются довольно часто, такие события имеют место в каждой клетке. Для устранения последствий подобных повреждений имеются специальные репарационные механизмы (например, ошибочный участок ДНК вырезается и на этом месте восстанавливается исходный). Мутации возникают лишь тогда, когда репарационный механизм по каким-то причинам не работает или не справляется с устранением повреждений. Мутации, возникающие в генах, кодирующих белки , ответственные за репарацию, могут приводить к многократному повышению (мутаторный эффект) или понижению (антимутаторный эффект) частоты мутирования других генов. Так, мутации генов многих ферментов системы эксцизионной репарации приводят к резкому повышению частоты соматических мутаций у человека, а это, в свою очередь, приводит к развитию пигментной ксеродермы и злокачественных опухолей покровов. Мутации могут появляться не только при репликации, но и при репарации — эксцизионной репарации или при пострепликативной.
В настоящее время существует несколько подходов для объяснения природы и механизмов образования мутаций. Общепринятой, в настоящее время, является полимеразная модель мутагенеза. Она основана на идее о том, что единственной причиной образования мутаций являются случайные ошибки ДНК-полимеразы. В предложенной Уотсоном и Криком таутомерной модели мутагенеза впервые была высказана идея о том, что в основе мутагенеза лежит способность оснований ДНК находиться в различных таутомерных формах. Процесс образования мутаций рассматривается как чисто физико-химическое явление. Полимеразно-таутомерная модель ультрафиолетового мутагенеза опирается на идею о том, что при образовании цис-син циклобутановых пиримидиновых димеров может изменяться таутомерное состояние входящих в них оснований. Изучается склонный к ошибкам и SOS-синтез ДНК, содержащей цис-син циклобутановые пиримидиновые димеры . Существуют и другие модели.
В полимеразной модели мутагенеза считается, что единственной причиной образования мутаций являются спорадические ошибки ДНК-полимераз . Впервые полимеразная модель ультрафиолетового мутагенеза была предложена Бреслером . Он предположил, что мутации появляются в результате того, что ДНК-полимеразы напротив фотодимеров иногда встраивают некомплементарные нуклеотиды. В настоящее время такая точка зрения является общепринятой . Известно А-правило (A-rule), согласно которому напротив поврежденных участков ДНК-полимераза чаще всего встраивает аденины. Полимеразная модель мутагенеза объясняет природу мишенных мутаций замены оснований .
Уотсон и Крик предположили, что в основе спонтанного мутагенеза лежит способность оснований ДНК переходить при некоторых условиях в неканонические таутомерные формы, влияющие на характер спаривания оснований. Эта гипотеза привлекала к себе внимание и активно развивалась. Обнаружены редкие таутомерные формы цитозина в кристаллах оснований нуклеиновых кислот , облученных ультрафиолетовым светом. Результаты многочисленных экспериментальных и теоретических исследований однозначно говорят о том, что основания ДНК могут переходить из канонических таутомерных форм в редкие таутомерные состояния. Было выполнено много работ посвященных исследованиям редких таутомерных форм оснований ДНК. С помощью квантовомеханических расчетов и метода Монте-Карло было показано, что таутомерное равновесие в цитозин — содержащих димерах и в гидрате цитозина сдвинуто по направлению к их имино формам как в газовой фазе, так и в водном растворе. На этой основе объясняется ультрафиолетовый мутагенез . В паре гуанин — цитозин устойчивым будет только одно редкое таутомерное состояние, в котором атомы водородов первых двух водородных связей , отвечающих за спаривание оснований, одновременно изменяют свои положения. А поскольку при этом изменяются положения атомов водорода, участвующих в Уотсон-Криковском спаривании оснований, то следствием может быть образование мутаций замены оснований, транзиций от цитозина к тимину или образование гомологичных трансверсий от цитозина к гуанину. Участие редких таутомерных форм в мутагенезе обсуждалось неоднократно.
В работах Полтева с соавторами предложен и обоснован молекулярный механизм узнавания полимеразами комплементарных пар оснований нуклеиновых кислот. На основании этой модели были изучены некоторые закономерности спонтанного и индуцированного аналогами оснований мутагенеза. Объяснено образование мутаций замены оснований в предположении, что главной причиной мутагенеза является образование неканонических пар оснований, типа Хугстиновских пар. .
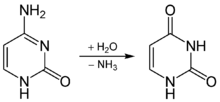
Предполагается, что одной из причин образования мутаций замены основания является дезаминирование 5-метилцитозина , что может вызывать транзиции от цитозина к тимину. Из-за дезаминирования цитозина напротив него в цепь ДНК может включаться урацил (образуется пара У-Г вместо канонической пары Ц-Г). При репликации ДНК напротив урацила в новую цепь включается аденин, образуется пара У-А, а при следующей репликации она заменяется на пару Т-А, то есть происходит транзиция (точечная замена пиримидина на другой пиримидин или пурина на другой пурин).
Существует несколько классификаций мутаций по различным критериям. Мёллер предложил делить мутации по характеру изменения функционирования гена на гипоморфные (измененные аллели действуют в том же направлении, что и аллели дикого типа; синтезируется лишь меньше белкового продукта), аморфные (мутация выглядит, как полная потеря функции гена, например, мутация white у Drosophila ), антиморфные (мутантный признак изменяется, например, окраска зерна кукурузы меняется с пурпурной на бурую) и неоморфные .
В современной учебной литературе используется и более формальная классификация, основанная на характере изменения структуры отдельных генов, хромосом и генома в целом. В рамках этой классификации различают следующие виды мутаций:
Геномные : — полиплоидизация (образование организмов или клеток, геном которых представлен более чем двумя (3n, 4n, 6n и т. д.) наборами хромосом) и анеуплоидия (гетероплоидия) — изменение числа хромосом, не кратное гаплоидному набору (см. Инге-Вечтомов, 1989). В зависимости от происхождения хромосомных наборов среди полиплоидов различают аллополиплоидов, у которых имеются наборы хромосом, полученные при гибридизации от разных видов, и аутополиплоидов, у которых происходит увеличение числа наборов хромосом собственного генома, кратное n.
При хромосомных мутациях происходят крупные перестройки структуры отдельных хромосом. В этом случае наблюдаются потеря ( делеция ) или удвоение части ( дупликация ) генетического материала одной или нескольких хромосом, изменение ориентации сегментов хромосом в отдельных хромосомах ( инверсия ), а также перенос части генетического материала с одной хромосомы на другую ( транслокация ) (крайний случай — объединение целых хромосом, так называемая робертсоновская транслокация , которая является переходным вариантом от хромосомной мутации к геномной).
На генном уровне изменения первичной структуры ДНК генов под действием мутаций менее значительны, чем при хромосомных мутациях, однако генные мутации встречаются более часто. В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. В том случае, когда под действием мутации изменяется лишь один нуклеотид, говорят о точечных мутациях .
Точечная мутация, или единственная замена оснований, — тип мутации в ДНК или РНК, для которой характерна замена одного азотистого основания другим. Термин также применяется и в отношении парных замен нуклеотидов. Термин точечная мутация включает так же инсерции и делеции одного или нескольких нуклеотидов. Выделяют несколько типов точечных мутаций.
Встречаются также сложные мутации. Это такие изменения ДНК, когда один её участок заменяется участком другой длины и другого нуклеотидного состава .
Точечные мутации могут появляться напротив таких повреждений молекулы ДНК, которые способны останавливать синтез ДНК. Например, напротив циклобутановых пиримидиновых димеров. Такие мутации называются мишенными мутациями (от слова «мишень») . Циклобутановые пиримидиновые димеры вызывают как мишенные мутации замены оснований [6 9], так и мишенные мутации сдвига рамки .
Иногда точечные мутации образуются на, так называемых, неповрежденных участках ДНК, часто в небольшой окрестности от фотодимеров. Такие мутации называются немишенными мутациями замены оснований или немишенными мутациями сдвига рамки .
Точечные мутации образуются не всегда сразу же после воздействия мутагена. Иногда они появляются после десятков циклов репликаций. Это явление носит название задерживающихся мутаций . При нестабильности генома, главной причине образования злокачественных опухолей, резко возрастает количество немишенных и задерживающихся мутаций .
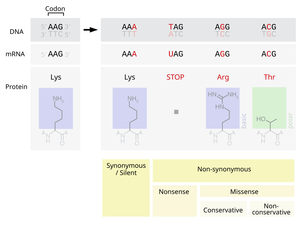
Возможны четыре генетических последствия точковых мутаций: 1) сохранение смысла кодона из-за вырожденности генетического кода (синонимическая замена нуклеотида), 2) изменение смысла кодона, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи (миссенс-мутация), 3) образование бессмысленного кодона с преждевременной терминацией (нонсенс-мутация). В генетическом коде имеются три бессмысленных кодона: амбер — UAG, охр — UAA и опал — UGA (в соответствии с этим получают название и мутации, приводящие к образованию бессмысленных триплетов — например амбер-мутация), 4) обратная замена (стоп-кодона на смысловой кодон).
По влиянию на экспрессию генов мутации разделяют на две категории: мутации типа замен пар оснований и типа сдвига рамки считывания (frameshift) . Последние представляют собой делеции или вставки нуклеотидов, число которых не кратно трём, что связано с триплетностью генетического кода.
Первичную мутацию иногда называют прямой мутацией , а мутацию, восстанавливающую исходную структуру гена, — обратной мутацией , или реверсией. Возврат к исходному фенотипу у мутантного организма вследствие восстановления функции мутантного гена нередко происходит не за счет истинной реверсии, а вследствие мутации в другой части того же самого гена или даже другого неаллельного гена. В этом случае возвратную мутацию называют супрессорной. Генетические механизмы, благодаря которым происходит супрессия мутантного фенотипа, весьма разнообразны.
Почковые мутации (спорты) — стойкие соматические мутации, происходящие в клетках точек роста растений. Приводят к клоновой изменчивости . При вегетативном размножении сохраняются. Многие сорта культурных растений являются почковыми мутантами .
Мутации, которые ухудшают деятельность клетки в многоклеточном организме, часто приводят к уничтожению клетки (в частности, к программируемой смерти клетки, — апоптозу ). Если внутри- и внеклеточные защитные механизмы не распознали мутацию, и клетка прошла деление, то мутантный ген передастся всем потомкам клетки и, чаще всего, приводит к тому, что все эти клетки начинают функционировать иначе.
Мутация в соматической клетке сложного многоклеточного организма может привести к злокачественным или доброкачественным новообразованиям , мутация в половой клетке — к изменению свойств всего организма-потомка.
В стабильных (неизменных или слабо изменяющихся) условиях существования большинство особей имеют близкий к оптимальному генотип, а мутации вызывают нарушение функций организма, снижают его приспособленность и могут привести к смерти особи. Однако в очень редких случаях мутация может привести к появлению у организма новых полезных признаков, и тогда последствия мутации оказываются положительными; в этом случае они являются средством адаптации организма к окружающей среде и, соответственно, называются адаптационными .
При существенном изменении условий существования те мутации, которые раньше были вредными, могут оказаться полезными. Таким образом, мутации являются материалом для естественного отбора . Так, мутанты- меланисты (темноокрашенные особи) в популяциях березовой пяденицы в Англии впервые были обнаружены учеными среди типичных светлых особей в середине XIX века. Темная окраска возникает в результате мутации одного гена. Бабочки проводят день на стволах и ветвях деревьев, обычно покрытых лишайниками , на фоне которых светлая окраска является маскирующей. В результате промышленной революции, сопровождающейся загрязнением атмосферы, лишайники погибли, а светлые стволы берез покрылись копотью. В результате к середине XX века (за 50-100 поколений) в промышленных районах темная морфа почти полностью вытеснила светлую. Было показано, что главная причина преимущественного выживания чёрной формы — хищничество птиц, которые избирательно выедали светлых бабочек в загрязненных районах.
Если мутация затрагивает «молчащие» участки ДНК либо приводит к замене одного элемента генетического кода на синонимичный, то она обычно никак не проявляется в фенотипе (проявление такой синонимичной замены может быть связано с разной частотой употребления кодонов). Однако методами генного анализа такие мутации можно обнаружить. Поскольку чаще всего мутации происходят в результате естественных причин, то в предположении, что основные свойства внешней среды не менялись, получается, что должна быть примерно постоянной. Этот факт можно использовать для исследования филогении — изучения происхождения и родственных связей различных таксонов , в том числе и человека . Таким образом, мутации в молчащих генах служат для исследователей «молекулярными часами». Теория «молекулярных часов» исходит также из того, что большинство мутаций нейтрально, и скорость их накопления в данном гене не зависит или слабо зависит от действия естественного отбора и потому остается постоянной в течение длительного времени. Для разных генов эта скорость, тем не менее, будет различаться.
Исследование мутаций в митохондриальной ДНК (наследуется по материнской линии) и в Y-хромосомах (наследуется по отцовской линии) широко используется в эволюционной биологии для изучения происхождения рас и народностей , реконструкции биологического развития человечества.
В 1940-е годы среди микробиологов была популярна точка зрения, согласно которой мутации вызываются воздействием фактора среды (например, антибиотика ), к которому они позволяют адаптироваться. Для проверки этой гипотезы был разработан флуктуационный тест и метод реплик .
Флуктуационный тест Лурии — Дельбрюка заключается в том, что небольшие порции исходной культуры бактерий рассеивают в пробирки с жидкой средой, а после нескольких циклов делений добавляют в пробирки антибиотик. Затем (без последующих делений) на чашке Петри с твердой средой высевают выживших устойчивых к антибиотику бактерий. Тест показал, что число устойчивых колоний из разных пробирок очень изменчиво — в большинстве случаев оно небольшое (или нулевое), а в некоторых случаях очень высокое. Это означает, что мутации, вызвавшие устойчивость к антибиотику, возникали в случайные моменты времени как до, так и после его воздействия.
Метод реплик заключается в том, что с исходной чашки Петри, где на твердой среде растут колонии бактерий, делается отпечаток на ворсистую ткань, а затем с ткани бактерии переносятся на несколько других чашек, где рисунок их расположения оказывается тем же, что на исходной чашке. После воздействия антибиотиком на всех чашках выживают колонии, расположенные в одних и тех же точках. Высевая такие колонии на новые чашки, можно показать, что все бактерии внутри колонии обладают устойчивостью.
Таким образом, обоими методами было доказано, что «адаптивные» мутации возникают независимо от воздействия того фактора, к которому они позволяют приспособиться, и в этом смысле мутации случайны. Однако несомненно, что возможность тех или иных мутаций зависит от генотипа и канализована предшествующим ходом эволюции (см. Закон гомологических рядов в наследственной изменчивости ).
Кроме того, закономерно различается частота мутирования разных генов и разных участков внутри одного гена. Также известно, что высшие организмы используют «целенаправленные» (то есть происходящие в определённых участках ДНК) мутации в механизмах иммунитета [ источник не указан 3474 дня ] . С их помощью создаётся разнообразие клонов лимфоцитов , среди которых в результате всегда находятся клетки, способные дать иммунный ответ на новую, неизвестную для организма болезнь. Подходящие лимфоциты подвергаются положительной селекции , в результате возникает иммунологическая память . (В работах Юрия Чайковского говорится и о других видах направленных мутаций.)
| Эволюционные процессы |

|
|
|---|---|---|
| Факторы эволюции | ||
| Генетика популяций | ||
| Происхождение жизни | ||
| Исторические концепции | ||
| Современные теории | ||
| Эволюция таксонов | ||