C-реактивный белок
- 1 year ago
- 0
- 0

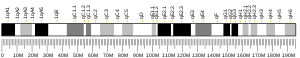
Бромодоменсодержащий белок 4 ( англ. Bromodomain-containing protein 4 , BRD4) — хроматинсвязывающий белок , который у человека кодируется геном BRD4 . Он узнаёт и связывается с остатками ацетилированного лизина в составе гистонов и других белков. BRD4 принимает участие в регуляции транскрипции и передаче эпигенетической информации при делении клетки . В 2016 году было установлено, что BRD4 обладает собственной гистонацетилтрансферазной активностью .
BRD4 принадлежит к семейству белков BET , содержащих два тандемных бромодомена и экстратерминальный домен . Кроме BRD4, к этому семейству относятся ещё три белка человека: BRD2, BRD3 и BRDT .
РНК BRD4 и белок присутствуют в подавляющем большинстве типов клеток и тканей .
BRD4 локализован главным образом в ядре клетки , но может присутствовать и в цитоплазме в небольшом количестве .
В результате альтернативного сплайсинга формируются две изоформы белка. Более длинная и более распространённая изоформа состоит из 1362 аминокислотных остатков и имеет молекулярную массу около 152 кДа . Биологическую активность BRD4 связывают именно с этой изоформой. Вторая изоформа укорочена с C-конца, состоит из 723 аминокислотных остатков и имеет молекулярную массу 88 кДа. Короткую изоформу удаётся обнаружить гораздо реже, чем длинную, и её функции неизвестны .
BRD4 содержит два консервативных бромодомена на N-конце молекулы. Бромодомены состоят из четырёх α-спиралей , соединённых подвижными петлями. Вместе спирали и петли формируют гидрофобный карман, который распознаёт остатки ацетиллизина в белках .
C-концевой экстратерминальный домен BRD4 принимает участие в связывании фактора транскрипции P-TEFb .
BRD4 выступает в качестве глобального регулятора транскрипции, при этом он может действовать несколькими способами, пассивно и активно.
Транскрипция существенной доли генов многоклеточных организмов останавливается на стадии ранней элонгации , когда длина синтезированной пре-мРНК составляет около 60 нуклеотидов . Одной из причин такой остановки является последовательное присоединение к транскрипционному комплексу негативных регуляторных факторов DSIF ( англ. DRB sensitivity inducing factor ) и NELF ( англ. negative elongation factor ), которые блокируют дальнейшее продвижение РНК-полимеразы II . Повторный запуск транскрипции осуществляется при участии фактора P-TEFb. P-TEFb представляет собой гетеродимер, состоящий из циклинзависимой киназы 9 и её регуляторной субъединицы — циклина T1, T2 или K. P-TEFb фосфорилирует DSIF и NELF и тем самым подавляет их активность. Кроме того, P-TEFb фосфорилирует гептадные повторы в С-концевом домене РНК-полимеразы II по положению Ser -2, что также способствует активной элонгации . За привлечение P-TEFb в транскрипционный комплекс отвечает BRD4, который связывается с околопромоторными областями генов за счёт взаимодействия с остатками ацетиллизина в гистонах .
BRD4 является атипичной протеинкиназой : он способен самостоятельно фосфорилировать РНК-полимеразу II по положению Ser-2 при отсутствии явной структурной гомологии с другими протеинкиназами .
BRD4 стимулирует транскрипцию гена CD274 , кодирующего белок PD-L1, независимо от MYC. На примере рака яичника и B-клеточной лимфомы было продемонстрировано, что применение ингибиторов BRD4, таких как JQ1 , приводит к снижению количества PD-L1 на поверхности опухолевых клеток и вызывает T-клеточный иммунный ответ на них .
В 2016 году было установлено, что BRD4 является гистоацетилтрансферазой . Он ацетилирует остатки лизина в N-концевых фрагментах гистонов 3 и 4, а также остаток Lys-122 в гистоне 3. Ацетилирование последнего остатка оказывает более существенное влияние на дестабилизацию нуклеосомы , потому что он находится в глобулярном C-концевом домене гистона и в непосредственном контакте с ДНК . Известны ещё только две гистоацетилтрансферазы, способные ацетилировать Lys-122 в гистоне 3: p300 и CBP. Гистонацетилтрансферазная активность BRD4 вызывает декомпактизацию хроматина (а следовательно, стимулирует транскрипцию) во многих участках генома , в том числе в районе генов MYC , FOS и AURKB .
Впервые особая роль BRD4 в заболеваниях человека была описана при анализе редкого типа карциномы , характеризующегося хромосомной транслокацией t(15;19) . При данной транслокации происходит слияние первой половины гена BRD4 и практический всей кодирующей области гена NUT . В результате формируется химерный онкобелок BRD4-NUT, который отвечает за необычно агрессивный характер данной карциномы .