Эпителий
- 1 year ago
- 0
- 0

.jpg)
Эпите́лий ( лат. epithelium , от греч. ἐπι- — «сверх-» и θηλή — «сосок молочной железы»), или эпителиа́льная ткань — совокупность дифференцированных клеток , тесно прилегающих друг к другу в виде пласта, лежащего на базальной мембране . Эпителий лежит на границе внешней или внутренней среды организма и образует большую часть желёз . В эпителии отсутствуют межклеточное вещество и кровеносные сосуды , однако имеется обильная иннервация . Эпителии могут происходить из всех трёх зародышевых листков : энтодермы , эктодермы и мезодермы , хотя большая часть эпителиев происходит из энтодермы и эктодермы. Существует несколько классификаций эпителиальных тканей, однако наиболее распространена морфологическая классификация, в которой эпителии разделяют по числу слоёв (одно- или многослойные) и форме клеток (плоская, кубическая, призматическая), а также способности к ороговению . Эпителиальным тканям присуща высокая регенеративная способность.
По морфофизиологическим свойствам клетки эпителиев весьма разнообразны, однако есть ряд черт, присущих всем клеткам эпителиев ( эпителиоцитам ). В эпителиях практически отсутствует межклеточное вещество , клетки плотно прилегают друг к другу и формируют между собой разнообразные межклеточные контакты — плотные контакты , щелевые контакты и десмосомы . Благодаря плотным контактам обеспечивается избирательная проницаемость эпителия . Эпителий располагается на базальной мембране толщиной около 1 мкм , компоненты которой образуют и сами клетки эпителия, и нижележащая соединительная ткань .
Форма эпителиальных клеток может быть кубической, цилиндрической, плоской и зависит от количества цитоплазмы и органелл в ней, что связано с уровнем метаболической активности клеток. Наименее активны плоские эпителиоциты. В столбчатых клетках высокоактивен эндоплазматический ретикулум , имеются многочисленные митохондрии . Иногда эпителиальные клетки содержат пигмент , например, к числу таких клеток относятся клетки радужной оболочки , сетчатки , кожи .
Базальная мембрана подразделяется на подэпителиальную электронно -прозрачную светлую пластинку толщиной от 20 до 40 нм ( ) и тёмную пластинку ( ) толщиной 20—60 нм (по другим данным, 20—100 нм ). Светлая пластинка может располагаться по обе стороны от тёмной пластинки или только по одну и сформирована богатым кальцием и бедным белками аморфным веществом. В состав тёмной пластинки входит аморфный , механическую прочность которого обеспечивает фибриллярный — белок базальной мембраны. Аморфное вещество базальной мембраны богато гликопротеинами , протеогликанами (например, ) и углеводами (в частности, гликозаминогликанами ). Эпителиальные клетки связаны с базальной мембраной с помощью полудесмосом при участии гликопротеинов фибронектина , ламинина и нидогена , которые образуют адгезивный субстрат. Кроме того, актиновые микрофиламенты цитоскелета эпителиоцитов связаны с базальной мембраной посредством фокальных контактов . Важную роль в прикреплении к базальной мембране играют ионы кальция. Базальная мембрана, помимо того, что служит механической опорой для эпителиального пласта, обладает трофической функцией — пропускает питательные вещества к клеткам эпителия, поскольку в эпителиях отсутствуют кровеносные сосуды. Также базальная мембрана выступает в роли барьера, через который осуществляется избирательный транспорт веществ, ограничивает возможность инвазивного роста эпителия и задействована в регенерации эпителия . Базальная мембрана связана с подлежащей соединительной тканью с помощью якорных фибрилл, сформированных . В тех случаях когда слои эпителия не разделены соединительной тканью (например, в альвеолах лёгкого ), базальные мембраны каждого слоя сливаются с образованием общей утолщённой базальной мембраны. Иногда с базальной мембраной тесно связана ретикулярная пластинка (lamina retucularis), образованная ретикулярными волокнами . В некоторых источниках описанную выше структуру называют базальной пластинкой, а термин «базальная мембрана» применяют по отношению к слою, образующемуся при слиянии двух базальных пластинок или базальной и ретикулярной пластинок и положительно окрашивающемуся при .

Эпителиоциты обладают выраженной полярностью: их апикальные и базальные части резко отличаются друг от друга, в связи с чем в них выделяют апикальный и базолатеральный домены, которые различаются и функционально. Наиболее ярко полярность клеток выражена в однослойных эпителиях, например, апикальные поверхности эпителиоцитов кишечника несут многочисленные микроворсинки , а в базальной части микроворсинок нет и происходит всасывание питательных веществ и выделение продуктов обмена в кровь или лимфу . Микроворсинки представляют собой пальцевидные выросты длиной около 1 мкм и шириной 0,08 мкм. Комплекс микроворсинок и покрывающего их гликокаликса можно увидеть в световой микроскоп ; он известен как . Очень длинные микроворсинки, лишённые подвижности, называют . Стереоцилии можно найти в придатке яичка (эпидидимисе) в мужской половой системе и у волосковых клеток внутреннего уха . Иногда апикальные домены эпителиоцитов несут реснички . Апикальный домен эпителиоцита несёт множество ионных каналов , белков-переносчиков , молекулы мембранной АТФазы , гликопротеины, гидролитические ферменты , а также аквапорины . Базальный домен может содержать инвагинации клеточной мембраны и образует полудесмосомы, закрепляющие клетку на субстрате. Впячивания базальной мембраны увеличивают площадь поверхности , участвующей в транспорте веществ . Впячивания могут образовываться и на боковых ( латеральных ) поверхностях эпителиальных клеток . В многослойных эпителиях проявляется полярность отдельных слоёв: эпителиоциты базального слоя и поверхностных слоёв резко отличаются друг от друга .
Эпителии обладают высокой способностью к регенерации благодаря митотическим делениям и дифференцировке стволовых клеток , которые сохраняют способность к делению в течение всей жизни организма .
В эпителии имеется обильная иннервация, и расположено множество чувствительных нервных окончаний ( рецепторов ) .
В качестве цитогенетического маркера эпителиальных клеток используется белок цитокератин , входящий в состав промежуточных филаментов . Известно более 20 форм цитокератина, которые присутствуют в разных видов эпителия. Благодаря гистохимическому обнаружению форм цитокератина в исследуемой ткани можно установить, что её происхождение связано с эпителием, что имеет огромное значение в гистологическом исследовании опухолей .
Эпителиальные ткани выполняют множество важных функций в организме. Они защищают внутреннюю среду организма, обеспечивают веществ, секретируют слизь , гормоны , ферменты и другие вещества, осуществляют всасывание веществ из просвета органа (в кишечнике или почечных канальцах ). Благодаря селективной проницаемости контактов между клетками эпителии контролируют транспорт веществ между различными органами . Некоторые производные эпителия, такие как вкусовые луковицы , сетчатка и специализированные волосковые клетки уха осуществляют сенсорную функцию .
Существует несколько классификаций эпителиев, которые учитывают различные особенности эпителиальных тканей: морфологическая, функциональная, онтофилогенетическая .

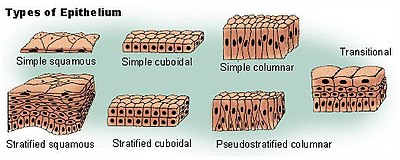
Наибольшее распространение получила морфологическая классификация, основанная на форме клеток и их связи с базальной мембраной. Согласно этой классификации, все эпителии подразделяются на однослойные, в которых все клетки связаны с базальной мембраной, и многослойные, в которых с базальной мембраной непосредственно взаимодействует лишь самый нижний слой клеток. По форме клеток однослойный эпителий подразделяют на плоский (сквамозный), кубический и призматический (столбчатый, цилиндрический ), в случае многослойных эпителиев при классификации учитывается форма только клеток наружного слоя .
.jpg)
Однослойные эпителии подразделяют на однорядные, в которых ядра эпителиоцитов лежат на одном уровне, и многорядные, или , у которых ядра клеток лежат на разных уровнях, хотя все клетки сохраняют связь с базальной мембраной .
Клетки однорядного эпителия имеют одинаковую форму (плоскую, кубическую, столбчатую), ядра занимают центральное положение . Клетки плоского эпителия сильно уплощены, цитоплазма формирует тонкую прослойку, и ядра образуют выпуклости на поверхности клетки . Однослойным плоским эпителием является мезотелий , который покрывает серозные оболочки (листки плевры , околосердечную сумку ( перикард ), висцеральную и париетальную брюшину ), а также, по мнению ряда исследователей, эндотелий , выстилающий кровеносные и лимфатические сосуды изнутри (некоторые учёные относят эндотелий к соединительным тканям). Однослойный эпителий покрывает тело некоторых животных, например, ланцетника или полухордовых . Клетки мезотелия несут микроворсинки, плоские, имеют многоугольную форму и неровные края. Мезотелий участвует в выделении и всасывании . Однослойный плоский эпителий, выстилающий околосердечную сумку, называют эндокардом . Эндотелий представляет собой пласт плоских клеток (эндотелиоцитов), которые относительно бедны органеллами, но содержат в цитоплазме пиноцитозные везикулы . Эндотелий задействован в обмене веществ и газов между кровью, лимфой и другими тканями . Кроме того, однослойный плоский эпителий выстилает альвеолы лёгких, петлю Генле и формирует париетальную выстилку в почечных клубочках .
Клетки однослойного кубического эпителия имеют кубическую форму, содержат центрально расположенное округлое ядро. Однослойный кубический эпителий выстилает проксимальные и дистальные почечные канальцы. Клетки проксимальных канальцев несут щёточную каёмку, которая состоит из множества микроворсинок и покрывающего их гликокаликса. В базальных отделах клеток имеются глубокие складки клеточной мембраны, между которыми находятся митохондрии, что придаёт базальной части эпителия проксимальных канальцев поперечную исчерченность. Эпителиальная выстилка почечных канальцев задействована в реабсорбции ряда веществ из первичной мочи . Однослойный кубический эпителий также выстилает многих желёз и покрывает яичники . Однослойный кубический эпителий, покрывающий яичники, называют . Типичный кубический эпителий можно обнаружить в щитовидной железы , на внутренней поверхности хрусталика и пигментном слое сетчатки . Однослойный кубический эпителий покрывает тело ланцетника .
Клетки однослойного призматического эпителия имеют удлинённую форму, их яйцевидные, нередко также удлинённые ядра, как правило, располагаются на одном уровне в базальных частях клеток . Однослойный призматический эпителий характерен для многих отделов пищеварительного тракта — желудка , тонкого и толстого кишечника , жёлчного пузыря , некоторых протоков печени , поджелудочной железы и некоторых других желёз. Апикальные поверхности клеток несут микроворсинки. Многочисленные контакты между эпителиоцитами, выстилающими пищеварительный тракт, не дают выйти в полость тела содержимому соответствующих органов . Клетки однослойного призматического эпителия, который выстилает матку , яйцеводы и мелкие бронхи , несут реснички . Ресничный призматический эпителий выстилает изнутри спинномозговую полость и желудочки головного мозга .

Многорядный (псевдомногослойный, ложномногослойный) эпителий характерен для воздухоносных путей — носовой полости , трахеи , бронхов и некоторых других органов. Многорядные эпителии нередки и среди беспозвоночных ; например, у двустворчатых моллюсков псевдомногослойный эпителий выстилает пищеварительную трубку , а у голожаберных моллюсков он покрывает большую часть тела . Хотя все клетки многорядного эпителия контактируют с базальной мембраной, лишь немногие выходят на поверхность. Клетки, не контактирующие с поверхностью, как правило, имеют широкое основания и сужаются в апикальной части, ядра располагаются в расширенных частях клеток и потому не лежат на одном уровне . В воздухоносных путях ряд клеток многорядного эпителия несёт реснички, а остальные клетки подразделяют на вставочные, базальные и бокаловидные клетки , продуцирующие слизь. Базальные клетки участвуют в регенерации эпителия, так как дифференцируются в ресничные и бокаловидные клетки. Ресничные клетки высокие, призматические, с помощью сгибательных движений ресничек очищают вдыхаемый воздух от частиц пыли . У человека многорядный эпителий, не несущий ресничек, можно обнаружить в мужской уретре , придатке яичка и крупных выводных протоках желёз .

В многослойных эпителиях с базальной мембраной связаны только клетки самого нижнего (базального) слоя. В многослойном плоском неороговевающем эпителии различают три слоя: базальный, шиповатый (промежуточный) и плоский (поверхностный). Клетки базального слоя кубические или призматические, среди них имеются стволовые клетки, сохраняющие способность к митозу; именно благодаря делениям этих клеток популяция эпителиоцитов восстанавливается, несмотря на постоянное отмирание поверхностных клеток эпителия. Шиповатый слой образован эпителиоцитами неправильной многоугольной формы. Клетки базального и шиповатого слоёв содержат хорошо выраженные пучки тонофиламентов кератина . Плоские клетки, образующие поверхностный слой эпителия, постоянно отмирают и отпадают с поверхности эпителия, однако, в отличие от роговых чешуек многослойного плоского ороговевающего эпителия, в них различимы ядра. Многослойный плоский неороговевающий эпителий покрывает роговицу глаза , выстилает ротовую полость и пищевод , а также влагалище и покрывает настоящие голосовые связки . Многослойный неороговевающий эпителий, содержащий множество одноклеточных слизистых желез, покрывает тело у круглоротых и хрящевых рыб .
Поверхность кожи покрыта многослойным плоским ороговевающим эпителием, который формирует эпидермис . В эпидермисе протекает ороговение (кератинизация), в ходе которого эпителиальные клетки кератиноциты дифференцируются в роговые чешуйки. В ходе кератинизации в цитоплазме кератиноцитов синтезируются и накапливаются специфические белки — кислые и щелочные кератины, филаггрин , кератолинин и другие. Большая часть клеток эпидермиса представлена кератиноцитами, которые по мере дифференцировки перемещаются из базального слоя в верхние слои эпидермиса. Помимо кератиноцитов, эпидермис содержит меланоциты , специализированные эпидермальные макрофаги ( клетки Лангерганса ), лимфоциты и клетки Меркеля ( осязательные клетки). Многослойный плоский ороговевающий эпителий подразделяется на несколько слоёв: базальный, шиповатый, зернистый, блестящий и роговой. Последние три слоя наиболее выражены в толстой коже, например, коже, покрывающей подошвы ног. Базальный слой представлен призматическими кератиноцитами, в цитоплазме которых образуется кератин, формирующий тонофиламенты. В базальном слое также находятся стволовые клетки, поэтому иногда базальный слой называют ростовым, или зачатковым. Шиповатый слой сложен кератиноцитами многоугольной формы, которые прочно связаны друг с другом с помощью десмосом. При фиксации клетки шиповатого слоя сморщиваются и отходят друг от друга, и десмосомы становятся хорошо заметны в виде шипиков (отсюда название слоя). Шиповатые кератиноциты содержат , состоящие из кератиновых тонофиламентов, и кератиносомы, или ламеллярные гранулы , содержащие липиды . Содержимое кератиносом посредством экзоцитоза выделяется наружу и формирует богатое липидами цементирующее вещество. Зернистый слой слагают кератиноциты уплощённой формы, содержащие базофильные гранулы. В зернистом слое начинается распад органелл и ядер клеток под действием гидролитических ферментов, связанный с их отмиранием. В зернистом слое синтезируются специфические белки — филаггрин и кератолинин, укрепляющий мембраны клеток. Блестящий слой имеется только в участках с выраженным ороговением эпидермиса (на подошвах и ладонях ). Его образуют плоские мёртвые кератиноциты, лишённые ядер и органелл. Под клеточной мембраной залегает слой укрепляющего белка кератолинина, а в цитоплазме кератогиалиновые гранулы сливаются, образуя единую светопреломляющую массу из кератиновых фибрилл, которые скрепляет аморфный матрикс, содержащий филлагрин. Роговой слой образуют отмершие плоские многоугольные кератиноциты, превратившиеся в роговые чешуйки. Они имеют плотную оболочку из кератолинина и полностью заполнены кератиновыми фибриллами, склеенными аморфным матриксом, не имеют ядер и цитоплазмы . Чешуйки склеивает цементирующее вещество, образованное содержимым кератиносом и обогащённое липидами, что придаёт ему водоотталкивающие свойства. Роговой слой также устойчив к механическим и химическим воздействиям, непроницаем для воды и многих водорастворимых веществ (в том числе ядовитых), имеет низкую теплопроводность . Самые верхние кератиноциты утрачивают связь с эпидермисом и покидают его. Состав кератиноцитов эпидермиса полностью обновляется каждые три—четыре недели . У некоторых амфибий, рептилий и птиц отмершие роговые чешуйки связаны друг с другом и отделяются в виде крупных фрагментов рогового слоя .
Очень необычное строение имеет эпидермис оболочников . У этих животных эпидермис несёт плотный, гибкий и прочный наружный слой, — тунику, состоящую из полисахарида туницина, близкого к целлюлозе растений . Туницин выделяют клетки эпидермиса, и они могут погружаться в него небольшими группами и поодиночке, при этом сохраняя связь с основным слоем эпидермиса при помощи отростков и специальных кровеносных сосудов. Благодаря тунике тело оболочника может прочно прикрепляться к субстрату . У костистых рыб эпидермис содержит множество одноклеточных желёз, а под ним, в дерме, залегают костные чешуи . У амфибий кожа лишена роговых образований, однако эпидермис покрыт одним слоем ороговевших клеток, из-за чего становится невозможной работа одноклеточных костных желёз, поэтому земноводные имеют только сложные многоклеточные железы . У рептилий роговой слой эпидермиса тонкий, твёрдый, гладкий, неэластичный; его клетки слущиваются и обновляются за счёт деления клеток нижележащих слоёв. Периодически в ростковом слое эпидермиса рептилий образуются особые клетки, которые подвергаются неполному ороговению и разрушаются, из-за чего тонкий поверхностный слой отслаивается и начинается линька .
Выделяют также многослойный плоский паракератинизированный эпителий, который близок к многослойному плоскому ороговевающему эпителию, однако самые верхние его клетки сохраняют пикнотические ядра. Таким образом, степень кератинизации такого эпителия занимает промежуточное положение между ороговевающим и неороговевающим эпителиями. Паракератинизированный эпителий покрывает твёрдое нёбо и дёсны .

Многослойный кубический эпителий состоит всего из двух слоёв кубических эпителиоцитов и выстилает протоки потовых желёз . Многослойный столбчатый эпителий в организме человека представлен мало, он встречается только в конъюктиве глаза, протоках слюнных желёз и некоторых участках уретры. В случае многослойного столбчатого эпителия с базальной мембраной контактируют кубические или многоугольные с небольшим числом граней клетки, а верхний слой сложен призматическими эпителиоцитами .

Мочеотводящие органы ( , мочеточники , мочевой пузырь , уретра) выстланы особым , который даёт возможность этим органам сильно растягиваться при заполнении мочой . Своё название он получил в связи с тем, что ранее считалось, что он является переходной формой между многослойным столбчатым и многослойным плоским эпителиями. Поскольку переходный эпителий встречается только в мочеотводящих путях, его также называют уротелием . Переходный эпителий обычно достигает 4—6 клеток в толщину . В переходном эпителии выделяют три слоя: базальный, промежуточный, поверхностный. Базальный слой образуют невысокие цилиндрические или кубические клетки . Промежуточный слой образован клетками многоугольной формы. Клетки поверхностного слоя очень крупные, нередко содержат два или три ядра, и в зависимости от степени растяжения стенки органа и его заполненности мочой имеют куполообразную или уплощённую форму. Когда стенка органа сокращается, некоторые клетки промежуточного слоя выходят на границу с поверхностным слоем и принимают грушевидную форму, а расположенные над ними поверхностные клетки становятся куполообразными. Когда орган заполнен мочой, но его стенка не сокращается, эпителий становится более тонким, поверхностные клетки принимают уплощённую форму .
Эпителий стрекающих также имеет ряд особенностей, отличающих его от эпителиев высших животных. Эпителий стрекающих архаичен, поскольку его клетки слабо дифференцированы: например, у гидры наружный эпителий может возникать за счёт дедифференцировки железистых клеток гастральной полости и их последующей дифференцировки в покровные клетки. У стрекающих также широко распространены эпителиально-мышечные клетки: их части, несущие ядра, образуют пласт на поверхности тела, а вытянутые базальные части содержат сократимые филаменты .
У паразитических плоских червей из групп цестод и трематод тело покрыто специализированным псевдокутикулярным погружённым эпителием. Особенно хорошо псевдокутикулярный погружённый эпителий выражен у цестод, которые лишены кишечника, и их покровный эпителий выполняет одновременно и барьерную, и всасывающую функции, а также защищает червя от действия гидролитических ферментов хозяина. Апикальные части эпителиальных клеток сливаются своими боковыми частями, формируя структурный и функциональный синцитий — тегумент , или псевдокутикулярная пластинка. Тегумент отделён от паренхимы , заполняющей тело червя, базальной мембраной. Базальная мембрана прерывается только в тех местах, где апикальные части эпителиальных клеток соединяются с базальными частями, несущими ядро и залегающими в паренхиме. Однослойный эпителий синцитиального строения характерен для некоторых других беспозвоночных: некоторых турбеллярий , коловраток , скребней и большинства нематод .
У современных многоклеточных животных наиболее распространённым вариантом кожных эпителиев являются кутикулярные эпителии. Они хорошо выражены у оболочников, аннелид , приапулид , моллюсков, членистоногих , нематод и других беспозвоночных. Кутикулярный эпителий состоит из однослойного эпителия и выделяемого им внеклеточного вещества, формирующего кутикулярную пластинку. Кутикулярная пластинка состоит из фибриллярных структур и цементирующего их аморфного матрикса. Фибриллярные структуры могут иметь полисахаридно - белковую природу (артроподный тип кутикулярного эпителия) или состоять из коллагена (аннелидный тип). У нематод имеется многослойная гетерогенная по составу кутикула (нематодный тип) .
По функциям выделяют два ключевых вида эпителиев: поверхностные, или покровные, и железистые. Однако это разделение является условным, так как существуют покровные эпителии, клетки которых способны к секреции (например, покровный эпителий желудка или эпителий, покрывающий тело полухордовых, в котором имеются и ресничные, и железистые клетки ), а в некоторых эпителиях, например, в выстилке тонкой кишки и трахеи, покровные клетки чередуются с бокаловидными клетками, секретирующими слизь . Поверхностные эпителии покрывают поверхности тела, формируют слизистые оболочки внутренних органов (желудка, кишечника, мочевого пузыря, матки и других) и выстилают вторичные полости тела . Эпителии, выстилающие полости внутренних органов, подстилаются слоем соединительной ткани, известным как (lamina propria), которая выполняет поддерживающую и трофическую функцию. Площадь контакта между эпителием и собственной пластинкой увеличивается за счёт особых структур — сосочков, которые представляют собой впячивание соединительной ткани в эпителий . Главная функция поверхностных эпителиев — отделение организма и его органов от внешней среды и осуществление обмена веществ между ними. Кроме того, поверхностные эпителии выполняют защитную функцию. Железистый эпителий образует многие железы организма и осуществляет секреторную функцию, выделяя пищеварительные ферменты , гормоны в эндокринных железах .

Железистые эпителии имеют выраженную секреторную функцию. Клетки, образующие железистый эпителий, называют грандулоцитами. Они синтезируют и выделяют специфические вещества ( секреты ) на поверхность кожи, слизистых оболочек , в полости некоторых внутренних органов (экзокринная секреция) или в кровь и лимфу (эндокринная секреция). Эпителии образуют в организме разнообразные железы, хотя некоторые железы имеют одноклеточное строение — например, бокаловидные клетки, выделяющие слизь . Чаще всего бокаловидные клетки находятся в слизистых (например, слизистой кишечника), но у некоторых животных, например, немертин и моллюсков, они многочисленны и в кожных покровах . Большая часть кожных желёз имеет одноклеточное строение у первичноводных позвоночных , таких как костистые и хрящевые рыбы . У земноводных все кожные железы многоклеточные, причем некоторые из них являются ядовитыми и выделяют токсины белковой природы . Грандулоциты отличаются наличием в цитоплазме секреторных включений, развитым эндоплазматическим ретикулумом (ЭПР) и аппаратом Гольджи (АГ), полярностью в расположении органелл и секреторных гранул. По химическим свойствам секретов экзокринные железы подразделяют на белковые (серозные), слизистые и смешанные . Грандулоциты подстилаются базальной мембраной, а их форма может быть разнообразной и зависит от фазы секреторного цикла. Ядра, как правило, крупные, могут иметь неправильную форму. Клетки, выделяющие белковый секрет, имеют развитый шероховатый ЭПР, а грандулоциты, выделяющие липидный секрет, имеют выраженный гладкий ЭПР. Форма и расположение АГ в клетке зависит от стадии секреторного цикла. Митохондрии многочисленны и, как правило, накапливаются в тех местах клетки, где образуется секрет. Размер и строение секреторных гранул зависят от химического состава секрета. Иногда грандулоциты имеют внутриклеточные секреторные канальцы, которые представляют собой глубокие впячивания клеточной мембраны, покрытые микроворсинками (например, у клеток, образующих в желудке). На базальных поверхностях клеток мембрана образует небольшие складки, вдающиеся в цитоплазму, которые наиболее хорошо выражены у грандулоцитов, секрет которых богат солями . Апикальная поверхность грандулоцитов покрыта микроворсинками. В целом, грандулоциты имеют выраженную полярность, которая обусловлена направленностью секреторных процессов. Периодические изменения железистой клетки, связанные с секреторными процессами, называют секреторным циклом. Вещества, необходимые для образования секрета, грандулоциты получают из крови и лимфы со стороны базальной поверхности. Образование секрета происходит в ЭПР, откуда он перемещается в АГ, где после дальнейших модификаций в виде оформляется в виде гранул. Выделяют три основных механизма выделения секрета грандулоцитами :
Существуют клетки эпителиального происхождения, специализированные на сенсорных функциях — . К их числу относят клетки вкусовых луковиц и клетки . также имеют эпителиальное происхождение, несут отростки и содержат миозин и актиновые филаменты. Они обеспечивают сдавливание протоков ряда желёз (молочных, потовых, слюнных) для выделения секрета .
Онтофилогенетическая классификация, созданная российским гистологом Николаем Григорьевичем Хлопиным , учитывает происхождение эпителиев из разных тканевых зачатков. Согласно онтофилогенетической классификации, выделяются следующие типы эпителиев :

Для млекопитающих весьма характерны роговые образования эпидермального происхождения: волосы, когти, ногти, копыта, рога и чешуйки .
Волосы — образование, присущее исключительно млекопитающим и, по-видимому, наиболее продвинутым терапсидам и не гомологичное каким-либо производным кожи других амниот. Их главная функция — теплоизоляция, хотя толчком для появления волосяного покрова послужила, скорее всего, осязательная функция . Помимо термоизолирующей и осязательной функций, волосы защищают кожу от повреждений и паразитов , улучшают аэро- и гидродинамические свойства тела, обеспечивают видоспецифичность окраски. Отсутствие волос у ряда млекопитающих ( слоны , сирены , часть носорогов , гиппопотамы , китообразные ) носит вторичный характер .
У большинства млекопитающих концевые фаланги пальцев защищены когтями , которые остры и сильно изогнуты у древесных форм и хищников, уплощены и расширены у роющих форм . У многих приматов когти преобразовались в плоские ногти , прикрывающие фалангу лишь сверху; у копытных в результате усложнения когтей сформировались копыта — толстые образования, которые играют роль своеобразного футляра для концевой фаланги, особо важный при беге и прыжках по твёрдому грунту . У многих млекопитающих на хвосте ( бобр , мышиные , шипохвостые , выхухоль , многие сумчатые ) или конечностях (роющие формы) имеются роговые чешуйки . У панголинов и броненосцев крупные роговые чешуи покрывают всё тело, причём у броненосцев они подстилаются ещё костными щитками, являющимися производными дермы и образующими панцирь . Чешуя млекопитающих вполне гомологична таковой у пресмыкающихся . Для некоторых млекопитающих ( носороги , вилороговые , полорогие ) характерно наличие рога — массивного разрастания ороговевшего эпителия, используемого для защиты и нападения .
К числу производных эпителия относятся перья птиц и : само перо или чешуя является производным эпидермиса, а питающий его сосочек принадлежит дерме. Перья производят особые фолликулы, расположенные в эпидермисе и производящие кератины. В отличие от волос, когтей и рогов млекопитающих, состоящих из , перья птиц и чешуи рептилий состоят из .
Первые клеточные слои, близкие к эпителиям, появились в ходе эволюции у губок , а именно, , покрывающая тело губки, и , выстилающая водоносные полости. Однако пинакоциты и хоаноциты не образуют характерных для настоящих эпителиев поясковых соединений , поэтому, вероятно, не функционируют в качестве разграничителей компартментов тела (хотя между клетками губок могут формироваться временные септированные контакты ). Пинакодерму и хоанодерму губок не считают истинными эпителиями ещё и потому, что в большинстве случаев они не подстилаются базальной мембраной. Тем не менее, губки имеют ген , кодирующий коллаген IV типа — маркер базальной мембраны. Кроме того, пинакоциты и хоаноциты губок обладают полярностью в апикально-базальном направлении, которая присуща эпителиоцитам Eumetazoa .
В отличие от губок, стрекающие , по сути, состоят преимущественно из эпителиев: их эпидермис, покрывающий тело снаружи, и , выстилающий гастральную полость, удовлетворяют всем критериям эпителиальных тканей, и единственные мезенхимоподобные клетки стрекающих располагаются в межклеточном матриксе — мезоглее . Предполагают, что для животных группы Eumetazoa клетки по умолчанию являются эпителиоцитами. Несмотря на существенное морфологическое и функциональное разнообразие эпителиев у животных, ключевые белки, необходимые для дифференцировки эпителиев и формирования контактов между эпителиальными клетками, одинаковы у всех Eumetazoa и появились рано в ходе эволюции .
Иногда в результате ответа на стресс, хроническое воспаление и другие неблагоприятные стимулы происходит эпителиальная метаплазия — обратимое превращение дифференцированных клеток эпителия одного типа в дифференцированные клетки эпителия другого типа. Наиболее часто наблюдается превращение призматического эпителия в столбчатый. Плоскоклеточная метаплазия часто происходит в псевдомногослойном эпителии трахеи и бронхов в ответ на длительное воздействие табачного дыма. При хронических протозойных инфекциях и шистосомозе может происходить плоскоклеточная метаплазия уротелия. Иногда происходит обратное превращение плоского эпителия в столбчатый, например, при гастроэзофагальном рефлюксе многослойный плоский неороговевающий эпителий нижней части пищевода превращается в столбчатый эпителий с бокаловидными клетками, похожий на кишечный эпителий. Метаплазия — обратимое преобразование, и, когда действие неблагоприятного фактора, вызвавшего метаплазию, прекращается, эпителий вновь принимает свой исходный вид .
Некоторые патогены нарушают избирательную проницаемость эпителиев, разрушая межклеточные контакты между эпителиоцитами. Например, патогенная бактерия Clostridium perfringens выделяет , который разрушает плотные контакты, связывая молекулы их важнейшего компонента — белка клаудина . Helicobacter pylori также разрушает плотные контакты в выстилке желудка. Целостность плотных контактов также нарушают некоторые РНК-содержащие вирусы , вызывающие энтериты у детей, и паразиты , такие как клещи .
Злокачественные опухоли , происходящие из эпителиальных тканей, называют карциномами . Карциномы могут формироваться как во взрослом организме, так и во время эмбриогенеза , однако у детей карциномы чрезвычайно редки . По типу поражённого эпителия выделяют аденокарциномы (опухоли из железистого эпителия), (опухоли из плоского эпителия), , анапластические карциномы, и .

Термин «эпитель» впервые использовал голландский учёный Фредерик Рюйш в 1703 году в своей работе Thesaurus Anatomicus . Он обозначил этим термином ткань, которую обнаружил на губах трупа. В середине XIX века швейцарский естествоиспытатель Альбрехт фон Галлер адаптировал термин, предложенный Рюйшем, в вариант «эпителий». Первая попытка классифицировать эпителии была предпринята Уильямом Шарпеем . Классификация Шарпея была приведена в семнадцатом издании труда Quain’s Elements of Anatomy , опубликованном в 1867 году. Шарпей подразделил эпителиальные ткани на физиологические категории, такие как эпидермис, железистый эпителий, слизистый эпителий, сосудистый эпителий, а также выделил основные морфологические типы эпителиев: столбчатые, сферические, «клетчатые», ресничные, плоские. Хотя многие термины, введённые Шарпеем, в настоящее время не используются, современная классификация основана на той, что предложил Шарпей. Двумя годами ранее публикации классификации и систематизированного описания эпителиев Шарпеем Вильгельм Гис предложил термин «эндотелий» для обозначения эпителия, выстилающего кровеносные сосуды, а в 1890 году американский исследователь предложил называть эпителий, покрывающий внешнюю поверхность тела, термином «эктотелий», а эпителий, выстилающий полости тела, — термином «мезотелий». Термины эндотелий и мезотелий используются и по сей день. В 1870 году Немецкий учёный Генрих Вильгельм Вальдейер высказал гипотезу, что клетки зародышевой линии происходят из эпителия, однако в дальнейшем эта гипотеза не подтвердилась. В 1960-х годах было доказано, что эпителий участвует в эмбриональном развитии многих органов .
|
|
|
|---|---|
| Словари и энциклопедии |
|
| В библиографических каталогах |






