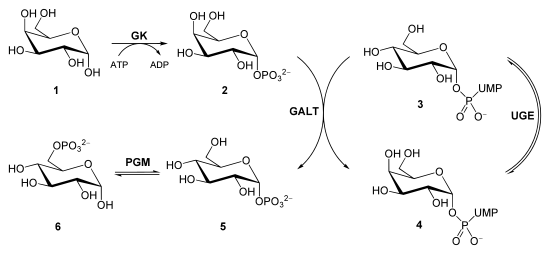
Гликолиз
- 1 year ago
- 0
- 0

Глюконеогене́з — метаболический путь , приводящий к образованию глюкозы из неуглеводных соединений (в частности, пирувата ). Наряду с гликогенолизом , этот путь поддерживает в крови уровень глюкозы, необходимый для работы многих тканей и органов, в первую очередь, нервной ткани и эритроцитов . Он служит важным источником глюкозы в условиях недостаточного количества гликогена , например, после длительного голодания или тяжёлой физической работы . Глюконеогенез является обязательной частью цикла Кори , кроме того, этот процесс может быть использован для превращения пирувата , образованного при дезаминировании аминокислот аланина и серина .
Суммарное уравнение глюконеогенеза выглядит следующим образом:
Глюконеогенез протекает в основном в печени , но менее интенсивно он протекает также в и слизистой кишечника .
Глюконеогенез и гликолиз регулируются реципрокно: если клетка снабжена энергией в достаточной степени, то гликолиз приостанавливается, и запускается глюконеогенез; напротив, при активизации гликолиза происходит приостановление глюконеогенеза .
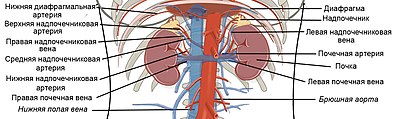
Глюконеогенез происходит у животных , растений , грибов и микроорганизмов . Его реакции одинаковы для всех тканей и биологических видов . Важными предшественниками глюкозы у животных выступают трёхуглеродные соединения, такие как лактат , пируват, глицерол , а также некоторые аминокислоты . У млекопитающих глюконеогенез происходит главным образом в печени , в меньшей степени — в корковом слое почек и эпителиальных клетках, выстилающих тонкую кишку . Образованная в ходе глюконеогенеза глюкоза уходит в кровь, откуда доставляется к другим тканям. После интенсивной физической работы лактат, образовавшийся при анаэробном гликолизе в скелетных мышцах , возвращается в печень и превращается там в глюкозу, которая снова поступает в мышцы или превращается в гликоген (этот круговорот известен как цикл Кори ). У проростков растений запасённые в семени жиры и белки превращаются, в том числе и через глюконеогенез, в дисахарид сахарозу , который транспортируется по всему развивающемуся растению. Глюкоза и её производные служат предшественниками для синтеза растительной клеточной стенки , нуклеотидов , коферментов и многих других жизненно важных метаболитов . У многих микроорганизмов глюконеогенез начинается с простых органических соединений, содержащих два или три атома углерода, например, ацетата , лактата и пропионата , которые содержатся в питательной среде .
Хотя реакции глюконеогенеза одинаковы у всех организмов, соседние метаболические пути и регуляторные пути глюконеогенеза отличаются у различных видов и в различных тканях . В этой статье рассмотрены особенности глюконеогенеза млекопитающих; о механизмах синтеза глюкозы растениями из первичных продуктов фотосинтеза см. Фотосинтез .
Глюконеогенез и гликолиз не являются полностью идентичными процессами, протекающими в противоположных направлениях, хотя несколько стадий являются общими для обоих процессов: 7 из 10 ферментативных реакций глюконеогенеза обратны соответствующим реакциям гликолиза. Однако 3 реакции гликолиза необратимы in vivo и не могут использоваться в глюконеогенезе: образование глюкозо-6-фосфата из глюкозы под действием фермента гексокиназы , фосфорилирование фруктозо-6-фосфата до фруктозо-1,6-бисфосфата фосфофруктокиназой-1 (PFK-1), а также превращение фосфоенолпирувата в пируват под действием пируваткиназы . В клеточных условиях эти реакции имеют большое отрицательное изменение энергии Гиббса , в то время как другие реакции гликолиза имеют ΔG около 0. При глюконеогенезе три необратимые стадии гликолиза заменены «обходными» реакциями, катализируемыми другими ферментами, и эти реакции также очень экзергоничны и потому необратимы. Таким образом, в клетках как гликолиз, так и глюконеогенез являются необратимыми процессами. У животных гликолиз происходит только в цитозоле , как и большая часть реакций глюконеогенеза, хотя некоторые его реакции происходят в митохондриях и эндоплазматическом ретикулуме . Это даёт возможность для их координированной и взаимно обратной регуляции. Регуляторные механизмы, различающиеся у гликолиза и глюконеогенеза, действуют на ферментативные реакции, уникальные для каждого процесса .
Ниже представлена схема реакций глюконеогенеза:

Ниже рассмотрены 3 стадии глюконеогенеза, отличные от реакций гликолиза, проведённых в обратном направлении.

Первой реакцией глюконеогенеза является превращение пирувата в фосфоенолпируват (ФЕП). Эта реакция не может быть обратной пируваткиназной реакции гликолиза, поскольку пируваткиназная реакция имеет большое отрицательное изменение энергии Гиббса и потому необратима в клеточных условиях. Вместо этого фосфорилирование пирувата осуществляется «окольным путём», для реакций которого у эукариот необходимы и цитозольные, и митохондриальные ферменты .
Вначале пируват переносится из цитозоля в митохондрии или образуется в митохондриях из аланина путём трансаминирования , при котором α-аминогруппа переносится с аланина на α-кетокарбоновую кислоту. После этого митохондриальный фермент , для активности которой необходим кофермент биотин , превращает пируват в оксалоацетат :
В этой реакции карбоксилирования участвует биотин как переносчик активированного бикарбоната . HCO 3 - фосфорилируется с затратой ATP с образованием смешанного ангидрида (карбоксифосфата). После этого на место фосфата в карбоксифосфате присоединяется биотин. Справа показан механизм этой реакции .
Пируваткарбоксилаза является первым регулируемым ферментом глюконеогенеза, её позитивным эффектором служит ацетил-КоА (ацетил-КоА образуется при β-окислении жирных кислот , повышение его концентрации сигнализирует о доступности жирных кислот как энергетического ресурса). Кроме того, пируваткарбоксилазная реакция поставляет промежуточные соединения в другой центральный метаболический путь — цикл трикарбоновых кислот .
Так как в митохондриальной мембране отсутствуют белки-переносчики оксалоацетата, до экспорта в цитозоль образовавшийся при пируваткарбоксилазной реакции оксалоацетат должен быть обратимо восстановлен в малат митохондриальным ферментом малатдегидрогеназой с затратой NADH:
Стандартное изменение энергии Гиббса для этой реакции довольно велико, однако при физиологических условиях (среди которых и очень низкая концентрация оксалоацетата) её ΔG ≈ 0, поэтому эта реакция обратима. Митохондриальная малатдегидрогеназа участвует и в глюконеогенезе, и в цикле трикарбоновых кислот, осуществляя и прямую, и обратную реакции . Оксалоацетат может также переноситься из митохондрии в цитозоль после трансаминирования в аспартат .
Малат покидает митохондрию через специальный белок-транспортер на внутренней митохондриальной мембране , и в цитозоле он вновь окисляется до оксалоацетата с образованием цитозольного NADH:
После этого оксалоацетат превращается в фосфоенолпируват под действием фосфоенолпируваткарбоксикиназы . В этой Mg 2+ -зависимой реакции донором фосфорильной группы выступает GTP:
В клеточных условиях эта реакция обратима; образование фосфоенопирувата компенсируется гидролизом другого высокоэнергетичного фосфатсодержащего соединения — GTP .
Общее уравнение для двух первых «обходных» реакций гидролиза выглядит следующим образом:
Два высокоэнергетичных фосфатных эквивалента (один от ATP и другой GTP), каждый из которых может дать 50 кДж / моль в клеточных условиях, затрачиваются для фосфорилирования одной молекулы пирувата с образованием фосфоенолпирувата. Однако в соответствующей реакции гликолиза (при образовании пирувата из ФЕП) образуется лишь одна молекула ATP из ADP. Хотя стандартное изменение энергии Гиббса ΔG' o в двухшаговом превращении пирувата в фосфоенолпируват равно 0,9 кДж/моль, реальное изменение энергии Гиббса (ΔG), посчитанное с учётом внутриклеточных концентраций соединений, имеет большое отрицательное значение (—25 кДж/моль). Причиной этого является быстрое использование фосфоенолпирувата в других реакциях, так что его концентрация остаётся относительно низкой. По этой причине образование ФЕП из пирувата под действием глюконеогенетических ферментов в клеточных условиях необратимо .
Тот же СО 2 , который присоединяется к пирувату в ходе пируваткарбоксилазной реакции, выделяется при фосфоенолпируваткарбоксикиназной реакции. Такое карбоксилирование-декарбоксилирование является путём «активации» пирувата, то есть декарбоксилирование оксалоацетата способствует образованию фосфоенолпирувата .
Отношение [NADH]/[NAD + ] в цитозоле равно 8 × 10 4 , что примерно в 10 5 раз меньше, чем в митохондриях. Поскольку цитозольный NADH используется в глюконеогенезе (при образовании глицеральдегид-3-фосфата из 1,3-бисфосфоглицерата ), биосинтез глюкозы не может происходить, если нет доступного NADH. Транспорт малата из митохондрии в цитозоль и превращение его в оксалоацетат в цитозоле эффективно переносит восстановительные эквиваленты в цитозоль, где их недостаточно. Таким образом, такой путь от пирувата к ФЕП обеспечивает важный баланс между потребляемым и образуемым в цитозоле NADH в ходе глюконеогенеза .
Выше отмечалось, что, кроме пирувата, предшественником для глюконеогенеза также может выступать лактат. Этот путь обеспечивает использование лактата, образовавшегося, например, при гликолизе в эритроцитах или в мышцах при анаэробных условиях. Особенно этот путь важен для крупных позвоночных после тяжёлой физической работы. Преобразование лактата в пируват в цитозоле гепатоцитов приводит к образованию NADH, поэтому в экспорте восстановительных эквивалентов (например, малата) из митохондрий здесь нет нужды. После того как пируват, образовавшийся в лактатдегидрогеназной реакции, транспортируется в митохондрии, он превращается в оксалоацетат под действием пируваткарбоксилазы, как писалось выше. Этот оксалоацетат, однако, превращается непосредственно в фосфоенолпируват митохондриальным изоферментом фосфоенолпируваткарбоксикиназой, а ФЕП выводится из митохондрий в цитозоль, где осуществляются дальнейшие реакции глюконеогенеза .
У растений и некоторых бактерий было обнаружено два фермента, способных образовывать ФЕП непосредственно из пируватата. К их числу относится бактерии Escherichia coli . При работе этого фермента его остаток гистидина связывает пирофосфатную группу, взятую от ATP. Далее пирофосфатная группа гидролизуется с выделением фосфата и образованием соединения фермент-His-P. Последний взаимодействует с пируватом, образуя ФЕП. Похожий механизм присущ , которая впервые была описана у тропических злаков и играет важную роль в С 4 -фотосинтезе , а также задействована в глюконеогенезе у Acetobacter . Отличие этого фермента от фосфоенолпируватсинтазы заключается лишь в том, что атакующей частицей является не вода, а неорганический фосфат .
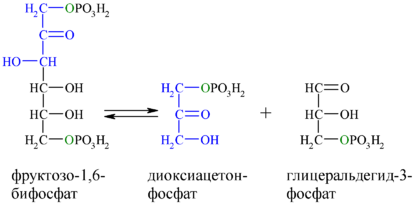
Второй реакцией гликолиза, которая не может дублироваться обратной реакцией в глюконеогенезе, является фосфорилирование фруктозо-6-фосфата фосфофруктокиназой-1. Так как эта реакция высоко экзергонична и поэтому необратима в клеточных условиях, образование фруктозо-6-фосфата из фруктозо-1,6-бисфосфата катализируется другим ферментом — Mg 2+ -зависимой фруктозо-1,6-бисфосфатазой-1 (FBPаза-1), которая катализирует необратимый гидролиз фосфата при первом атоме углерода (а не перенос фосфорильной группы на ADP):
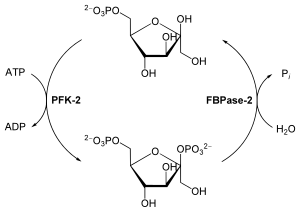
Кроме фруктозо-1,6-бисфосфатазы-1 существует фруктозо-1,6-бисфосфатаза-2, выполняющая регуляторные функции .
Третья «обходная» реакция является последней реакцией глюконеогенеза: дефосфорилирование глюкозо-6-фосфата с образованием глюкозы. Если бы гексокиназа осуществляла эту обратную реакцию, то ей бы сопутствовал перенос фосфорильной группы с глюкозо-6-фосфата на ADP с образованием ATP, что энергетически невыгодно. Реакция, катализируемая глюкозо-6-фосфатазой , не включает синтеза ATP и представляет собой простой гидролиз фосфатного эфира:
Этот Mg 2+ -зависимый фермент встречается на люменальной стороне эндоплазматического ретикулума гепатоцитов, в клетках почек и эпителиальных клетках тонкой кишки, однако в других тканях его нет, поэтому остальные ткани неспособны поставлять глюкозу в кровь. Если бы в них была глюкозо-6-фосфатаза, то она бы гидролизовала глюкозо-6-фосфат, который необходим этим тканям для гликолиза. Глюкоза, образовавшаяся в ходе глюконеогенеза в печени и почках или поглощённая с пищей разносится по кровотоку к этим тканям, в числе которых — мозг и мышцы .
Суммарное уравнение биосинтетических реакций глюконеогенеза, приводящих к образованию глюкозы из пирувата, выглядит так:
На каждую молекулу глюкозы, образовавшуюся из пирувата, необходимо 6 высокоэнергетичных фосфатных групп, 4 из которых берутся от ATP и 2 — от GTP. Кроме того, для восстановления двух молекул 1,3-бисфосфоглицерата необходимы 2 молекулы NADH. В то же время для гликолиза нужны лишь 2 молекулы ATP. По этой причине синтез глюкозы из пирувата является затратным процессом. Большая часть затрачиваемой энергии обеспечивает необратимость глюконеогенеза. В клеточных условиях суммарное изменение энергии Гиббса при гликолизе составляет −63 кДж/моль, а при глюконеогенезе — −16 кДж/моль. Таким образом, в клеточных условиях и гликолиз, и глюконеогенез необратимы .
Описанный выше биосинтетический путь образования глюкозы относится к синтезу глюкозы не только из пирувата, но также 4-, 5- и 6-углеродных промежуточных соединений цикла трикарбоновых кислот. Цитрат , изоцитрат , α-кетоглутарат , , сукцинат , фумарат и малат — все промежуточные продукты цикла трикарбоновых кислот могут окисляться до оксалоацетата. Некоторые или все атомы углерода большей части аминокислот могут быть катаболизированы в пируват или промежуточные соединения цикла трикарбоновых кислот. Поэтому эти аминокислоты могут подвергнуться превращению в глюкозу и называются . Аланин и глутамин — важнейшие молекулы, переносящие аминогруппы в печень из других тканей — служат особенно важными глюкогенными аминокислотами у млекопитающих. После того, как эти аминокислоты отдают свои аминогруппы в митохондриях печени, их углеродные скелеты (пируват и α-кетоглутарат соответственно) вовлекаются в глюконеогенез . Аминокислоты образуются при распаде белков мышц и соединительной ткани , их включение в глюконеогенез происходит при продолжительном голодании или длительной физической нагрузке .
У растений, дрожжей и многих бактерий имеется путь, позволяющий получать углеводы из жирных кислот — глиоксилатный цикл . У животных ключевых ферментов этого цикла нет, и, ввиду необратимости пируватдегидрогеназной реакции , они не могут получать пируват из ацетил-КоА, а значит, образовывать углеводы из жирных кислот (следовательно, и из липидов ). Тем не менее, они могут использовать для глюконеогенеза те небольшие количества глицерола, который образуется при распаде жиров. При этом глицерол фосфорилируется , далее следует окисление центрального атома углерода с образованием дигидроксиацетонфосфата , который является промежуточным соединением глюконеогенеза .
Глицеролфосфат является необходимым промежуточным соединением при синтезе жиров ( триглицеридов ) в адипоцитах , однако эти клетки лишены глицеролкиназы и поэтому не могут осуществлять фосфорилирование глицерола. Вместо этого адипоциты могут осуществлять сокращённый вариант глюконеогенеза, известный как глицеронеогенез : преобразование пирувата в дигидроксиацетонфосфат через первые реакции глюконеогенеза, вслед за которым следует восстановление дигидроксиацетонфосфата до глицеролфосфата .
Если бы гликолиз и глюконеогенез протекали одновременно и на большой скорости, то результатом стали бы расход ATP и образование тепла . Например, фосфофруктокиназа-1 и фруктозо-1,6-бисфосфатаза-1 катализируют противоположные реакции:
Суммой этих двух реакций является
Эти две ферментативные реакции, как и ряд других реакций этих двух путей, регулируются аллостерически и ковалентных модификаций. Гликолиз и глюконеогенез регулируются реципрокно, то есть если поток глюкозы, проходящей через гликолиз, растёт, то поток пирувата, проходящего через глюконеогенез, спадает, и наоборот . Например, FBPаза-1 строго подавляется аллостерическим связыванием AMP , так что когда клеточные запасы ATP невелики, а уровень AMP высок, ATP-зависимый синтез глюкозы приостанавливается, а катализирующая соответствующую реакцию гликолиза PFK-1, наоборот, активируется AMP . Хотя PFK-1 активируется фруктозо-2,6-бисфосфатом , на FBPазу-1 это соединение имеет противоположный эффект: он снижает его сродство к субстратам и тем самым замедляет глюконеогенез .
Транскрипционные факторы и глюконеогенез


На пути преобразования пирувата в глюкозу первой контрольной точкой, на которой определяется дальнейшая судьба пирувата в митохондрии, является то, будет ли он превращён в ацетил-КоА пируватдегидрогеназным комплексом с дальнейшим вовлечением в цикл трикарбоновых кислот или же в оксалоацетат под действием пируваткарбоксилазы, чтобы начать глюконеогенез. Когда в качестве источника энергии доступны жирные кислоты, то при их разложении в митохондриях образуется ацетил-КоА, выступающий в качестве сигнала о том, что в дальнейшем окислении глюкозы нет нужды. ацетил-КоА являются положительным аллостерическим модулятором пируваткарбоксилазы и отрицательным модулятором пируватдегидрогеназного комплекса; его действие опосредуется стимуляцией протеинкиназы, которая инактивирует дегидрогеназу. Когда энергетические потребности клетки удовлетворены, окислительное фосфорилирование замедляется, концентрация NADH по сравнению с NAD + увеличивается, цикл трикарбоновых кислот подавляется, и происходит накопление ацетил-КоА. Повышенная концентрация ацетил-КоА подавляет пируватдегидрогеназный комплекс, тем самым замедляя образование ацетил-КоА из пирувата и стимулируя глюконеогенез через активацию пируваткарбоксилазы, что позволяет превратить излишек пирувата в оксалоацетат (а впоследствии и глюкозу) .
Полученный таким образом оксалоацетат превращается в фосфоенолпируват под действием фосфоенолпируваткарбоксикиназы. У млекопитающих регуляция этого важнейшего фермента глюконеогенеза осуществляется на уровне его синтеза и распада под влиянием режима питания и гормональных сигналов. Так, его промотор имеет 15 или более регуляторных элементов, распознаваемых по крайней мере 12-ю известных транскрипционных факторов , и, как предполагается, ещё большим числом пока не описанных. Голодание или высокий уровень глюкагона увеличивают транскрипцию этого фермента и стабилизируют его мРНК . Действие глюкагона опосредовано транскрипционным фактором CREB ( англ. cyclic AMP response element binding protein ), который активирует синтез глюкозо-6-фосфатазы и фосфоенолпируваткарбоксилазы в ответ на вызванное глюкагоном увеличение внутриклеточной концентрации cAMP . Инсулин или высокое содержание глюкозы в крови имеют противоположный эффект. Эти изменения, вызываемые в основном внеклеточными сигналами (питание, гормоны), могут длиться от нескольких минут до нескольких часов . Инсулин также замедляет экспрессию генов глюкозо-6-фосфатазы и фруктозо-1,6-бисфосфатазы. Другим транскрипционным фактором, регулирующим экспрессию генов ферментов глюконеогенеза, является FOXO1 ( англ. forkhead box other ). Инсулин активирует протеинкиназу В , которая фосфорилирует FOXO1, находящийся в цитозоле. С фосфорилированным FOXO1 связывается убиквитин , и FOXO1 разрушается в протеасоме , однако в отсутствие фосфорилирования или при дефосфорилировании FOXO1 может проникать в ядро , связываться с соответствующим регуляторным элементом на ДНК и запускать транскрипцию генов фосфоенолпируваткарбоксикиназы и глюкозо-6-фосфатазы. Фосфорилированию FOXO1 протеинкиназой В препятствует глюкагон .
При снижении использования лактата в качестве субстрата для глюконеогенеза, которое может вызываться дефектом ферментов глюконеогенеза, концентрация лактата в крови повышается, что приводит к понижению pH крови и развитию лактатацидоза . Кратковременный лактатацидоз встречается и у здоровых людей при интенсивной мышечной работе, в этом случае он компенсируется путём гипервентиляции лёгких и ускоренным выведением углекислого газа .
На глюконеогенез существенное влияние оказывает этанол . В результате его катаболизма увеличивается количество NADH, что смещает равновесие в лактатдегидрогеназной реакции в сторону образования лактата, снижению образования пирувата и замедлению глюконеогенеза .