Ракообразные
- 1 year ago
- 0
- 0

Веслоно́гие ракообра́зные , или веслоно́гие , или копепо́ды ( лат. Copepoda ) , — подкласс ракообразных из класса . Один из крупнейших таксонов ракообразных (по разным данным, число видов копепод составляет от 10 до 20 тысяч). Наука, изучающая копепод — (раздел карцинологии ).
Большинство групп копепод — эктопаразиты беспозвоночных и позвоночных животных. Свободноживущие представители отрядов Calanoida (каланоиды) и Cyclopoida (циклопоиды) — важнейший компонент зоопланктона морей и континентальных водоёмов. Виды отряда Harpacticoida (гарпактициды) — обычные представители бентоса и , реже встречаются в планктоне. Ниже рассматриваются в основном строение и образ жизни свободноживущих представителей копепод.
Существует Всемирное общество копеподологов ( англ. World Association of Copepodologists ), издающее информационный бюллетень « Monoculus copepod newsletter » .

Размеры большинства видов составляют от 1 до 10 мм, есть более мелкие виды и более крупные (до 2—3 см) глубоководные и паразитические. Паразитические копеподы часто достигают намного более внушительных размеров, так например Kroyeria caseyi достигает 6,5 см .


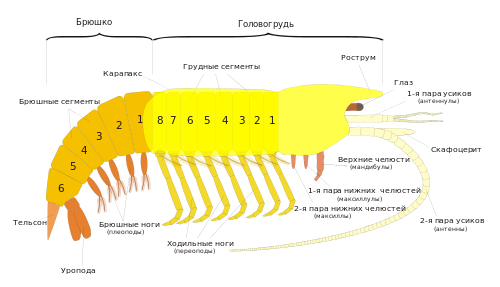
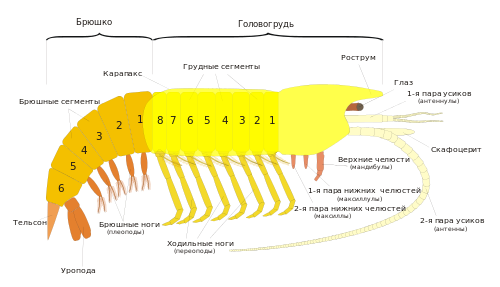
Тело копепод делится на три тагмы : голову — цефалосому (в копеподологии иногда её называют цефалоторакс, головогрудь), грудь (торакс) и брюшко (абдомен). При этом тельсон (анальную лопасть) многие копеподологи называют последним брюшным (анальным) сегментом.
Тело копепод может «складываться» пополам, изгибаясь в сагиттальной плоскости. При этом граница между функционально передним отделом тела (просомой) и функционально задним (уросомой) у циклопоид и гарпактицид проходит между сегментами груди, несущими четвёртую и пятую пары ног. Эти группы объединяют под названием «Podoplea» — «ногобрюхие». У каланоид граница между просомой и уросомой проходит позади сегмента, несущего пятую пару ног, поэтому их называют «Gymnoplea» — «голобрюхие». Этому признаку, который хорошо коррелирует с другими особенностями строения, придаётся высокий таксономический вес, а Podoplea и Gymnoplea рассматриваются как таксономические категории (в современных классификациях копепод — как надотряды).
Голова несёт одноветвистые антенны первые (антеннулы, усы), двуветвистые антенны вторые (антенны), мандибулы , максиллы первые (максиллулы), максиллы вторые ( максиллы ) и максиллипеды ( ногочелюсти ) — придатки сросшегося с головой первого грудного сегмента. У представителей большинства семейств гарпактицид и у некоторых представителей других отрядов с головой сливается и следующий сегмент груди, несущий плавательные конечности, которые при этом могут сильно видоизменяться.
На голове, между мандибулами, располагается ротовое отверстие, прикрытое спереди крупной верхней губой, а сзади — небольшой нижней губой. На переднем крае головы находится направленный вниз вырост — рострум, иногда отчленённый. Антенны I (антеннулы) всегда одноветвистые. Число их члеников различается у представителей разных отрядов. Так, у гарпактицид обычно 5—8 члеников (у самцов до 14); у большинства каланоид 21—27 члеников; у циклопоид от 9 до 23 члеников. У типичных представителей различается относительная длина антеннул: у каланоид они примерно равны телу, у циклопоид равны головогруди, а у гарпактицид заметно короче головогруди. Антенны I участвуют в локомоции , а также несут сенсиллы .
Антенны II обычно двуветвистые (у многих циклопоид одноветвистые), участвуют в создании токов воды для плавания и питания. Мандибула подразделяется на коксу, которая образует жевательный вырост (гнатобазу) с зубцами и щетинками, и щупик, который исходно состоит из базиса, экзо- и эндоподита. Часто ветви, а иногда и основание щупика редуцируются. Так, у многих циклопов от мандибулы отходят лишь три щетинки, которые считаются рудиментом щупика. На жевательных зубцах мандибулы многих морских копепод есть «коронки» из оксида кремния, которые помогают им разгрызать прочные домики диатомовых водорослей.
На следующих за ногочелюстным сегментом четырёх сегментах груди располагаются двуветвистые плавательные конечности — ноги уплощённой формы, служащие основными двигателями при плавании, за наличие которых отряд и получил своё название. Плавательная конечность состоит из двучленикового , базальный (проксимальный) членик которого называется кокса, а дистальный — базис, и двух отходящих от базиса ветвей (иногда считается, что в состав протоподита входит ещё один членик — прекокса, который слабо отчленён от тела). Наружная (экзоподит) и внутренняя (эндоподит) ветви состоят из 2—4 члеников каждая и несут длинные щетинки, покрытые длинными тонкими выростами (сетулами), и более короткие шипы.
На последнем сегменте груди расположена пятая пара грудных ног, которые обычно не участвуют в плавании и у многих групп сильно уменьшены или видоизменены. У самцов большинства семейств каланоид они резко асимметричны. Эндоподиты обеих ног часто рудиментарны, экзоподит одной из ног служит для переноса сперматофора к семяприёмникам самки при спаривании, а более крупный экзоподит другой конечности несёт длинный когтевидно изогнутый шип, участвующий в удержании самки. Строение и вооружение пятой пары ног для циклопоид и каланоид служит важнейшим таксономическим признаком.
Брюшко состоит обычно из 2—4 сегментов (не считая тельсона). На первом сегменте брюшка находятся парные половые отверстия. У гарпактицид и циклопоид на нём расположена рудиментарная шестая пара ног, у каланоид он лишён конечностей. Остальные сегменты брюшка конечностей не несут. На тельсоне располагаются два подвижных придатка — вилочка, или фурка (фуркальные ветви). Эти придатки состоят из одного членика и не гомологичны конечностям. На фурке находятся фуркальные щетинки, длина и расположение которых — важный таксономический признак.
Как правило, у самок сливаются первый и второй сегменты брюшка, образуя крупный генитальный сегмент; у самцов такого слияния не происходит, поэтому у самцов на один брюшной сегмент больше, чем у самок. У представителей Cyclopoida и Harpacticoida самцы обычно заметно мельче самок, имеют крючковидно изогнутые, укороченные антенны I, служащие для схватывания и удержания самок при спаривании. У многих Calanoida самки и самцы по размерам не различаются. У самцов имеется одна видоизменённая антенна I, которая называется геникулирующая антенна. Она расширена в средней части и способна «складываться пополам»; как и у циклопов , она служит для удержания самки при спаривании. В отдельных случаях наблюдается половой диморфизм в строении практически любых пар конечностей и сегментов тела .
|
|
Этот раздел статьи
ещё
не написан
.
|
Центральная нервная система состоит из головного мозга и соединённой с ним окологлоточным нервным кольцом брюшной нервной цепочки. От головного мозга отходит непарный нерв к науплиальному глазу и парные нервы к фронтальному органу, а также нервы к антеннулам и антеннам (последние — от тритоцеребрума). В состав подглоточного ганглия входят ганглии мандибул, первых и вторых максилл. Ганглии брюшной нервной цепочки слабо отграничены друг от друга. Вся брюшная нервная цепочка расположена в головогруди, в брюшко она не заходит .
У многих видов копепод аксоны окружены миелиновой оболочкой , что встречается редко среди беспозвоночных (другие примеры — некоторые аннелиды и высшие ракообразные , такие как креветки -палемониды и пенеиды). Более того, у копепод миелин высоко организован и напоминает своей многослойной структурой таковой челюстноротых позвоночных.
|
|
Этот раздел статьи
ещё
не написан
.
|
Узкий эктодермальный пищевод направлен от ротового отверстия к спинной стороне тела и назад. Он переходит в расширенную энтодермальную среднюю кишку, которая, постепенно сужаясь, тянется через всю головогрудь и большую часть брюшка. В задней части брюшка средняя кишка переходит в короткую эктодермальную заднюю кишку, которая открывается анальным отверстием на заднем крае тельсона, между основаниями фуркальных ветвей.
|
|
Этот раздел статьи
ещё
не написан
.
|
У каланоид в первом свободном грудном сегменте над кишечником расположено яйцевидное сердце, снабжённое одной парой — отверстий, через которые в него попадает гемолимфа . Кровеносных сосудов нет, через отверстие в передней части сердца гемолимфа изливается в полость тела. У представителей большинства других отрядов копепод отсутствует и сердце, так что (редкий случай среди членистоногих) кровеносная система полностью редуцирована. При этом у циклопоид гемолимфа (полостная жидкость) перебалтывается в полости тела за счёт движений кишечника, который поднимают и опускают специальные мышцы.
У копепод полностью отсутствуют специальные органы дыхания, что довольно необычно для членистоногих. Мелкие размеры и обилие выростов, увеличивающие относительную площадь поверхности, а также очень небольшая толщина хитинового покрова позволяют копеподам дышать всей поверхностью тела .
На личиночных стадиях органами выделения служат парные антеннальные (расположенные в сегменте антенн вторых) железы, а на взрослой стадии — максиллярные (расположенные в сегменте вторых максилл). Считается, что и те, и другие представляют собой видоизменённые целомодукты . Они состоят из замкнутого мешочка (остатка целомической полости) и извитого выводного канала.
|
|
Этот раздел статьи
ещё
не написан
.
|
|
|
Этот раздел статьи
ещё
не написан
.
|
Многие копеподы — типичные представители зоопланктона , имеющие характерные адаптации к жизни в толще воды. Это прежде всего приспособления, позволяющие удерживаться в толще воды с минимальными затратами энергии: мелкие размеры (за счёт этого увеличивается относительная площадь поверхности и отношение величины силы трения к массе тела); наличие увеличивающих площадь трения о воду выростов (антеннулы, щетинки антеннул и фурки); тонкие покровы; запасание жира, уменьшающего плотность тела.
Большинство свободноживущих копепод питаются одноклеточными или мелкими колониальными водорослями , которых они отфильтровывают в толще воды, а также донными диатомовыми , бактериями и детритом, которые они могут собирать или соскабливать на дне. Многие виды каланоид и циклопоид — хищники, поедающие другие виды ракообразных (молодь копепод и кладоцер ), коловраток , личинок насекомых I—II возраста (в том числе личинок хирономид и кулицид ) и др. Копеподитные стадии некоторых пресноводных циклопоид залезают в выводковые камеры дафний , где пожирают яйца .
При более детальном изучении «фильтрационного» питания копепод с помощью скоростной микрокиносъёмки выяснилось, что многие из них «охотятся» на отдельные клетки водорослей, которые отлавливают поодиночке. Питающиеся водорослями копеподы запасают энергию пищи в жировых каплях, которые содержатся в их тканях и часто окрашены в желтовато-оранжевый цвет. У полярных видов, питающихся в первую очередь диатомовыми, в период массового весеннего «цветения» объём жировых запасов может достигать половины объёма тела.
Большинство видов копепод — раздельнополые животные. Практически всем им свойственен половой диморфизм , особенно ярко выраженный у многих паразитических видов.
Спариванию предшествует сложное половое поведение, в котором, видимо, обычно играют важную роль как хеморецепция, так и механорецепция. Самки копепод выделяют половые феромоны , которые воспринимаются самцами при помощи хемосенсорных щетинок (эстетасков) первых антенн .
При спаривании у большинства семейств каланоид самец сперва захватывает самку за тельсон или фуркальные ветви при помощи геникулирующей антенны, затем — за участок тела, расположенный перед или сразу после генитального сегмента при помощи ног пятой пары, при этом самец и самка обычно располагаются «головой к хвосту» друг друга . Спаривание продолжается от нескольких минут до нескольких суток.
Свободноживущие копеподы имеют сперматофорное оплодотворение. Крупные, сравнимые по размерам с размерами брюшка животного, сперматофоры каланоид переносятся на генитальный сегмент самки во время спаривания при помощи левой пятой ноги самца; на её конце имеются «щипчики», удерживающие бутылковидный сперматофор за суженную базальную часть.
Из яиц копепод выходит личинка ортонауплиус, имеющая три передних пары конечностей — одноветвистые антенны I и двуветвистые антенны II и мандибулы . После первой линьки ортонауплиус превращается в метанауплиуса, у которого имеются зачатки следующей пары конечностей (максилл II).
Светятся представители многих семейств каланоид, а также Oncaea conifera ( Poecilostomatoida ) . У последней светящийся секрет не выделяется наружу, как это обычно происходит у других копепод, но остаётся внутри многочисленных одноклеточных желёз, сообщающихся порами с внешней средой. Эти железы выдают короткие (80—200 мс) вспышки, до 30 вспышек в секунду и до нескольких сотен вспышек подряд до истощения люминесценции.
У всех изученных в этом отношении копепод люциферином служит широко распространённый среди морских организмов целентеразин .
Копеподы играют исключительно важную роль в водных экосистемах и во всей биосфере. Видимо, они имеют наибольшую биомассу среди всех групп водных животных и почти наверняка занимают первое место по доле во водоёмов. Как потребители фитопланктона копеподы являются главными консументами I порядка в морях и пресных водах. Копеподы служат основной пищей для многих других водных животных, от стрекающих и гребневиков до усатых китов .
Поверхностные воды океанов в настоящее время считаются самым крупным поглотителем углекислого газа (2 миллиарда тонн в год — вероятно, около трети антропогенной эмиссии). Во многом поглощение избыточной углекислоты обеспечивается жизнедеятельностью копепод.
Многие планктонные копеподы питаются ночью в поверхностных слоях океана, а днём мигрируют на глубину, чтобы избежать выедания зрительными хищниками. Их трупы, сброшенные экзувии и опускаются в более глубокие слои воды. Это (как и дыхание живых копепод, переваривающих пищу на глубине) способствует перемещению биогенного углерода из верхних слоёв воды в донные осадки. Кроме того, образование в ходе питания копепод фекальных пеллетов способствует очистке верхних слоёв воды от минеральных взвесей. Это увеличивает прозрачность воды и, следовательно, продукцию фитопланктона.

По разным оценкам, от трети до половины всех видов копепод — комменсалы и паразиты разных групп животных.
Хозяевами копепод могут быть сифонофоры , коралловые полипы , моллюски , кольчатые черви , ракообразные , иглокожие , оболочники , а также костные и хрящевые рыбы , амфибии и водные млекопитающие .
Благодаря своей распространённости и широком представительстве среди планктона, копеподы обеспечивают питание многих рыб, тем самым являясь для человека полезными; в то же время, паразитические копеподы поголовье рыб снижают, но это входит в механизм естественного регулирования численности последних [ источник не указан 3776 дней ] . Представители копеподов циклопы являются промежуточными хозяевами опасного паразита человека — широкого лентеца и ришты . Холерный вибрион — бактериальный партнер веслоногих.
В ископаемом состоянии копеподы достоверно известны с каменноугольного периода , следующие по возрасту находки происходят из ранней юры .
В американском мультсериале «Губка Боб Квадратные штаны» присутствует копепод по имени Планктон — главный злодей сериала.
В одной из недавних сводок по систематике ракообразных (Martin, Davis, 2001) предлагается рассматривать копепод в качестве подкласса, который делится на два инфракласса и несколько отрядов:
В предложенной системе копеподы делятся на 218 семейств.
В 2017 году система была значительно переработана, отряд Harpacticoida распался на 2 отряда, а и Cyclopoida стали одним отрядом :
| Copepoda |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Для улучшения этой статьи
желательно
:
|