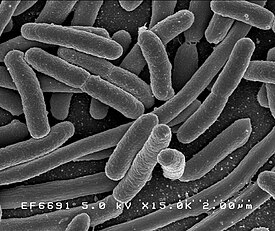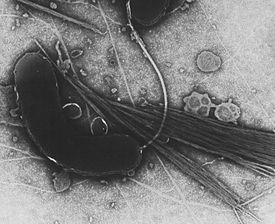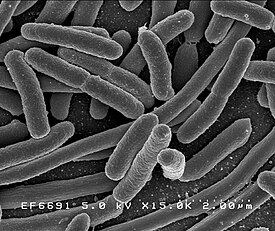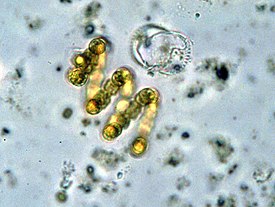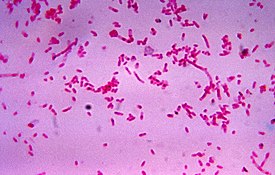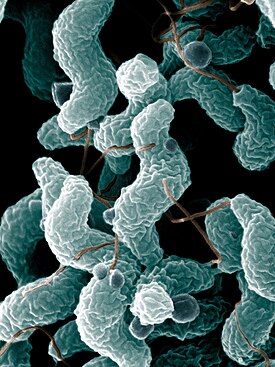
Бактерии
- 1 year ago
- 0
- 0


Сульфатредуци́рующие прокарио́ты (другие названия — десульфатирующие, сульфидогенные) — разнородная группа анаэробных прокариотов ( бактерий и архей ), способных получать энергию в анаэробных условиях за счёт сульфатного дыхания — окисления водорода или других неорганических или органических веществ, используя в качестве конечного акцептора электронов сульфат .
Возраст древнейших ископаемых остатков сульфатредукторов оценивают в 3,5 миллиардов лет, таким образом их рассматривают в числе древнейших живых организмов, участвовавших в вскоре после зарождения жизни на Земле . Предполагается, что сульфатредукторы стали доминирующей формой жизни в ставших анаэробными океанах во время события Великого вымирания на границе пермского и триасового периодов (около 250 млн лет назад) и именно они ответственны за массовое формирование и отложение сероводорода в этот период .
C точки зрения метаболических путей cульфатредукторы — довольно разнообразная группа прокариотов, включающая в том числе и таких представителей, которые наряду с сульфатным дыханием способны реализовывать и другие пути диссимиляции питательных субстратов. Терминальными акцепторами электронов у них также могут становиться фумарат ( фумаратное дыхание ), нитраты и нитриты ( денитрификация ), трёхвалентное железо ( ) и диметилсульфоксид .
Большинство видов рассматриваются как облигатные анаэробы , хотя для ряда представителей показана аэротолерантность — способность переносить присутствие кислорода . Аэротолерантные сульфатредукторы используют ферменты, нейтрализующие активные формы кислорода ( каталазы , супероксиддисмутазы , , ), а также различные способы химического восстановления молекулярного кислорода, в том числе в дыхательной цепи с синтезом АТФ . Тем не менее даже способные к кислородному дыханию сульфатредукторы не могут расти в аэробных условиях, переходя малоактивное состояние .
Диссимиляционное восстановление сульфата (сульфатное дыхание) предполагает, что осуществляющий его организм хемотрофен , то есть использует в качестве источника энергии окислительно-восстановительные реакции . При этом разные сульфатредукторы по отношению к источнику органических веществ оказываются автотрофами и гетеротрофами , а по отношению к природе окисляемому в ходе дыхания субстрату — литотрофами и органотрофами .
Органотрофные представители окисляют широкий круг органических веществ: метан (окисляется до гидрокарбоната ), углеводы , спирты , органические кислоты (в том числе жирные , до C 18 ), аминокислоты и ароматические соединения . В частности, способность некоторых сульфатредукторов к росту на ароматических углеводородах позволяет использовать их для биологической очистки почв и водоносных горизонтов от загрязнения бензолом , толуолом , ксилолом , этилбензолом и некоторыми другими нефтепродуктами .
Большинство литотрофных сульфатредукторов осуществляют окисление молекулярного водорода . Некоторые виды способны окислять угарный газ и железо .
Некоторые виды способны к брожению: пируват до ацетата, малат до сукцината, пропионата и ацетата, сахара до ацетата, этанола, лактата. Во всех случаях [ чего? ] образуются также CO 2 и H 2 .
Конечным акцептором электронов при сульфатном дыхании в большинстве случаев служит ион сульфата . Некоторые виды помимо сульфата способны вовлекать в этот процесс другие соединения серы ( тиосульфат , сульфит , молекулярную серу ), однако часть исследователей не относит эти пути к собственно сульфатному дыханию. В частности, диссимиляционное восстановление серы называют .
Конечным продуктом восстановления сульфата оказывается сульфид (S 2− ) через образование в качестве промежуточного продукта сульфита (S0 3 2− ). Для участия в процессе сульфат активируется молекулой АТФ с образованием аденозинфосфосульфата (АФС) и пирофосфата (ФФ н ). Далее сера в составе АФС восстанавливается с образованием сульфита (S0 3 2− ) и АМФ . В ходе дальнейших реакций сульфит у разных видов прокариотов восстанавливается по одному из двух следующих путей:
Часть реакций протекает на белках и белковых комплексах, связанных с цитоплазматической мембраной. Предполагают также, что существует , способная регенерировать АТФ из АМФ и пирофосфата с затратами энергии протонного градиента.
Сульфатредукция — обратимый процесс. Ряд сульфатредукторов могут осуществлять реакции диспропорционирования промежуточных продуктов (сульфит, тиосульфат) до сульфида и сульфата с выделением энергии .
Известен организм — — способный расти, используя в качестве единственных источников энергии сульфит или тиосульфат. Также эта бактерия способна диспропорционировать серу (на те же продукты, однако эта реакция (ΔG = +10 кДж/моль), видимо, сама по себе не вносит вклада в запасание энергии). Ещё эта бактерия способна с низкой скоростью расти в присутствии метагидроксида железа (FeO(OH)), восстанавливая его до сульфида железа(II) , попутно окисляя сульфид до серы (сумму этой реакции и реакции диспропорционирования серы можно рассматривать как вариант неорганического брожения: сера + гидроксид (3) → сульфид (2) + сульфат, ΔG = −34 кДж/моль в пересчёте на серу).
Некоторые представители группы способны к восстановительному дехлорированию (связь C—Cl восстанавливают до связи C—H):
Известен представитель , способный использовать в качестве акцептора электронов 3-хлорбензойную кислоту и другие м-хлорароматические соединения (восстанавливает их до бензоата).
Представитель способен расти с использованием в качестве донора электронов водорода, и в качестве акцептора — промышленный растворитель тетрахлорэтен (восстанавливает до цис-дихлорэтена).
.png)
Следует различать диссимиляционную сульфатредукцию (сульфатное дыхание) и , служащую не для создания протонного градиента на цитоплазматической мембране и выработки энергии, а для включения серы в состав органических соединений. Данный путь известен для многих организмов — прокариот, грибов, растений. Процесс ассимиляции идёт медленнее диссимиляционного пути и не приводит к накоплению больших количеств сероводорода, а сульфид на выходе включается в состав серосодержащих аминокислот . , задействованные в ассимиляционном процессе, не связаны с клеточными мембранами и представлены глобулярными белками. В ходе реакций ассимиляции сульфата образуются следующие продукты:
На данный момент доказана способность некоторых сульфатредукторов расти автотрофно . Ассимиляция CO 2 осуществляется либо по ацетил-КоА пути , либо по пути восстановительного ЦТК .
Для многих представителей группы характерна способность к азотофиксации.

Продуктами катаболизма органических соединений у разных сульфатредукторов являются либо углекислый газ («полное окисление», обычно происходит либо в реакциях модифицированного ЦТК, либо (чаще) в реакциях обратных ацетогенезу), либо углекислый газ и ацетат («неполное окисление», этот вариант подразумевает окисление органики до ацетил-КоА, который далее окисляться непосредственно не может, и преобразуется в цепи реакций, напоминающих либо «замкнутый» ЦТК, либо обращённый ацетил-КоА-путь ассимиляции углекислого газа) .
Обычно обитают в донных морских осадках (в отличие от метаногенов, растущих обычно в осадках пресных водоёмов) или встречаются в водоёмах, богатых разлагающейся органикой. Входят в состав сульфидогенных микробных сообществ. Большая часть сероводорода на планете генерируется именно в процессах сульфатного дыхания и диссимиляционной сульфаторедукции. Значительная часть органики в донных осадках разлагается именно сульфидогенными сообществами. Представители группы являются важным звеном в глобальном круговороте серы. Это основной источник сульфидов для аноксигенного фотосинтеза других прокарий .
Именно сульфатредукторы ответственны за характерный запах илов и некоторых солончаков, за характерный чёрный оттенок осадочных пород (то есть за наличие в них сульфидов металлов).
Обычно конкурируют с метаногенами за водород и другие субстраты.
Сульфатредукторы способны расти при более низких концентрациях водорода (реакция сульфатредукции энергетически выгоднее) и обычно доминируют над метаногенами, подавляют их. Способны использовать более широкий набор субстратов.
В сообществе сульфатредукторы, в первую очередь, осуществляют сток водорода (что обеспечивает работу синтрофов и поддерживает благоприятный термодинамический баланс сообщества). Немаловажна способность сулльфатредукторов использовать лактат (образуемый молочнокислыми бактериями) — и таким образом предотвращать закисление среды. Ценой за энергетические выгоды, утилизацию метана, лактата и водорода, помощь в утилизации ацетата, жирных кислот, служит образование сероводорода (обычная концентрация 2—3 мМ, это же — порог для существования большинства видов), который угнетает большинство сообществ (за исключением тиофильных, способных существовать при более высоких концентрациях сульфида) .
Для сульфидогенных сообществ важным лимитирующим фактором является приток сульфата извне. Регенерация сульфата в сообществе возможна двумя путями: верхние члены сообщества окисляют сероводород аэробно либо аноксигенные фототрофные бактерии — члены сообщества окисляют его анаэробно.
Говоря о конкуренции метаногенной и сульфидогенной флоры, нужно также упомянуть о существовании «неконкурентного» пути метаболизма одноуглеродных соединений: например, сульфатредукторы могут вырабатывать метанол, который сами далее не используют — и его могут свободно использовать присутствующие в сообществе метаногены.
Накопление сульфида в водоёмах может приводить к замору рыб (равно как и других обитателей этих водоёмов).
Образование биоплёнок (важной частью которых как раз и являются сульфатредукторы) на поверхностях труб в нефтепроводах является одной из существенных проблем транспортировки и добычи нефти .
Масштабы коррозии, вызванной жизнедеятельностью сульфатредукторов, огромны. По подсчётам за 1956 г. ущерб от биологической коррозии труб и конструкций нефтяных сооружений составил около 600 млн. долларов в США, порядка 20 млн. фунтов стерлингов в Англии и порядка 0,2 млн. долларов в Японии.
Коррозии подвергаются не только трубы из сплавов железа , но и иные конструкции из тех же, равно как и из других сплавов (например, алюминия ), а также бетонные конструкции. Показано, что к коррозии в присутствии сульфатредукторов устойчивы олово, цинк, свинец (возможно, ввиду их токсичности) .
Механизм коррозии описывают как . На примере железа и его сплавов (более изученный случай). С одной стороны, железо медленно корродирует в кислой среде и на поверхности металл-раствор образуется плёнка молекулярного водорода, защищающая металл от дальнейшей коррозии. Показано, что не только сульфатредукторы, но и другие бактерии, обладающие гидрогеназами, способны расходовать этот водород в своих метаболитических процессах, что ведёт к ускорению коррозии железа. Дополнительный вклад в ускорение коррозии вносит и то, что ионы железа могут связываться в сульфид или в гидроксид (образуются дополнительные гальванические пары на поверхности металла) . Помимо этого ионы железа могут транспортироваться внутрь клетки и использоваться для метаболитических нужд.
Гидрогеназы — в основном, локализованные в периплазматическом пространстве трансмембранные ферментативные комплексы . Содержат Fe, Ni, Fe-S-кластеры.

Описывают микробиоценозы из железобактерий и сульфатредукторов, ускоряющие коррозию водопроводов. И сообщества из мицелиальных грибов, бактерий рода Pseudomonas и сульфатредукторов, ускоряющих коррозию алюминиевых конструкций .
В силу способности окислять метан до гидрокарбоната в присутствии сульфата, могут мешать добыче метана и его транспортировке.
К группе относят организмы с различными морфологическими, физиологическими, биохимическими признаками, организмы с сильно различными геномами (например, диапазон доли ГЦ-оснований — 37—67 %) .
По состоянию на 2009 год известно 220 видов сульфатредукторов в 60 родах, являющихся представителями пяти неродственных групп .
Сульфатредукторы, окисляющие субстраты полностью, описаны в большом количестве. Остальных называют «неполными окислителями». И эти последние растут гораздо быстрее и легче выделяются из сообществ.
Род (типовый вид A. fulgidus ). Археи с нерегулярной формой клеток. Выделены из горячих морских осадков вблизи гидротермальных подводных источников , из чёрных курильщиков Срединно-Атлантического хребта, из нефтяных скважин Северного моря , из-под замёрзшей поверхности северного склона Аляски . Могут существовать и как авто- и как гетеротрофы. Осуществляют сульфатное и сульфитное дыхание. Не могут использовать в качестве акцепторов электронов нитраты и нитриты. В процессе роста выделяют сероводород и метан (последний образуется иначе, нежели у метаногенов). По результатам секвенирования группа родственна метаногенам. Кроме сульфата акцептором электронов могут быть: элементарная сера, сульфит, тиосульфат, железо (3). Серу не восстанавливают. Донорами электронов могут быть: глюкоза, формиат, формамид, пируват, лактат, ацетат, изопропанол, этанол, фумарат, водород (по некоторым данным — последний не годится для роста с сульфатом) .
Роды и . Найдены близ гидротермальных источников и нефтяных месторождений .