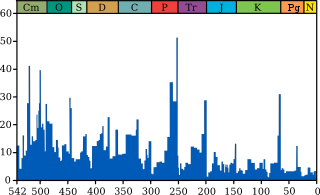
Вымирание в четвертичном периоде
(или
четвертичное вымирание
, также
позднечетвертичное вымирание
) —
массовое вымирание
живого мира (видов животных) в
четвертичном периоде
эры
кайнозоя
. Началось в позднем
плейстоцене
(130 тысяч лет назад), в
Австралии
— около 50 тыс. лет назад, в
Америке
— 15 тыс. лет назад, совпав по времени с
расселением по планете
человека разумного
. Возможные основные причины вымирания —
антропогенное воздействие
и изменение климата
. Отличается высокой скоростью исчезновения видов по всему земному шару. Особенно пострадала
мегафауна
— наземные животные тяжелее 45 килограммов
. Основное отличие от других, более ранних вымираний, это некомпенсированные вымирания крупных видов наземных травоядных, которые были удобной добычей для человека
.
Наиболее быстрое вымирание видов происходит в
голоцене
(начиная с 12 тыс. лет назад и ускоряется в настоящее время, см.
Голоценовое вымирание
)
.
Содержание
Основные события позднечетвертичного вымирания
Масштабы вымирания
Доля вымерших крупных видов млекопитающих (массой более 10 кг) по регионам мира за период от 132 000 лет до 1000 лет
назад
(данные в долях 1). Тёмно-серым цветом выделены регионы, не включённые в анализ (покрытые ледниками в конце плейстоцена или острова)
Реконструкция растительного покрова на
последнем ледниковом максимуме
,
около 18 000 лет назад
, на основе образцов ископаемой пыльцы, извлечённых из озёрных и болотных отложений.
За период от 132 000 лет до 1000 лет
до н.в.
в Африке вымерло 16 % существовавших тогда видов
мегафауны
(18 видов), в Азии 52 % (38 видов), в Европе 59 % (19 видов), в Австралии и Океании 71 % (26 видов), в Северной Америке 74 % (43 вида), в Южной Америке 82 % (62 вида)
. В мире, за исключением Африки, вымерли почти все виды наземных животных с массой более тонны, обитавшие здесь до позднего плейстоцена
. Всего за этот период, на планете вымерли 65 % известных науке видов мегафауны
. Учёные отмечают увеличение количества вымерших видов от Африки к Америкам, связывая это с направлением миграции людей
.
В отличие от Австралии, Южной и Северной Америки, — в
Африке
и на юге Азии род
Homo
, начиная с
Homo erectus
, пересекался с местной фауной 2 миллиона лет, постепенно развиваясь. И животные Африки и южной Азии научились бояться человека, вырабатывая недоверчивость и осторожность
. На север Евразии, в Австралию и на Американский континент попал уже
человек современного типа
, причём относительно недавно, и у многих крупных видов этих регионов не осталось возможности приспособиться к новому типу хищника
.
Данную гипотезу подтверждает то, что на изолированных от людей островах, вымирание животных мегафауны происходило на тысячи лет позже, обычно после появления там людей, что снижает весомость климатической гипотезы. Вымирание почти не затронуло мелкие виды (кроме
симбионтов
мегафауны), а также морских животных
. В этот период не обнаружено и каких-либо катастроф глобального масштаба (вроде падения астероидов), способных повсеместно уничтожить фауну
.
Вымирание животных несло с собой резкие изменения в растительном покрове. Когда вымерли
шерстистый носорог
и
шерстистый мамонт
, вслед за ними изменилась флора —
тундростепи,
на которых они паслись, заросли берёзой и малопродуктивной хвойной
лесотундрой
. Это связано с тем, что стада носорогов и мамонтов поедали молодую поросль берёз, не давая им излишне сильно вырастать, одновременно удобряя почву навозом и способствуя росту трав
.
Стеллерова корова
обитала у Командорских островов в течение 10 000 лет, после полного вымирания вблизи континентов, этот вид был уничтожен людьми всего через 27 лет после открытия. Шерстистые мамонты
острова Врангеля
и
о-ва Святого Павла
пережили материковых мамонтов более чем на 6 000 лет
. Ленивцы вида
мегалокнус
жили на Антильских островах и были уничтожены 4000 лет назад, вскоре после появления человека на островах, при этом все виды
гигантских ленивцев
, жившие на Американском континенте, были уничтожены на 7 000 лет раньше
.
Итоги вымирания
Вымерло процентов от общего числа видов мегафауны по регионам:
В Африке к югу от Сахары 18 (16 % видов)
В Азии 38 (52 %)
В Европе 19 (59 %)
В Австралазии 26 (71 %)
В Северной Америке 43 (74 %)
В Южной Америке 62 (82 %)
Всего за период 132 — 1 тыс. лет назад вымерли 177 известных науке видов наземных млекопитающих массой более 10 кг, в том числе 154 вида массой более 45 кг
.
Основные гипотезы
Животные вымерли из-за изменения климата, связанного с продвижением и отходом крупных ледяных шапок или ледяных покровов, с последующей сменой растительности
.
Животные были уничтожены людьми: «гипотеза доисторической революции» или «доисторического перепромысла»
.
Результаты математического моделирования истории климатических изменений и истории заселения планеты людьми показали, что в 20% случаев вымирания мог преобладать климатический фактор, в 64% случаев большее совпадение с процессом вымирания показали модели, базирующиеся на истории заселения людьми
.
Африка и Азия
Африка
и южная
Азия
относительно не сильно пострадали от четвертичного вымирания, потеряв только 16 процентов фауны и мегафауны в том числе. Это единственные географические регионы, которые сохранили у себя мегафауну, с животными весом больше 1000 кг. На остальных континентах подобная мегафауна была утеряна навсегда.
При этом прослеживается зависимость начала вымирания видов в Африке 2 миллиона лет назад, с появлением там гоминид видов —
Homo habilis
и
Homo erectus
.
В Азии, после появления там
Homo erectus
1,8 млн лет назад. Прослеживается следующая тенденция — с
позднего плейстоцена мегафауна
стала терять виды, которые при этом не замещались другими видами животных того же размера. При природных изменениях климата этого не происходит, освобождающиеся постепенно ниши успевают занимать другие крупные животные. Но в случае возможного антропогенного воздействия, это не происходило, мегафауна не успевала приспособиться к воздействию человека и начать жить по новым условиям
.
Некоторые роды вымерших животных
Мегафауна, исчезнувшая в Африке и Азии во время раннего и среднего плейстоцена
Южный мамонт
- вымерший вид мамонтов, появился в Африке, затем распространился в Евразии и Северной Америке. Предок евразийских и американских видов мамонта, был постепенно замещён в Евразии степным мамонтом около 700 тыс. лет назад
Степной мамонт
— вымерший вид рода
мамонты
, один из крупнейших видов семейства
слоновых
(весом до 10 тонн), живший в среднем плейстоцене в степях и лесах Евразии, до 120 тыс. лет назад. На него охотились неандертальцы, был постепенно замещён шерстистым мамонтом.
Pachycrocuta
—
род
доисторических
гиен
. Pachycrocuta впервые появилась на протяжении позднего
плиоцена
приблизительно три миллиона лет назад, и вымерла в течение среднего
плейстоцена
400 000 лет назад.
Puma pardoides
— вымерший вид кошачьих. Исторически рассматривался как один из древнейших представителей рода
Panthera
.
Стегодоны
— род вымерших
млекопитающих
из отряда
хоботных
. Стегодоны жили во многих частях Азии в
плиоценовую
и плейстоценовую эпохи. Начали исчезать ещё с распространением
Человека прямоходящего
. На отдалённых островах Юго-Восточной Азии (Тимор, Ява, Сулавеси) существовали ещё 130 тыс. лет назад, на территории Южного Китая вымерли между 60 000 и 12 000 лет назад.
Эласмотерии
— род
носорогов
, обитавших в степях
Евразии
с
плиоцена
до конца плейстоцена. Отличались крупными размерами (длина до 6 метров, высота до 2,5 метра, вес до 5
тонн
), вымерли около 30 тыс. лет назад
.
Megalochelys atlas
— род вымерших в плейстоцене исполинских сухопутных черепах весом от 1 до 4 тонн. Все виды жили в Южной Азии, вымерли, вероятно, из-за охоты на них человека прямоходящего
.
Человек флоресский
— гипотетический ископаемый карликовый вид
людей
. Из-за малого роста флоресский человек известен также как «хоббит» (по аналогии с
существами
, придуманными
Дж. Р. Р. Толкином
). Останки
Homo floresiensis
обнаружены в 2003 году в
Индонезии
(остров
Флорес
, пещера Лианг-Буа), где найдены несколько скелетов разной степени сохранности возрастом приблизительно в 13—95 тысяч лет (по последним альтернативным оценкам — 60—100 тысяч лет).
Неандерталец
— вымерший или
ассимилированный
представитель рода
Люди
(
Homo
). Первые люди с чертами протонеандертальца существовали в Европе ещё 350—600 тысяч лет назад, последние неандертальцы жили около 40 тыс. лет назад.
Саблезубые кошки
— вымершее подсемейство
кошачьих
. Появились в раннем или среднем
миоцене
в
Африке
. В Африке вымерли 1,5 млн. лет назад, на юге Азии - 0,5 млн. лет назад, в Европе 30 тыс. лет назад, в Америке около 10 тысяч лет назад. Их вымирание на всех континентах происходило вскоре после расселения там людей
.
Большинство находок подтверждает, что некомпенсированные позднечетвертичные вымирания начались вскоре после прибытия первых людей в Австралию
. В тот период Австралия ещё была
Сахулом
— единым континентом с
Новой Гвинеей
и Тасманией. Вымирания начались 55 тыс. лет назад, в течение 13 тыс. лет наблюдался пик вымираний
. В это время человек проводил экспансию, осваивая новые, до этого незаселённые
гоминидами
территории. На островах происходили схожие процессы, которые продолжались до голоцена —> прибытие людей —> вымирание части фауны. При этом хронология большинства вымираний в Австралии не коррелирует с климатическими изменениями
.
В итоге между 55 и 36 тыс. лет назад
Австралия
и
Океания
потеряли почти всю свою
мегафауну
, около 26 известных науке ископаемых крупных видов
. На сегодняшний день в этих регионах не существует животных весом больше 45 килограммов (кроме пары видов кенгуру в Австралии), которых бы не завозили с других континентов. При этом в предыдущие миллионы лет развития и эволюции, мегафауна этих регионов переживала засухи, ухудшения климата, изменения температур, но не вымирала
. По одним оценкам, 90% вымираний пришлось на период около 46 тыс. лет назад
, по другим, пик вымираний 14 видов происходил около 42 тыс. лет назад
.
Данный факт указывает на то, что причиной вымирания мегафауны стал вероятнее всего человек,
антропогенный фактор
, включая охоту с помощью выжигания растительности
. На территории Австралии археологи нашли свидетельства, что первые люди могли достичь северной оконечности Австралии около 65 тыс. лет назад: найдены отшлифованные каменные топоры, следы перемалывания и обжаривания диких семян. Это говорит о довольно высоком изначальном уровне пришедших людей
.
Некоторые учёные отдают предпочтение климатической гипотезе, считая, что преимущественное влияние человека на некомпенсированные вымирания мегафауны в Австралии не доказано, хотя и не отрицают, что до наших дней за 50 тыс. лет могла дойти лишь ничтожная часть ископаемых свидетельств охоты людей
. Но следует принимать во внимание тот факт, что животные австралийской мегафауны пережили два миллиона лет климатических изменений, а вымирание наиболее крупных видов произошло 50 — 45 тыс. лет назад, вскоре после заселения Австралии первобытными людьми
.
Есть и более экзотические гипотезы. Новозеландские учёные в 2021 году предположили, что по времени пик вымираний в Австралии мог совпасть с резким ослаблением геомагнитного поля Земли примерно 42 - 41 тыс. лет назад (палеомагнитный
экскурс Лашамп
), которое, в свою очередь, могло вызвать сокращение защитного
озонового слоя
атмосферы и повышение уровня жёсткого ультрафиолетового излучения, которое могло привести к
опустыниванию
Австралии
, усугубив воздействие первобытного человека на мегафауну, что в итоге привело к её вымиранию
. Но много крупных видов животных Австралии вымерло и до и после этого события, в период от 48 до 36 тыс. лет назад
.
Дипротодон
— крупнейшее известное
сумчатое
, когда-либо обитавшее на земле, размером с бегемота и весом более 2 т. Дипротодон принадлежит к так называемой
австралийской мегафауне
— группе необычных видов, живших в Австралии в период приблизительно от 1,6 миллионов до 40 тысяч лет назад, то есть бо́льшую часть
плейстоцена
.
Зигоматурусы
— Зигоматурусы были массивными животными с толстыми ногами, по размеру и по телосложению сходными с современными
карликовыми гиппопотамами
. Передвигались на четырёх ногах. Обитали на морском побережье. Вымерли около 45 тыс. лет назад
Diprotodontidae
— семейство очень крупных вымерших сумчатых, эндемичных для
Австралии
. В него включают крупнейших известных науке сумчатых — дипротодонов. Diprotodontidae жили с
олигоцена
по плейстоцен (от 33,9 млн до 40 тыс. лет назад)
Прокоптодоны
— был самым большим из всех кенгуру, когда-либо обитавших на нашей планете. Рост прокоптодона составлял до 3 метров, а вес его палеонтологи оценивают в 232 килограмма.
Проплеопусы
— вымерший род сумчатых. Известны два вида:
P. chillagoensis
, существовавший в плиоцене и плейстоцене, и
P. oscillans
, существовавший в плейстоцене.
Protemnodon
— Животные были подобны
валлаби
, но гораздо крупнее. Самый мелкий из видов, входящих в этот род,
Protemnodon hopei
, весил около 45 килограмм. Представители же других видов могли весить и 110 кг.
Phascolonus
— род вымерших сумчатых
млекопитающих
из семейства
вомбатовых
. Крупнейший вид рода, Phascolonus gigas, весил около 200 кг. Phascolonus появились в
плиоцене
и исчезли в верхнем плейстоцене (5,333 млн — 11,7 тыс. лет назад)
Палорхесты
— род вымерших наземных травоядных сумчатых семейства
Palorchestidae
, размером с лошадь и весом до 1000 кг, похожий на тапира. Эти животные жили в Австралии начиная с позднего
миоцена
до конца плейстоцена и вымерли 50 - 40 тыс. лет назад.
Zaglossus hacketti
— вид вымерших яйцекладущих млекопитающих из семейства
ехидн
, известный из плейстоценовых отложений
Западной Австралии
. Считается крупнейшим из известных науке видов однопроходных. Скорее всего был длиной в метр и весил около 30 кг, как некрупная
овца
, хотя некоторые источники приписывают ему вес до 100 кг.
Вонамби
— род крупных вымерших
змей
, состоящий из двух известных видов. Это были не
питоны
, подобно другим большим констрикторам
Австралии
рода
Morelia
, а члены теперь вымершего семейства Madtsoiidae. Этот род был частью вымершей
мегафауны Австралии
.
Типовой вид
—
Wonambi naracoortensis
, змея 5—6 метров длиной.
Сумчатый лев
— вымерший вид хищных сумчатых из отряда
двурезцовых сумчатых
. Сумчатый лев, ископаемые останки которого находят в отложениях позднего
плейстоцена
, являлся крупнейшим хищным млекопитающим
Австралии
своего времени. По последним данным, связанным с находкой изображения сумчатого льва в пещере, последние сумчатые львы жили одновременно с первобытными
аборигенами Австралии
, то есть 46,000 лет назад.
Сумчатый волк
— вымершее
сумчатое
млекопитающее, единственный доживший до исторической эпохи представитель семейства сумчатых волков.
Мегалания
— крупнейшая из известных науке наземных
ящериц
. Мегалания относилась к семейству
варанов
(Varanidae), которое по принятой в настоящее время классификации включает всего один современный род
Varanus
со множеством подродов и видов. По расчётам самых разных учёных, длина крупных мегаланий варьировалась от 4,5 до 9 м, а масса от 331 до 2200 кг. Этот вид обитал в
Австралии
в
плейстоценовую
эпоху, начиная с 1,6 млн назад и заканчивая примерно 40 000 лет назад.
Квинкана
— вымерший род сухопутных
крокодилов
, представители которого обитали в
Австралии
в
неогене
—
антропогене
. Квинканы появились около 24 млн лет назад, и вымерли около 40 000 лет назад. Во время своего существования квинканы стали одними из главнейших хищников Австралии.
Данное определение вмещает весь Европейский континент и
Кавказ
,
Северную Азию
,
Северный Китай
,
Сибирь
и
Берингию
— нынешние
Берингов пролив
,
Чукотка
,
Камчатка
,
Берингово море
,
Чукотское море
и часть
Аляски
. Во время позднего плейстоцена отмечается большое разнообразие видов животных и семейств, высокая динамика их смешивания, перемещения. Особенностью воздействия оледенений и оттепелей, является высокая скорость с которой они случались — в течение века температуры могли иметь сильный разбег, это обуславливало большие миграции животных в поисках более удобных условий для жизни, что провоцировало генетическое перекрещивание видов.
Последний ледниковый максимум
произошёл между 25 000 и 18 000 лет назад, когда ледник покрыл большую часть
Северной Европы
. Альпийский же ледник покрыл значительную часть Центрально-Южной Европы. В Европе, и в частности в Северной Евразии, температура была ниже, чем в нынешние дни, также климат был более сухим. Огромные пространства покрывала так называемая мамонтовая степь —
тундростепь
. В наши дни похожие климатические условия сохранились в
Хакасии
, на Алтае и в отдельных участках на территории
Забайкалья
и
Прибайкалья.
Данную систему характеризует ивовый кустарник, высокие питательные травы. Биоресурсы тундростепи позволяли поддерживать жизнь и процветать многочисленным млекопитающим, от мамонтов и огромных стад овцебыков и лошадей до грызунов. Низкая высота снежного покрова позволяла травоядным питаться высохшими на корню травами даже продолжительной зимой. Зона включала в себя площадь от
Испании
до
Юкона
в Канаде. По разнообразию пород и их огромной численности, тундростепь почти не уступала африканским саваннам с их огромными стадами антилоп и зебр.
Животные тундростепи включали в себя шерстистого мамонта, шерстистого носорога, степного зубра, предков лошадей, наподобие современных лошадей Пржевальского, овцебыка, оленей, антилоп. Хищники — пещерный медведь, пещерный лев, лиса, серый волк, арктическая лиса, пещерная гиена. Также были тигры, верблюды, лоси, зубры, росомахи, рыси, леопарды,
красные волки
и так далее. При этом численность животных была несравненно выше, выше разнообразие видов, нежели в современный период. В горных частях тундростепи жили архары, снежные барсы, муфлоны, серны.
Во время межледниковий — отступления ледников, область распространения южных животных смещалась на Север. В том числе в Англии 80 000 лет назад жили бегемоты, а в Нидерландах 42 000 лет назад — слоны.
Вымирание произошло в два больших этапа. В первый период, между 50 000 и 30 000 лет назад, вымерли —
прямобивневый лесной слон
,
европейский бегемот
,
,
гомотерии
,
неандертальцы
. Ископаемые кости прямобивнего лесного слона очень часто находятся рядом с кремнёвыми орудиями первобытных людей, которые охотились на них
[
нет в источнике
]
. Второй этап был намного более кратким и скоротечным, между 13 000 и 9 000 лет назад вымерла оставшаяся часть видов мегафауны, включая шерстистого мамонта и шерстистого носорога
.
Кипрский карликовый слон
— считается, что он произошёл от
прямобивневых слонов
. Этот слон населял
Кипр
и некоторые другие средиземноморские острова в
плейстоцене
. По оценкам масса карликового слона была всего 200 кг, что составляет лишь 2 % от массы его предшественников, достигавших 10 тонн.
Cuon alpinus europaeus
— вымерший европейский подвид красного волка. Встречался на большей части Западной и Центральной Европы в течение среднего и позднего
плейстоцена
. Он практически неотличим от современного красного волка, но заметно крупнее. По размерам
Cuon alpinus europaeus
приближался к
серому волку
.
Гомотерии
—
вымерший
род
саблезубых кошек
, обитавших в
Евразии
,
Африке
и
Северной Америке
со среднего
плиоцена
(3—3,5 миллиона лет назад) до конца позднего
плейстоцена
(10 тысяч лет назад). Вымирание гомотериев началось с Африки, откуда эти кошки исчезли примерно 1,5 млн лет назад, в Евразии данный род вымер около 30 тысяч лет назад, и дольше всех просуществовал вид Homotherium serum в Северной Америке — до конца плейстоцена, около 10 тысяч лет назад.
Пещерная гиена
—
вымерший
подвид
современной
пятнистой гиены
(
Crocuta crocuta
), появилась в
Европе
приблизительно 500 000 лет назад и была широко распространена в
плейстоцене
Евразии
, от Северного
Китая
до
Испании
и
Британских островов
. Пещерные гиены начали постепенно исчезать из-за смены природных условий и вытесняемые другими хищниками, а также людьми, примерно 20 000 лет назад, и полностью исчезли из Западной Европы около 14—11 тыс. лет назад, а в некоторых областях и ранее.
Большинство вымираний, после многочисленных проверок и сравнений радиоуглеродных анализов, отнесены к краткому промежутку между 11 500 — 10 000 лет до нашей эры. Этот промежуток длительностью в полторы тысячи лет совпадает с прибытием и развитием на территории северной Америки людей
культуры Кловис
. Меньшая часть вымираний произошла позже и раньше этого временного интервала.
Предыдущие североамериканские вымирания произошли в конце оледенения, но не с таким перекосом в сторону крупных животных. Также важно отметить — прошлые вымирания, имевшие явно природные причины, не антропогенные, не были массовыми, а наоборот были постепенными. Родственники слонов —
мастодонты
, вымершие в Азии и Африке ещё 3 млн лет назад, в Америке дожили до прихода современных людей. При этом биологические ниши от вымерших животных благодаря плавности вымираний успевали быть занятыми другими видами, приспособившимися к новым условиям.
Как и в
Евразии
, при антропогенном воздействии в Северной Америке вымирания происходили массово, во многом хаотично, очень быстро по меркам природы и биологические ниши оставались незанятыми, что провоцировало дальнейший дисбаланс в фауне и флоре
.
Первые, точно датированные поселения людей на
Аляске
, севере Северной Америки, появились 22 000 лет назад, куда человек перебрался из Азии по
Берингии
. После отступления ледников на Аляске 16 000 лет назад, люди очень быстро, в течение 1-2 тыс. лет смогли заселить всю остальную Северную и Южную Америки
, увеличив численность населения в 60 раз за этот период
.
Интересен
вилорог
— единственный представитель рода вилороговых на сегодняшний день, самое быстрое наземное животное после гепарда. Как предполагается, именно высокая скорость передвижения сделала его сложной добычей и он смог дожить до наших дней.
При этом есть животное, которое, на первый взгляд, не вписалось в концепцию антропогенного истребления видов. Это
бизон
. Данный вид появился не в Северной Америке, он мигрировал через Берингию и в течение последующих 200 000 лет был отделён от охотников людей ледниками. Сообразно представлениям этологов, животные за 200 000 лет должны были стать такими же наивными, как и фауна Австралии, но, видимо, этого не произошло из-за наличия крупных и быстрых хищников (медведи, пумы, волки), и бизоны сохранили осторожность либо оказались слишком быстрыми и опасными для первобытного человека, подобно кафрским буйволам, и потому не были истреблены. У индейцев до прибытия европейцев не было лошадей, необходимых для преследования бизонов. Известны случаи, когда стада бизонов затаптывали людей, не имевших лошадей и огнестрельного оружия. Овцебыки, не пытающиеся бежать при приближении человека, сохранились в небольшом числе только на нескольких труднодоступных приполярных островах Северной Америки и были открыты европейцами лишь в конце XVII века.
Культура людей, связанная с самой мощной волной вымирания —
Кловис
, имеет
древне-индейское
происхождение. Они охотились на крупных
хоботных
(мамонтов, мастодонтов, гомфотериевых
), используя метательные копья, для увеличения силы броска применяли
атлатль
. Из-за доверчивости крупных травоядных, не имевших природных врагов и не видящих в людях опасности, охота на этих животных не представляла сложности для человека. «
Можно было подходить и вплотную колоть этих зверей, и они даже не понимали, что происходит
» — пишет антрополог
Станислав Дробышевский
. Исследователи не отрицают возможного смешивания двух факторов, способствовавших вымиранию — окончание ледникового периода 14 - 12 тыс. лет назад с резким изменением климата и снижением продуктивности кормовой базы, и вкупе с ним резко возросшая охота людей культуры Кловис, которые вынуждены были ориентироваться именно на добывание животной пищи. При этом пик вымирания мегафауны совпал с расцветом этой культуры, так что люди могли быть причастны к исчезновению мегафауны
. Известны как минимум 12 «мест забоя и разделки
хоботных
» людьми Кловис, что является очень большим числом для такой кратковременной культуры
. В итоге всё это могло сложиться в крайне неблагоприятную формулу и случилось резкое сокращение видового разнообразия на континенте
.
Южная Америка
Life restoration of the Yukon wild horse,
.
В силу длительной, в течение нескольких миллионов лет, полной изоляции, данный континент не имел широкого набора представителей фауны, если сравнивать с Евразией или Северной Америкой. Между двумя Америками произошло интересное событие —
Великий межамериканский обмен
— 3 миллиона лет назад участки морского дна поднялись и образовали современный панамский перешеек. Это спровоцировало первое, подтверждаемое раскопками, большое вымирание в Южной Америке, когда виды из Северной Америки стали мигрировать на новый континент. До этого события, Южная Америка обладала уникальной фауной — почти все животные были эндемиками, жившими только на этом континенте.
В результате первичного вымирания, именно природного, неотропические виды оказались заметно менее успешными, нежели виды, пришедшие из Северной Америки, за исключением нескольких видов гигантских ленивцев, переселившихся из Южной в Северную Америку.
В плейстоцене Южную Америку практически не затронуло оледенение, за исключением Андских гор. К началу голоцена, 11 000 - 9 000 лет назад, через 2 - 3 тыс. лет после начала заселения людьми, 82% видов мегафауны вымерли
. В этот период вымерли
кювьерониусы
и
стегомастодоны
(родственники слонов)
, гигантские броненосцы весом до 2 тонн —
дедикурусы
и
глиптодоны
,
гигантские ленивцы
, достигающие 4 тонн веса,
южноамериканские копытные
—
макраухении
и
токсодоны
размером с носорога,
гиппидионы
. Меньшие по размерам броненосцы дожили до наших дней. Нишу сумчатых заняли опоссумы. Последние гигантские ленивцы на островах Куба и Гаити просуществовали до 2 тысячелетия до нашей эры, исчезнув вскоре после появления людей на этих островах.
При этом, гигантские ленивцы, стегомастодоны и кювьерониусы были пластичны в питании, обитали в разных климатических зонах, от пустынь до тропических джунглей, так что трудно объяснить их вымирание изменениями климата
.
На сегодняшний день крупнейшими наземными млекопитающими Южной Америки являются разновидности верблюдов —
гуанако
и
викунья
, а также
центральноамериканский тапир
— достигающий веса в 300 кг. Другие сохранившиеся, относительно крупные представители фауны прошлого это, —
пекари
, пумы, ягуары, гигантские муравьеды, кайманы, капибары, анаконды.
Изменение численности мегафауны на различных участках суши планеты с указанием времени прибытия на них людей
Пока что нет общей теории, которая бы разграничивала
голоценовое
и позднечетвертичное вымирания, а также вымирание из-за антропогенных и климатических факторов. По одной из точек зрения, изменения климата и человеческий фактор должны быть связаны воедино, другие учёные отстаивают теорию о том, что нужно разделять эти причины в отдельные эпизоды, либо отдают приоритет одной из причин.
При этом часть учёных связывает вымирание крупных животных в
Африке
, Евразии, с тем что 200—100 тысяч лет назад люди современного типа стали резко расти в численности, научились охотиться при помощи камней, метательных копий и так далее и тем самым резко повысили свою эффективность, как охотников и при этом свою способность в уничтожении животных. Для изолированных от гоминид островов
Новой Зеландии
и на
Мадагаскаре
, фауны Южной Америки, Австралии и Северной Америки, было достаточно даже относительно среднего воздействия новых хищников, чтобы начать терять разнообразие видов крупных животных. Воздействие человека на природу в процессе развития только усиливается, в последующем антропогенный фактор стал причиной исчезновения растений, загрязнения и окисления выбросами воздуха и океана.
Гипотеза охоты и разрушения людьми среды обитания животных
Данная гипотеза увязывает охоту человека на крупных
млекопитающих
с тем, что после их выбивания и исчезновения из фауны, вслед за ними вымирали хищники, которые специализировались на охоте на крупных животных. В пользу такого взгляда говорят находки, где на костях животных найдены характерные повреждения от стрел, копий, следы обработки и разделки туш, при которых наносились повреждения на кости. В европейских пещерах найдено много изображений, где изображена именно охота на крупную добычу. Медленный рост, долгий период созревания крупных животных, их высокая пищевая ценность (больше мяса в единицу времени), хорошая заметность повышали уязвимость крупных животных перед первобытными охотниками
.
Также прослеживается зависимость в сохранении фауны и в начале экспансии человека. В
Африке
и южной Азии животные, находясь рядом с предками человека несколько миллионов лет, постепенно смогли научиться бояться людей. Люди не сразу стали умелыми охотниками и допускали ошибки, у них поначалу не было оружия, тактики и умений, которые они вырабатывали постепенно. В итоге африканская фауна и в особенности крупные животные, хоть и пострадали, потеряв немало родов и видов, но сумели приспособиться, научились либо убегать, либо прятаться, либо атаковать и отбивать атаки людей
. Дожившие до наших дней слоны, носороги и бегемоты, несмотря на свой вес, могут развивать скорость бега больше чем у человека, при этом по пересечённой местности, хотя естественных врагов в природе у них почти нет.
Так, наиболее опасными животными в итоге стали слоны, львы, бегемоты и носороги. На сегодняшний день в Африке самым опасным животным, по статистике убийств, являются именно бегемоты, которые при всей своей кажущейся неповоротливости очень активно защищают себя, свою территорию и тем более потомство. Это увязывается с тем, что бегемоты явно являлись лакомой добычей для людей — они имеют огромный вес и на вид относительно безобидны. Длительная эволюция, с постепенно развивающимися людьми, сделала бегемотов и носорогов грозными противниками, места обитания которых люди впоследствии стали избегать. Если присмотреться к копытным, то они также умеют за себя постоять и делают это активно — зебры умеют драться всеми ногами и зубами. Антилопы вступают в противоборство даже с прайдами львов, что не единожды записывалось на видео исследователями, вплоть до того, что антилопы сбиваются в ударные группы самцов и нападают на прайды, во главе которых стоят крупные самцы львов. Такое поведение говорит о том, что даже травоядные в Африке приучены защищать себя активно.
Кроме того, тропическая Африка — место распространения множества опасных болезней и паразитов, ещё недавно смертельных для человека и домашнего скота:
трипаносом
(«
сонная болезнь
»), переносимых мухой
цеце
, малярии, различных тропических лихорадок,
африканской чумы свиней
и т. д. Дикие животные Африки за миллионы лет выработали к ним иммунитет, а люди и домашний скот — нет. Все это, до недавнего времени, препятствовало освоению Тропической Африки под пастбища и посевы и спасало районы обитания крупных животных от людей
.
Первичным и самым простым способом охоты группой было отнимание уже убитой добычи у крупных хищников. Подтверждено многочисленными наблюдениями зоологов — ряд хищников очень легко бросает даже только что убитую добычу, если его окружают стервятники или мелкие хищники. Древние люди использовали схожую тактику — они окружали хищника, кричали, забрасывали камнями, пугали палками и копьями. Хищник пугался и оставлял свежую добычу. Данный подход, возможно, способствовал вымиранию ряда родов кошачьих, в том числе и крупных.
Впоследствии люди освоили именно охоту группой, когда часть людей отвлекает крупного зверя, а другие стараются ранить ему ноги и живот. Охота на слонов и мамонтов также привела к появлению оригинальных способов. К примеру, люди стали делать небольшие ямы-ловушки, как раз чтобы в яму немного провалилась нога слона или мамонта. На дне ямы были установлены колья — они ранили стопу животного. Слон в силу большого веса и габаритов не способен долго стоять и передвигаться на трёх ногах, и в течение пары часов он падал. Дальше люди убивали добычу. Такой способ позволяет не тратить много сил на преследование добычи и не рисковать жизнью, подкрадываясь к опасному животному из засады. Люди освоили «дальнобойное» оружие — метательные копья,
копьеметалки
. Однако это же и способствовало быстрому истреблению многих
хоботных
, включая мамонта и ряд других.
При этом на других континентах, в особенности на тех, куда человек пришёл позже, животные, в том числе и крупные, были доверчивы, наивны, они не видели опасности в существах, намного меньших их по размеру
. Люди пришли в ту же Австралию, Северную и Южную Америку, север Евразии уже значительно более умелыми. Они были вооружены луками, копьями, пращами, умели работать в команде, атакуя животных разом
.
Мамонты
,
мастодонты
и
гомфотериевые
,
гигантские ленивцы
были истреблены в Америке всего через 2—3 тыс. лет после появления там людей 15 000 лет назад
, так как были незнакомы с человеком, не умели или не могли ему противостоять
. Все эти животные сотни тысяч лет жили в разных климатических зонах, но вымерли почти одновременно с расселением человека
. Человек пришёл в Австралию владея огнём и мог устраивать подпалы — поджигать сухую траву
. Такая подготовка в итоге трагически сказалась на фауне — островная фауна оказалась в особенности уязвимой — наиболее показателен пример нелетающих и медлительных
дронтов
,
моа
или
эпиорнисов
, которые вообще не были способны защитить себя от крупного хищника, включая человека, в отличие от тех же цапель в Африке.
Австралийские племена с таким подходом выжгли траву и растительность почти на всём континенте. Охота путём загона животных при помощи пожара, несла в себе колоссальный урон биосфере и стала одной из базовых причин вымирания уникальной фауны и флоры континента
.
При этом корреляция между прибытием людей и вымиранием
мегафауны
практически прямая, без поправок.
Шерстистый мамонт
выжил на недоступных для людей островах
Врангеля
и
Прибылова
вплоть до 1700 года до нашей эры (спустя 5000 лет после вымирания на материке), при этом изменения климата (окончание оледенения и повышение температуры) не спровоцировали его вымирание в течение тысяч лет. Гигантские ленивцы
мегалокнусы
жили на
о. Куба
и
Гаити
ещё 2 000 лет до нашей эры, спустя 7 000 лет после вымирания на Американском континенте, но вымерли вскоре после появления первых людей на этих островах
.
Волна тотальных вымираний в Австралии 50 000 лет назад, не увязывается с климатом — резких изменений не было, но имеет прямую связь с прибытием людей на континент
.
Исследования от 2017—2018 года, в журнале
Science
, подтверждает прямую зависимость, между прибытием на тот или иной континент людей рода
Хомо Сапиенс
и последующим резким вымиранием мегафауны. Выявлено, что в течение
кайнозойской эры
, вымирания шли плавно и глобально, вымирали в равной степени как крупные, так и мелкие виды животных. 29 миллионов лет назад случился кризис в вымирании именно мелких существ, в связи с сокращением площадей лесов и увеличением доли саванн и степей.
Принципиально иная ситуация сложилась в ходе Четвертичного периода (
антропоген
) и в частности во время Четвертичного вымирания. В промежуток между 130—15 тысяч лет назад, в
поздний плейстоцен
, вымирание животных обрело направленность в сторону крупных видов
. Подобная тенденция сохранилась до наших дней — наиболее активно истребляются и затем вымирают, именно представители мегафауны. Те животные, что обладают меньшим весом, не так уязвимы и не представляют столь удобной добычи, быстрее размножаются и адаптируются к преследованию со стороны человека, а также к изменению внешних условий. Например, у
слоновых
, к которым относятся мамонты, половое созревание наступает в возрасте 10 — 15 лет
, в неблагоприятных условиях ещё позже, в 17 — 20 лет, тогда как лоси начинают размножаться уже в возрасте 2 года, что делало популяции мамонтов ещё уязвимее при интенсивной охоте в неблагоприятных климатических условиях. В суровых условиях Арктики, у первобытного человека не было такого выбора пищевых объектов, как у людей, живущих в тропических районах, где вегетация круглогодичная, поэтому человек в Арктике, чтобы выжить, вынужден был охотиться на любую добычу, особенно на такую крупную, как мамонты. При этом в
голоцене
избирательность несколько сгладилась, стали вымирать и мелкие животные, но это объясняется всё более возраставшим
антропогенным воздействием
, при котором стала резко сокращаться свободная от людей зона обитания диких животных, площади лесов, природных степей.
«Почему ты охотишься только на медведя? — спросил его как-то Клош-Кван. И Киш сумел дать ему надлежащий ответ:
— Кто же не знает, что только на медведе так много мяса.»
Данные факты указывают, что ситуация с вымиранием животных в Четвертичном периоде, является уникальной для всей кайнозойской эры и не имеет аналогов в плане избирательности, когда больше всего пострадали именно крупные млекопитающие —
мегафауна
. Столь узкого перекоса в сторону вымирания мегафауны, не наблюдалось и в другие периоды, когда были
массовые вымирания
.
Также подтверждено, что резкое изменение климата не способно избирательно привести к вымиранию именно мегафауны
.
В итоге учёные находят всё больше подтверждений, что превращение человека рода Хомо сапиенс в своего рода
супер хищника
, умевшего охотиться разными способами, обладавшего к тому же развитым интеллектом, является главной причиной вымирания крупных животных в Четвертичный период. Из-за этого охотничьего статуса и умений человека разумного, за последние 125 000 лет фауна резко измельчала. Причём динамика исчезновения крупных видов по континентам, почти точно отражает расселение людей рода Хомо на эти континенты.
Австралия
— резкое вымирание мегафауны между 55 − 40 тысячами лет назад — первые люди пришли на континент 60 тысяч лет назад.
Север
Евразии
— 25 − 15 тыс. лет назад, когда потепление климата и отступление ледников позволили людям заселить ранее недоступные районы.
При этом
Южная
и
Северная
Америки, в период этих вымираний, были по сути заповедниками, где животный мир не сокращал резко своё видовое разнообразие, включая крупных животных. Этот факт напрямую связывается с тем фактом, что люди на эти континенты ещё не мигрировали. Но между 15 — 11 тысячами лет назад, на этих континентах также произошло резкое вымирание
мегафауны
, напрямую
коррелирующее
с прибытием на эти континенты людей. Люди смогли перебраться в Северную Америку по
Берингии
и расселиться там 15 000 лет назад.
Компьютерное моделирование, проведённое в 2015 году по моделям Мосманна и Мартина
и Уиттингтона и Дайка
, подтвердило эти выводы. Были заложены данные о климате на всех континентах за последние 90 000 лет, время вымирание видов и прихода людей на разные континенты. Время вымирания животных совпадало с приходом людей в обеих моделях. При этом климат не становился причиной вымираний, но при активном антропогенном воздействии, усугублял вымирание животных. Также было отмечено, что вымирание имело относительно низкую скорость в Азии, по сравнению с Австралией, островами и Северной и Южной Америками. Данный факт увязывается с тем, что сначала люди попали именно в Азию и там они ещё были относительно не развиты, по сравнению с тем моментом, когда мигрировали на другие континенты, и животные, частично, но успевали приспособится к новому типу хищника
.
Выводы и возражения против гипотезы неумеренной охоты
Люди и мамонты в южной Сибири сосуществовали бок о бок более 12 000 лет, с 32 000 до 20 000 лет назад, до начала резких колебаний климата, сокративших площадь растительности, пригодной для обитания мамонтов. Люди, в этом случае, возможно были вторичной причиной исчезновения, добив сократившиеся популяции мамонтов и не дав им возможности восстановиться
.
Хищники в природе не могут излишне интенсивно охотиться на тот или иной тип добычи, так как энергетические затраты на погоню за добычей, ставшей редкой, рано или поздно перестанут окупаться её пищевой ценностью. Хищник начнёт голодать, не сможет больше преследовать жертву и давать отпор конкурентам. В первую очередь, человек всегда охотился за наиболее доступной добычей, имеющей наиболее высокую пищевую ценность — на крупных, медлительных травоядных животных, которых легче было преследовать и которые при этом давали много мяса: мамонтов, мастодонтов, гигантских ленивцев,
гигантских броненосцев
, гигантских сумчатых. Ранее у таких животных почти не было врагов в природе из-за их размеров и силы, опасности в ближней схватке. На таких животных человек мог нападать за 10-15 м, забрасывая их копьями из-за пределов досягаемости их когтей, копыт и зубов, либо устраивая им ловушки
. Поэтому такие животные вымерли в первую очередь
. Но у людей в тропиках был большой выбор альтернативных продуктов, включая полностью растительную диету, если та или иная дичь становилась редкой, что позволило некоторым крупным видам животных сохраниться в Африке и на юге Азии. Из-за эпидемий тропических болезней, кровососущих насекомых (переносчиков инфекций и паразитов), многие районы джунглей и саванн Азии и Африки были труднодоступны и опасны для человека и домашнего скота. Поэтому, до недавнего времени, большинству видов диких животных удавалось там сохранить жизнеспособные популяции, даже при воздействии человека
. На выживание азиатских слонов повлияло то, что они были частично одомашнены. Кроме того, распространение буддизма и индуизма, в которых слоны и некоторые крупные копытные считаются священными животными, позволило им сохраниться в горных районах Южной и Юго-Восточной Азии, малопригодных для сельского хозяйства. Тогда как на Ближнем Востоке (
Месопотамия
, Сирия, Ирак, Малая Азия, Пакистан), на большей части Китая, в Северной Африке слоны были истреблены людьми, вытеснены сельхозугодьями, и, частично,
аридизацией
климата уже к началу нашей эры
.
Часть животных в Северной Америке не вымерла, включая бизонов. При этом данный вид находился в полной изоляции от человека на протяжении 240 тысяч лет и растерял былую осторожность по отношению к людям, но не стал таким же наивным как обитатели фауны Австралии, так как в Северной Америке оставались крупные и быстрые хищники — волки, пумы, медведи гризли. Белые переселенцы в Америку застали огромные стада бизонов. До появления у индейцев прерий лошадей и огнестрельного оружия, завезённых европейцами, они не могли эффективно преследовать бизонов, которые были достаточно быстрыми и опасными для пешего охотника стадными животными. Индейцы, до прибытия европейцев, не имели домашний скот (кроме
ламы
в Андах), вытеснявший стада диких копытных.
Рождаемость охотничьих популяций людей была очень высокая, так как контрацепции не было в принципе
. Но такой же высокой была и естественная смертность в прошлом (от болезней, голода, межплеменных войн, травм и увечий) — люди жили в среднем не больше 30 лет. У первобытных народов (
огнеземельцы
, индейцы) в частые периоды голода практиковались
геронтицид
и
инфантицид
. При этом охота на того же мамонта давала огромное количество мяса и жира и его физически было бы слишком много, чтобы была необходимость продолжать охоту, — пока мамонты не были полностью истреблены. Это заставило людей голодать и искать более стабильные источники пропитания, заботиться о сохранности своих охотничьих ресурсов.
Стоит учитывать колоссальную разность
менталитетов
охотников прошлого и современных техногенных сообществ. Охотники, те же индейцы племени лакота, чукчи, ненцы, якуты, никогда не убивали добычи больше, чем нужно для пропитания и для необходимых запасов мяса, защищали свои охотничьи угодья от посягательств других племён. Индейцы лакота убивали строго определённое количество бизонов, при этом в использование шла обязательно вся туша, без остатков, чем не может похвастать современная технологическая культура, которая оставляет много отходов. Лакота имели доступ к миллионным стадам бизонов, но никогда не брали больше, чем необходимо. Чукчи в Чукотском крае также строго придерживались принципа — только необходимое количество мяса. убивается всегда ровно столько китов, чтобы прокормить всех и сделать запасы в
ледниках
, но не больше.
. Однако, современные учёные возражают на это, что у индейцев, до прибытия европейцев, просто не было лошадей и огнестрельного оружия для массового истребления бизонов, а лодки чукчей и эскимосов не позволяли им преследовать китов вдали от берега
.
В межплеменных войнах, от болезней и голода погибало
избыточное население
первобытных охотников, если природная среда
не могла прокормить
всех. Поколения охотников знали охотничью ёмкость своих угодий, но в случае голода всякие запреты на охоту забывались ради выживания. Редкость крупных животных не была для человека запретом их добычи. Напротив, они становились наиболее желанной и ценной добычей, так как представляли собой огромную и хорошо заметную гору мяса и жира, и человек целенаправленно выслеживал и преследовал мегафауну, не позволяя крупным животным восстанавливать свою численность
.
Европейские переселенцы в США, с огнестрельным оружием выбивали тысячи бизонов просто ради развлечения, либо для подрыва продовольственной базы индейцев, полностью уничтожив миллионные стада бизонов, миллиарды
странствующих голубей
и прочих массовых видов за каких-то 50 лет
. При этом каких-либо археологических свидетельств массового истребления бизонов 150 лет назад, сейчас почти не сохранилось, что отвергает весомость аргументов «климатистов» о редкости свидетельств охоты первобытных людей на мегафауну, вроде застрявших в костях мамонтов каменных наконечников копий. За прошедшие десятки тысячи лет до нас могла дойти лишь ничтожная часть таких артефактов
.
Гипотеза об изменении климата
Изменения температуры за последние 5 миллионов лет
[
нет в источнике
]
Уже в конце 19 века и начале 20-го, учёные заметили цикличность оледенений, а также то, как изменялась фауна, вымирали виды и их ниши занимали новые животные. Это навело на мысль о взаимосвязи климата и состава фауны и флоры.
Однако критики возражают на это тем, что оледенений и потеплений было очень много, но при этом фауна никогда столь резко не сокращалась и при этом успевала заместить вымерших животных новыми видами. Именно в период между 20 — 9 тысячами лет назад, произошёл огромный мегафаунный провал, вымерло очень много родов крупных животных, и это совпадает с всплеском численности людских сообществ и, в том числе, с появлением современного типа человека — кроманьонца, который был таким же умным, как и современные люди, и был способен организовывать охоту на любых животных, которых хотел добыть.
Анализ бивней мастодонтов в районе Великих озёр, говорит о том, что в течение нескольких тысяч лет до исчезновения, мастодонты умирали более старыми и оставляли всё меньше потомства. Это плохо согласуется с климатическими изменениями, которые должны были сокращать время жизни, но логично, если предположить, что охотившиеся люди столетие за столетием сокращали поголовье мастодонтов и у оставшихся уменьшалась внутривидовая конкуренция, они не рисковали в стычках с соперниками за самок и за пастбища.
. Охотники
Кловис
выбивали в первую очередь одиноких молодых самцов мастодонта и мамонта, изгоняемых из семейного стада по достижении половой зрелости, как это принято у слоновых (на одиночных животных проще и безопаснее охотиться), либо детёнышей, сокращая тем самым генофонд и возможность размножения этих животных
.
Повышение температуры
Наиболее явным последствием окончания очередного оледенения является повышение температуры. Между 15 000 и 11 000 лет назад наблюдалось повышение среднегодовой общепланетной температуры на 10-12 градусов Цельсия
. По данной теории, такое потепление создавало плохие условия для тех животных, которые приспособились жить в холодном климате, за счёт изменения растительности, которые питались травоядные животные мегафауны. Из-за таяния ледникового щита, уровень мирового океана поднялся на десятки метров, затопив прибрежные низменности. Выросла влажность и высота снежного покрова зимой в северных районах, что привело к исчезновению
тундростепей
и затрудняло крупным травоядным добывание пищи из-под снега, южные районы тундростепей заросли хвойной тайгой, а южные степи (
прерии
) стали более сухими в летний период, из-за усиления континентальности климата.
Согласно данным ДНК и археологическим изысканиям, температура явно оказывала влияние на видообразование, на вымирание тех или иных животных и растений и замену их другими. При этом человек мог послужить тем фактором, который мешал естественному видо-замещению, выбивая те популяции крупных животных, которые могли бы заменить уже вымершие или вымирающие, охотился на путях миграций крупных животных, тем самым ещё больше усугубляя вымирание.
Изменения растительности: географические
Возможно, что резкие изменения климата и смена растительности повлияли на виды и многие животные не смогли приспособится. Укоротившиеся сезоны роста трав могли вызывать разное влияние на разных млекопитающих. Некоторые авторы считают, что бизоны и другие жвачные чувствовали себя лучше, нежели лошади и слоны. У зубров и им подобных лучше развита способность к перевариванию жёсткой, трудноусваиваемой клетчатки и способность противостоять токсинам в травах. В итоге, те животные, которые были излишне специализированы на одном типе корма, становились намного уязвимей при смене растительного покрова. К примеру, наиболее известный подобный вид —
большая панда
— поедает определённые виды бамбука, как основу растительной диеты и незначительное количество животной пищи. Но именно бамбук и его побеги служат главной пищей для панд, и в случае гибели побегов бамбука, панды гибнут от голода. При этом
корова
являет собой пример высокой степени приспособленности к любым растительным диетам, включая и сочные, мягкие травы и побеги кустарников и молодые деревца и жёсткие травы, сухие по своей структуре.
. Однако, при сравнении рациона овцебыков и северных оленей выяснилось, что овцебыки более приспособлены к питанию различными травами и кормами, чем северные олени, так что в вымирании овцебыков главную роль сыграла охота людей
. Слоны в Африке могут существовать в пустынях Намибии и Южной Африки, в тропических джунглях Конго, болотах
Окаванго
, поедая любую растительность, от колючек и сухой травы до ветвей деревьев, кочуя, при необходимости, на десятки километров
.
Изменения уровня осадков
Усилившаяся континентальность климата привела к менее предсказуемому количеству осадков. Это стало прямым образом влиять на флору — травы и деревья, а значит на кормовые базы. Колебания уровня осадков ограничили периоды, благоприятные для размножения и питания. Для крупных животных подобная смена циклов может оказаться фатальной, при сочетании прочих неблагоприятных факторов. Учитывая, что возраст полового созревания и сроки беременности у таких животных намного выше, мелкие животные опять таки находятся в выгодном положении — у них гибче периоды спаривания, короче сроки полового созревания и беременности, значит им легче размножаться, быстро и эффективно восстанавливая свою популяцию. Поэтому, в условиях неблагоприятного изменения климата, при усилении пресса охотников, более всего страдают виды крупных животных.
Исследования 2017 года по экологии в Европе, Сибири и Америке между 25 000 и 10 000 лет назад, показало, что длительное потепление, приведшее к оттаиванию ледников и повышению уровня осадков, произошло непосредственно перед трансформацией пастбищ. До этого пастбища стабилизировались в плане осадков водно-болотными площадями, обеспечивавшими относительное постоянство кормовых угодий. Из-за роста влажности и уровня CO
2
в атмосфере, выросла высота снежного покрова зимой в северных районах, что привело к исчезновению
тундростепей
, затрудняя добывание пищи крупным травоядным (мамонтам, шерстистым носорогам) из-под снега в достаточном количестве
.
Когда баланс осадков сменился, старые кормовые угодья исчезли и под ударом оказалась мегафауна. Однако транс-экваториальная позиция Африки позволила сохранить пастбищные угодья между пустынями и центральными лесами и поэтому в Африке мегафауна относительно несильно пострадала от подвижек климата
.
Аргументы против гипотезы потепления климата
Противники теории о повышенной температуре, как причине вымирания, указывают, что оледенения и последующие потепления — это циклический, глобальный процесс, который происходит на земле уже сотни тысяч и миллионы лет. При этом многие крупные животные прекрасно адаптировались к циклам похолоданий-потеплений. Поэтому одного только повышения температуры недостаточно для столь массовых и избирательных вымираний
.
Так, мамонты выживали долгое время на острове Врангеля и
о-ве Святого Павла
(Аляска), спустя 5000 лет после потепления, благодаря отсутствию людей на этих островах. Известно, что именно малые популяции наиболее склонны к вымиранию из-за любых изменений. Но с мамонтами этого не произошло на фоне колебаний температуры
.
Потепление климата, отступление ледников, развитие методов охоты способствовало росту населения и распространению людей — охотников в ранее незаселённые районы Арктики 30 000 −15 000 лет назад
. Мамонты, до расселения современного человека, 50 − 70 тыс. лет назад обитали не только в тундростепи, но и в межгляциальной лесостепи Испании и Англии, в смешанных лесах Китая. Шерстистый носорог населял сухие степи Монголии. Болота, гнус, высокий снежный покров не мешали мамонтам, как и современным
лесным бизонам
в тайге Канады. Расселение людей и пресс охоты вынуждали мамонтов и шерстистых носорогов отступать всё дальше на север. В периоды
дриасовых
похолоданий климата уже не происходило, как раньше, увеличения их численности и восстановления ареала, так как этому мешал человек
.
Гипотеза о влиянии климата на вымирание частично подтверждается только для севера Евразии, и даже тут оспаривается
. Для других регионов мира вымирание не коррелирует с колебаниями климата, но напрямую совпадает с расселением людей. Больше всего пострадали регионы и континенты, которые человек заселил относительно недавно
.
Вымершие животные должны были наоборот, начать процветать. В частности, для травоядных стало больше травы. Для мамонтов и лошадей, прерии по всем выводам должны были стать не менее удобными, чем прошлые ландшафты
.
Разные виды
мамонтов
,
американские мастодонты
,
гомфотериевые
,
токсодоны
,
гигантские ленивцы
, гигантские броненосцы -
глиптодоны
жили в совершенно разных климатических зонах Северной и Южной Америки (в тундре, степи, в лесах умеренного пояса, тропических джунглях
), но все они вымерли вскоре после расселения человека на Американском континенте 15 - 12 тыс. лет. назад
. При этом, на такой огромной территории, как Американский континент, джунгли, леса, степи, тундра не исчезали в этот период, несмотря на все климатические изменения и сохранились до наших дней, а мегафауна исчезла
.
Западная лошадь
вымерла в Северной Америке 11 тыс. лет назад, но когда лошади были восстановлены в дикой природе в 16 веке в виде одичавших домашних европейских (
мустанги
), они не начали снова вымирать. Наоборот — обучились находить корм в любой период года. При этом лошади адаптировались к тем травам, которые содержат токсины, сроки беременности не мешают лошадям размножаться, несмотря на периоды засух и низкого количества и качества трав.
Обычно крупные млекопитающие успешно мигрируют в поисках пастбищ, что наглядно демонстрируется в современной Африке огромными миграциями антилоп и слонов. Потепление климата происходило не мгновенно, а в течение сотен и тысяч лет, что позволяло крупным животным откочёвывать в пригодные для них климатические зоны. Трансэкваториальная позиция Американского континента позволяла это сделать, но из-за расселения человека по всей Америке 15 − 12 тыс. лет назад, у мегафауны Америки уже не было возможности приспособиться к новому глобальному суперхищнику, либо мигрировать и она почти полностью вымерла.
По результатам раскопок известно, что гигантские ленивцы, стегомастодоны и кювьерониусы в Америке были очень пластичны в питании, могли обитать в разных климатических зонах, так что трудно объяснить их вымирание изменениями климата. Изменения климата в тропиках Америки не слишком отличались от изменений климата в тропиках Африки, на той же широте и в тот же период 15 - 11 тыс. лет назад, но степень вымирания мегафауны резко отличалась на этих континентах
.
Большие животные имеют бо́льшие жировые запасы, это должно было помогать им переживать засухи, морозы и сложные периоды
.
На Аляске в данный период очень низко-питательные почвы. На этом сложено предположение, что истребление человеком мегафауны привело к деградации северных ландшафтов и постепенному зарастанию
мамонтовой степи
хвойной тайгой и лесотундрой, а не изменение климата
. Как показывает история наблюдений за слонами в национальных парках Африки, слоны активно препятствуют зарастанию саванн кустарником, поедая их поросль, ломая и объедая даже достаточно крупные деревья
.
В Австралии вымирание мегафауны началось 55 − 45 тыс. лет назад, задолго до изменения климата в конце плейстоцена, но после появления там людей
.
Циклические изменения климата, периодически приводящие к уменьшению численности крупных травоядных, заставляли голодающих людей ещё больше усиливать пресс охоты на оставшихся крупных животных, изобретать новые, более изощрённые методы их добычи, не позволяя мегафауне восстанавливать свою численность
. В результате, преимущество получили быстро размножающиеся мелкие животные, которых человеку труднее было выслеживать и преследовать.
Теория заболеваний, эпидемий
Основана на предположении, что следующие за людьми животные — домашние собаки — являлись переносчиками сильно заразных, вирулентных заболеваний. Для млекопитающих, не имевших иммунитета к нему, такое заболевание становилось фатальным. Схожий процесс имел место в историческую эпоху — на Гавайях от завезённых людьми заболеваний пострадали популяции диких птиц.
Но для подобного уровня, при котором вымирание постигло огромное количество животных, включая крупных, на колоссальных площадях, размером почти с Евразию, заболевание должно удовлетворять многим факторам. Во первых она должна иметь постоянный природный очаг где бы болезнь сохранялась, даже если в других местах нет новых заражённых животных. Во вторых заражаемость болезнью должна быть полной — все возрасты и размеры, самцы и самки. В третьих смертность должна превышать 50 — 75 процентов. В четвёртых — болезнь должна уметь заражать несколько видов животных, при этом не являясь смертельной для людей.
Смилодон в сравнении с человеком.
Лев — обитал в Америке.
Однако, если предположить, что заболевания передавались с домашними собаками, то вымирание видов в Австралии и Океании не подпадает под такое объяснение. Собаки появились в этих местах лишь спустя 30 000 лет после тотального сокращения мегафауны Австралии и Океании.
При этом многие дикие виды животных — волки, верблюды, мамонты, лошади, постоянно мигрировали, и переселялись даже между континентами. Так,
лошадиные
, как семейство, зародились в Северной Америке (смотреть —
Эволюция лошади
) и затем только мигрировали по
Берингии
в Евразию и Африку.
Аргументы против эпидемий как причины вымирания
Во-первых, даже такое высоко
вирулентное
заболевание как
лихорадка Западного Нила
, не вызывает такого массового вымирания и может уничтожить лишь локальные популяции. Популяции, не имеющие контактов с заражённой, отделённые естественными преградами, не будут заражены. Во-вторых, болезнь должна быть чрезвычайно избирательной, заражать строго определённые виды мегафауны, не трогая более мелкие виды. Кроме того, такое заболевание должно иметь крайне широкий ареал (миллионы квадратных километров) с различным климатом, водными и пищевыми ресурсами, а также звеньями пищевых цепей, состоящих из различных животных по видам и особенностям питания. При этом заболевание должно убивать нелетающих птиц, а летающих почти не затрагивать. Науке болезни с таким набором особенностей неизвестны
.
Гипотеза о Хищниках второго порядка
Гипотеза Охоты — тотального убийства человекомl и хищник второго порядка.
Комбинированные гипотезы: изменение климата, избыток + изменение климата, хищник второго порядка + изменение климата.
Сценарий
Гипотеза выводит следующие события. После того как люди начали мигрировать по
Берингии
в Северную Америку, а затем в Южную Америку, они в первую очередь старались уничтожать наиболее опасных для себя соперников — крупных местных хищников. Это происходило как в борьбе за безопасность, так и за новые зоны охоты, люди вступали таким образом в борьбу за места, где можно было охотится на травоядных млекопитающих. Учитывая, что плотоядные животные также не встречали до этого крупных обезьян и
гоминид
в частности, они не понимали опасности, которая им грозит от относительно небольших, в сравнении с бизонами, животных.
В итоге хищные млекопитающие были значительно сокращены в численности в короткий период, а американские львы и
смилодоны
вообще были истреблены. Это вызвало цепную реакцию — травоядные млекопитающие, при наличии огромной кормовой базы и при отсутствии хищников в нужном количестве, начали излишне размножаться.
Основные этапы:
После прибытия Хомо сапиенса в Северную Америку, существующие хищники должны «делиться» охотничьими угодьями с новым конкурентом. Это вызывает конфликт
Хищник второго порядка — Хомо сапиенс — начинает убивать хищников первого порядка.
В итоге хищники первого порядка истреблены почти полностью, нарушается баланс биосистемы, сложившийся в течение миллионов лет, до прихода гоминид в Новый Свет.
При отсутствии регулирования хищниками, численность мелких травоядных резко растёт, после чего начинается кризис кормовой базы. Следом начинается голод для травоядных в силу истощения пастбищ. Под ударом оказываются виды, которые зависят от большого количества сочной травы, такие как хоботные. Следом вымирают животные, по биологическим механизмам не адаптированные к выживанию на малом количестве корма.
В силу давления животных на пастбища, пастбища вытаптываются, меняется характер растительности. Вслед за ним меняется климат, становясь всё более континентальным, влажность падает.
Аргументы за данную теорию
Компьютерная мультивидовая модель вымирания, составленная для плейстоцена (перепромысел первобытными людьми, но с их приоритетным воздействием на хищников I порядка), подтверждает положения теории. Учитываются — численность травоядных, хищников, гоминид и кормовая база, необходимая для каждой ниши. Данная программа и её вводные, пока самые точные, для моделирования ситуаций в глобальном, континентальном масштабе.
Аргументы против
Модель предполагает высокие показатели вымирания на луго-пастбищных угодьях, но большинство вымерших видов жило в разных зонах растительности. Историческая плотность населения копытных была очень высокой на Великих равнинах; саванны поддерживают высокое разнообразие копытных во всей Африке, а интенсивность вымирания была одинаково серьёзной и в лесных условиях.
Не объясняется, почему крупные травоядные популяции не регулировались выжившими плотоядными животными, такими как медведи гризли, волки, пумы и ягуары, чья популяция быстро увеличивалась бы, в ответ на потерю конкурентов.
Это не объясняет, почему почти все вымершие крупные плотоядные животные были узкоспециализированными хищниками, такими как саблезубые кошки и короткомордые медведи, охотившиеся на молодь крупных травоядных, а большинство других крупных хищников выжило.
Нет никаких исторических свидетельств циклов бума и спада, вызывающих даже локальные вымирания в регионах, где крупные хищники, охотившиеся на млекопитающих, были уничтожены охотой. Недавняя охота людей на оставшихся хищников, на большей части Соединённых Штатов, не вызвала массовых изменений в численности копытных.
Всё, что объясняется базовой моделью «плейстоценового перепромысла», также объясняется мультивидовой моделью, но с меньшим количеством допущений, поэтому базовая модель плейстоценового перепромысла выглядит более ограниченной. Однако, многовидовая модель не объясняет сдвиги в растительности и не может имитировать альтернативные гипотезы. Поэтому мультивидовая модель требует дополнительных предположений.
Примечания
↑
Paul S. Martin, Richard G. Klein.
. — The University of Arizona Press, 1984. — 892 с. —
ISBN 0816511004
.
↑
Буровский А.М.
// Биосфера. — 2010. —
Т. 2
,
вып. 1
. —
ISSN
.
29 апреля 2021 года.
Пучков П.В.
// Биосфера. — 2010. —
Т. 2
,
вып. 3
. —
ISSN
.
1 мая 2021 года.
↑
(неопр.)
. www.vokrugsveta.ru. Дата обращения: 28 ноября 2019.
20 мая 2019 года.
↑
(англ.)
. postnauka.ru. Дата обращения: 24 декабря 2019.
23 декабря 2019 года.
↑
Екатерина Русакова.
(неопр.)
. nplus1.ru. Дата обращения: 26 марта 2020.
26 марта 2020 года.
↑
(рус.)
. «Элементы». Дата обращения: 16 апреля 2020.
22 февраля 2020 года.
↑
(рус.)
. «Элементы». Дата обращения: 22 мая 2020.
4 августа 2019 года.
↑
(рус.)
. «Элементы». Дата обращения: 16 апреля 2020.
27 ноября 2020 года.
↑
(неопр.)
www.e-anthropology.com
. Дата обращения: 5 октября 2020.
9 октября 2020 года.
↑
Felisa A. Smith, Rosemary E. Elliott Smith, S. Kathleen Lyons, Jonathan L. Payne.
(англ.)
// Science. — 2018-04-20. —
Vol. 360
,
iss. 6386
. —
P. 310—313
. —
ISSN
. —
doi
:
.
31 марта 2020 года.
↑
(неопр.)
. cyberleninka.ru. Дата обращения: 24 декабря 2019.
20 декабря 2019 года.
Kolbert, Elizabeth (2014).
The Sixth Extinction: An Unnatural History
. Bloomsbury Publishing. ISBN 9780805092998.
Paul L. Koch, Anthony D. Barnosky.
(англ.)
// Annual Review of Ecology, Evolution, and Systematics. — 2006-12. —
Vol. 37
,
iss. 1
. —
P. 215–250
. —
ISSN
. —
doi
:
.
9 марта 2021 года.
Curry, Andrew (2012-05-03). «Ancient migration: Coming to America».
Nature
.
485
(7396): 30-32. doi:10.1038/485030a.
↑
П.В. Пучков.
// Вестник зоологии. — Інститут зоології ім. І. І. Шмальгаузена НАН України, 1992. —
№ 4
. —
С. 73-81
. —
ISSN
.
2 января 2021 года.
↑
П.В. Пучков.
//
Інститут зоології ім. І. І. Шмальгаузена НАН України
Вестник зоологии. — 1991. —
№ 5
. —
С. 45-53
. —
ISSN
.
2 января 2021 года.
↑
Гржимек Б.
Для диких животных места нет / Предисл. и коммент.
А. Г. Банникова
и А. Г. Пегушева. —
М.
: Мысль, 1977. — 270 с.
(неопр.)
.
обращения: 9 декабря 2019.
9 декабря 2019 года.
(неопр.)
. Дата обращения: 1 мая 2018.
1 мая 2018 года.
↑
Frédérik Saltré, Marta Rodríguez-Rey, Barry W. Brook, Christopher N Johnson, Chris S. M. Turney.
// Nature Communications. — 2016-01-29. —
Т. 7
. —
ISSN
. —
doi
:
.
12 ноября 2020 года.
↑
Christopher Sandom, Søren Faurby, Brody Sandel, Jens-Christian Svenning.
// Proceedings of the Royal Society B: Biological Sciences. — 2014-07-22. —
Т. 281
,
вып. 1787
. —
С. 20133254
. —
doi
:
.
19 июля 2020 года.
↑
Paul S. Martin, David W. Steadman.
(англ.)
// Extinctions in Near Time: Causes, Contexts, and Consequences / Ross D. E. MacPhee. — Boston, MA: Springer US, 1999. —
P. 17–55
. —
ISBN 978-1-4757-5202-1
. —
doi
:
.
↑
П. В. Пучков, А. М. Буровский.
//
/ Л.Е. Гринин. —
Российская Академия наук
, МГУ им. Ломоносова. —
М.
: ООО "Издательство "Учитель", 2015. — Vol. 7. — С. 169-218. — 223 с. —
ISBN 978-5-7057-4566-1
.
↑
(рус.)
. «Элементы». Дата обращения: 17 мая 2020.
21 февраля 2020 года.
↑
(неопр.)
. www.vokrugsveta.ru. Дата обращения: 29 ноября 2019.
20 ноября 2019 года.
↑
(рус.)
. «Элементы». Дата обращения: 22 мая 2020.
17 июля 2020 года.
П.В.Пучков.
// Вестник зоологии. — 1992. —
№ 1
. —
ISSN
.
24 марта 2021 года.
↑
(рус.)
.
Газета.Ru
. Дата обращения: 1 декабря 2019.
20 июня 2022 года.
R. Gillespie.
(англ.)
// Quaternary Science Reviews. — 2008. —
Vol. 27
,
iss. 27-28
. —
P. 2522
. —
ISSN
. —
doi
:
.
13 июня 2020 года.
Lisa Nagaoka, Torben Rick, Steve Wolverton.
// Ecology and Evolution. — 2018-09-05. —
Т. 8
,
вып. 19
. —
С. 9683–9696
. —
ISSN
. —
doi
:
.
10 ноября 2020 года.
Werdelin, Lars; Lewis, Margaret E. (2013-03-06). «Temporal Change in Functional Richness and Evenness in the Eastern African Plio-Pleistocene Carnivoran Guild».
PLOS ONE
.
8
(3): e57944. doi:10.1371/journal.pone.0057944. ISSN 1932—620
Tollefson, Jeff (2012). «Early humans linked to large-carnivore extinctions».
Nature
. doi:10.1038/nature.2012.10508.
Wong, Kate. «Rise of Humans 2 Million Years Ago Doomed Large Carnivores».
Scientific American Blog Network
. Retrieved 2017-09-11.
«These Early Humans Lived 300,000 Years Ago—But Had Modern Faces». 2017-06-07. Retrieved 2017-10-13.
Zimmer, Carl (2017-07-04). «In Neanderthal DNA, Signs of a Mysterious Human Migration».
The New York Times
. ISSN 0362-4331. Retrieved 2017-10-13.
«Almost all living people outside of Africa trace back to a single migration more than 50,000 years ago».
Science | AAAS
. 2016-09-20. Retrieved 2017-10-14.
«Human Migration: Challenging the Chronology of Our First Road Trip — Dead Things».
Dead Things
. 2016-09-21. Retrieved 2017-10-14.
Julien Louys; Darren Curnoe; Haowen Tong. (2007). «Characteristics of Pleistocene megafauna extinctions in Southeast Asia».
Palaeogeography, Palaeoclimatology, Palaeoecology
.
243
: 152—173. doi:10.1016/j.palaeo.2006.07.011.
Horowitz, Aharon (2014-05-10).
The Quaternary of Israel
. Academic Press. ISBN 9781483267234.
Yuan Wang, Chang-zhu Jin, Jim I. Mead.
(англ.)
// Quaternary International. — 2014-08-07. —
Vol. 339—340
. —
P. 90—96
. —
ISSN
. —
doi
:
.
↑
(рус.)
. antropogenez.ru. Дата обращения: 26 марта 2020.
26 марта 2020 года.
Pavel Kosintsev, Kieren J. Mitchell, Thibaut Devièse, Johannes van der Plicht, Margot Kuitems.
(англ.)
// Nature Ecology & Evolution. — 2019-01. —
Vol. 3
,
iss. 1
. —
P. 31–38
. —
ISSN
. —
doi
:
.
6 октября 2021 года.
↑
Rhodin, A.G.J., Thomson, S., Georgalis, G., Karl, H.-V., Danilov, I.G., Takahashi, A., de la Fuente, M.S., Bourque, J.R., Delfino, M., Bour, R., Iverson, J.B., Shaffer, H.B., and van Dijk, P.P.]
// IUCN/SSC Tortoise and Freshwater Turtle Specialist Group. — 2015. —
С. 15, 23-24
. —
ISBN 978-0-9653540-9-7
. —
doi
:
.
1 августа 2020 года.
Giles Hamm, Peter Mitchell, Lee J. Arnold, Gavin J. Prideaux, Daniele Questiaux.
(англ.)
// Nature. — 2016-11. —
Vol. 539
,
iss. 7628
. —
P. 280–283
. —
ISSN
. —
doi
:
.
21 апреля 2021 года.
↑
(рус.)
. «Элементы». Дата обращения: 17 мая 2020.
12 октября 2019 года.
↑
Chris S. M. Turney, Timothy F. Flannery, Richard G. Roberts, Craig Reid, L. Keith Fifield.
// Proceedings of the National Academy of Sciences of the United States of America. — 2008-08-26. —
Т. 105
,
вып. 34
. —
С. 12150–12153
. —
ISSN
. —
doi
:
.
8 марта 2021 года.
C. N. Johnson, J. Alroy, N. J. Beeton, M. I. Bird, B. W. Brook.
// Proceedings of the Royal Society B: Biological Sciences. — 2016-02-10. —
Т. 283
,
вып. 1824
. —
ISSN
. —
doi
:
.
14 ноября 2020 года.
↑
(рус.)
. «Элементы». Дата обращения: 17 мая 2020.
4 августа 2019 года.
Marwick, Ben. «Buried tools and pigments tell a new history of humans in Australia for 65,000 years».
The Conversation
. Retrieved 2017-09-07
Webb, Steve (2013-02-27).
Corridors to Extinction and the Australian Megafauna
. Newnes. ISBN 9780124078406.
Lourandos, Harry (1997-02-28).
Continent of Hunter-Gatherers: New Perspectives in Australian Prehistory
. Cambridge University Press. ISBN 9780521359467.
Long, John A.; Archer, Michael (2002-01-01).
Prehistoric Mammals of Australia and New Guinea: One Hundred Million Years of Evolution
. UNSW Press. ISBN 9780868404356.
Stephen Wroe, Judith H. Field, Michael Archer, Donald K. Grayson, Gilbert J. Price.
// Proceedings of the National Academy of Sciences of the United States of America. — 2013-05-28. —
Т. 110
,
вып. 22
. —
С. 8777–8781
. —
ISSN
. —
doi
:
.
14 февраля 2021 года.
Alan Cooper, Chris S. M. Turney, Jonathan Palmer, Alan Hogg, Matt McGlone.
(англ.)
// Science. — 2021-02-19. —
Vol. 371
,
iss. 6531
. —
P. 811–818
. —
ISSN
. —
doi
:
.
31 августа 2021 года.
(англ.)
.
ScienceDaily
. Дата обращения: 22 февраля 2021.
22 февраля 2021 года.
(англ.)
.
phys.org
. Дата обращения: 22 февраля 2021.
19 февраля 2021 года.
(амер. англ.)
.
Science News
(18 февраля 2021). Дата обращения: 24 июля 2021.
19 февраля 2021 года.
Редакция журнала Наука и жизнь.
(рус.)
. www.nkj.ru. Дата обращения: 16 апреля 2020.
15 декабря 2019 года.
Серебрянный Леонид Рувимович.
Вымирание животных в позднем плейстоцене
// Древнее оледенение и жизнь. —
М.
: Наука, 1980. — 128 с.
Zimov, S. A.; Zimov, N. S.; Tikhonov, A. N.; Chapin III, F. S. (2012-12-04). «Mammoth steppe: a high-productivity phenomenon».
Quaternary Science Reviews
.
57
: 26-45. Bibcode:2012QSRv…57…26Z. doi:10.1016/j.quascirev.2012.10.005.
Zimov, S. A.; Zimov, N. S.; Tikhonov, A. N.; Chapin III, F. S. (2012-12-04). «Mammoth steppe: a high-productivity phenomenon».
Quaternary Science Reviews
.
57
: 26-45. Bibcode:2012QSRv…57…26Z. doi:10.1016/j.quascirev.2012.10.005
Sher, A. V.; Kuzmina, S. A.; Kuznetsova, T. V.; Sulerzhitsky, L. D. (2005-03-01). «New insights into the Weichselian environment and climate of the East Siberian Arctic, derived from fossil insects, plants, and mammals».
Quaternary Science Reviews
.
24
(5-6): 533—569. Bibcode:2005QSRv…24..533S. doi:10.1016/j.quascirev.2004.09.007.
Hoffecker, John F.; Elias, Scott A. (2012-05-29).
Human Ecology of Beringia
. Columbia University Press. ISBN 9780231503884.
Vereshchagin, N. K.; Baryshnikov, G. F. (1991-01-01). «The ecological structure of the „Mammoth Fauna“ in Eurasia».
Annales Zoologici Fennici
.
28
(3/4): 253—259. JSTOR 23735450.
Adams, J. M.; Faure, H.; Faure-Denard, L.; McGlade, J. M.; Woodward, F. I. (1990-12-27). «Increases in terrestrial carbon storage from the Last Glacial Maximum to the present».
Nature
.
348
(6303): 711—714. Bibcode:1990Natur.348..711A. doi:10.1038/348711a0.
Álvarez-Lao, Diego J.; García, Nuria (2011-03-15). «Geographical distribution of Pleistocene cold-adapted large mammal faunas in the Iberian Peninsula».
Quaternary International
. Quaternary Floral and Faunal Assemblages: Ecological and Taphonomical Investigations.
233
(2): 159—170. Bibcode:2011QuInt.233..159A. doi:10.1016/j.quaint.2010.04.017.
↑
(рус.)
. «Элементы». Дата обращения: 8 марта 2020.
15 декабря 2019 года.
Marc A. Carrasco, Anthony D. Barnosky, Russell W. Graham.
(англ.)
// PLOS ONE. — 2009-12-16. —
Vol. 4
,
iss. 12
. —
P. e8331
. —
ISSN
. —
doi
:
.
14 декабря 2014 года.
Todd A. Surovell, Spencer R. Pelton, Richard Anderson-Sprecher, Adam D. Myers.
(англ.)
// Proceedings of the National Academy of Sciences. — 2016-01-26. —
Vol. 113
,
iss. 4
. —
P. 886–891
. —
ISSN
. —
doi
:
.
22 мая 2020 года.
(рус.)
. «Элементы». Дата обращения: 26 мая 2020.
16 декабря 2019 года.
↑
Редакция журнала Наука и жизнь.
(рус.)
. www.nkj.ru. Дата обращения: 8 марта 2020.
15 декабря 2019 года.
↑
(англ.)
. Дата обращения: 8 марта 2020.
9 декабря 2019 года.
(рус.)
. www.vokrugsveta.ru. Дата обращения: 8 марта 2020.
19 июля 2019 года.
(неопр.)
. evolbiol.ru. Дата обращения: 22 мая 2020.
4 июня 2020 года.
Doughty, C. E.; Wolf, A.; Field, C. B. (2010). «Biophysical feedbacks between the Pleistocene megafauna extinction and climate: The first human‐induced global warming?».
Geophys. Res. Lett
.
37
(15): L15703.
Grayson, Donald K.; Meltzer, David J. (December 2012). «Clovis Hunting and Large Mammal Extinction: A Critical Review of the Evidence».
Journal of World Prehistory
.
16
(4): 313—359
Kolbert, Elizabeth (2014).
The Sixth Extinction: An Unnatural History
. Bloomsbury Publishing.
Perry, George L. W.; Wheeler, Andrew B.; Wood, Jamie R.; Wilmshurst, Janet M. (2014-12-01). «A high-precision chronology for the rapid extinction of New Zealand moa (Aves, Dinornithiformes)».
Quaternary Science Reviews
.
105
: 126—135.
Crowley, Brooke E. (2010-09-01). «A refined chronology of prehistoric Madagascar and the demise of the megafauna».
Quaternary Science Reviews
. Special Theme: Case Studies of Neodymium Isotopes in Paleoceanography.
29
(19-20): 2591—2603.
(неопр.)
. Дата обращения: 5 мая 2018.
20 июня 2018 года.
Vernon Smith.
// Economics Faculty Articles and Research. — 1975-01-01.
15 апреля 2021 года.
(рус.)
. vestifinance.ru. Дата обращения: 29 ноября 2019.
7 ноября 2018 года.
Marcella Alsan.
(англ.)
// American Economic Review. — 2015/01. —
Vol. 105
,
iss. 1
. —
P. 382–410
. —
ISSN
. —
doi
:
.
26 апреля 2022 года.
(рус.)
. www.krugosvet.ru. Дата обращения: 16 апреля 2020.
22 октября 2020 года.
Mossiman, J. E. & Martin, P. S. (1975). «Simulating Overkill by Paleoindians».
American Scientist
.
63
: 304-13. Bibcode:1975AmSci..63..304M.
Whittington, S. L. & Dyke, B. (1984). «Simulating overkill: experiment with the Mossiman and Martin model». In Martin, P.S. & Klein, R.G.
Quaternary extinctions: A prehistoric revolution
. Tucson, AZ: Univ. Arizona Press. pp. 451-66. ISBN 0-8165-1100-4.
Lewis J. Bartlett, David R. Williams, Graham W. Prescott, Andrew Balmford, Rhys E. Green.
(англ.)
// Ecography. — 2016. —
Vol. 39
,
iss. 2
. —
P. 152–161
. —
ISSN
. —
doi
:
.
15 мая 2021 года.
Diamond J (2008). «Palaeontology: The last giant kangaroo».
Nature
.
454
(7206): 835-6. Bibcode:2008Natur.454..835D. doi:10.1038/454835a. PMID 18704074.
Martin, P. S. (2005).
Twilight of the Mammoths: Ice Age Extinctions and the Rewilding of America
. University of California Press. ISBN 0-520-23141-4.
Burney, D. A.; Flannery, T. F. (July 2005). «Fifty millennia of catastrophic extinctions after human contact» (PDF).
Trends in Ecology & Evolution
.
20
(7): 395—401. doi:10.1016/j.tree.2005.04.022. PMID 16701402. Archived from the original (PDF) on 2010-06-10.
Flannery, Tim (2002-10-16).
The future eaters: an ecological history of the Australasian lands and people
. New York: Grove/Atlantic, Inc. ISBN 0-8021-3943-4. OCLC 32745413.
Diamond, J. (1984). «Historic extinctions: a Rosetta stone for understanding prehistoric extinctions». In Martin, P.S.; Klein, R.G.
Quaternary extinctions: A prehistoric revolution
. Tucson, AZ: Univ. Arizona Press. pp. 824-62. ISBN 0-8165-1100-4.
Diamond, J. (1997).
Guns, germs, and steel; the fates of human societies
. New York: Norton. ISBN 0-393-31755-2.
Alroy, J. (2001). «A multispecies overkill simulation of the end-Pleistocene megafaunal mass extinction» (PDF).
Science
.
292
(5523): 1893-6. Bibcode:2001Sci…292.1893A. doi:10.1126/science.1059342. PMID 11397940.
«Humans responsible for demise of gigantic ancient mammals». University of Exeter. 13 August 2015. Retrieved 14 August 2015.
(рус.)
.
Газета.Ru
. Дата обращения: 1 декабря 2019.
20 июня 2022 года.
↑
Редакция журнала Наука и жизнь.
(рус.)
.
www.nkj.ru
. Дата обращения: 2 августа 2020.
17 апреля 2021 года.
(рус.)
. russian7.ru. Дата обращения: 1 декабря 2019.
9 июля 2018 года.
May, R. M. (2001).
Stability and complexity in model ecosystems
. Princeton: Princeton Univ. Press. ISBN 0-691-08861-6.
Grayson, D.K.; Meltzer, D.J. (2003). «A requiem for North American overkill».
Journal of Archaeological Science
.
30
(5): 585—593. doi:10.1016/s0305-4403(02)00205-4.
Fiedel, S., Haynes, G., 2004. A premature burial: comments on Grayson and Meltzer’s
Requiem for overkill
Journal of Archaeological Science 31, 121—131.
Scott, J. (2009-02-26). «Camel-butchering in Boulder, 13,000 years ago».
Colorado Arts and Sciences Magazine
. University of Colorado at Boulder. Archived from the original on 2010-06-10. Retrieved 2009-05-01.
Surovell, Todd A; Brigid S Grund (2012). «The associational critique of Quaternary overkill».
American Antiquity
.
77
(4): 673—688
Bell, C.J.; et al. (2004). «The Blancan, Irvingtonian, and Rancholabrean mammal ages». In Woodburne, M.O.
Late Cretaceous and Cenozoic Mammals of North America: Biostratigraphy and Geochronology
. New York: Columbia Univ. Press.
Scott, E., Cox, S.M. (2008). «Late Pleistocene distribution of
Bison
(Mammalia; Artiodactyla) in the Mojave Desert of Southern California and Nevada». In Wang, X.; Barnes, L.G.
Geology and Vertebrate Paleontology of Western and Southern North America
. Los Angeles: Natural History Museum of Los Angeles County. pp. 359—382.
Sanders, A.E., R.E. Weems, and L.B. Albright III (2009). «Formalization of the mid-Pleistocene „Ten Mile Hill beds“ in South Carolina with evidence for placement of the Irvingtonian-Rancholabrean boundary». In Albright III, L.B.
Papers on Geology, Vertebrate Paleontology, and Biostratigraphy in Honor of Michael O. Woodburne
. Flagstaff: Museum of Northern Arizona. pp. 369—375.
Shapiro, B.; et al. (2004). «Rise and Fall of the Beringian Steppe Bison».
Science
.
306
(5701): 1561—1565. Bibcode:2004Sci…306.1561S. doi:10.1126/science.1101074
Wilsom, M.C., L.V. Hills, and B. Shapiro (2008). «Late Pleistocene northward-dispersing
Bison antiquus
from the Bighill Creek Formation, Gallelli Gravel Pit, Alberta, Canada, and the fate of
Bison occidentalis
».
Canadian Journal of Earth Sciences
.
45
(7): 827—859.
Nadasdy, Paul (2006). «Transcending the Debate over the Ecologically Noble Indian: Indigenous Peoples and Environmentalism».
Ethnohistory
.
52
(2): 291—331.
Svenson, Sally. «Bison Archived 2008-09-13 at the Wayback Machine..» South Dakota Department of Game, Fish and Parks, 1995.
(неопр.)
. Дата обращения: 5 мая 2018.
5 мая 2018 года.
↑
Scott, E. (2010). «Extinctions, scenarios, and assumptions: Changes in latest Pleistocene large herbivore abundance and distribution in western North America».
Quat. Int
.
217
: 225—239.
Willis, Paul; Bryce, Clay; Searle, Mike (17 August 2006). «Thylacoleo — The Beast of the Nullarbor».
Catalyst
http://www.abc.net.au/catalyst/stories/s1717424.htm
Prideaux GJ, Long JA, Ayliffe LK, et al. (January 2007). «An arid-adapted middle Pleistocene vertebrate fauna from south-central Australia».
Nature
.
445
(7126): 4
Roberts RG, Flannery TF, Ayliffe LK, et al. (June 2001). «New ages for the last Australian megafauna: continent-wide extinction about 46,000 years ago».
Science
.
292
(5523): 1888-92.
Louys, Julien; Curnoe, D.; Tong, H. (2007). «Characteristics of Pleistocene megafauna extinctions in Southeast Asia».
Palaeogeography, Palaeoclimatology, Palaeoecology
.
243
(1-2): 152—173.
Fisher, Daniel C. (2009). «Paleobiology and Extinction of Proboscideans in the Great Lakes Region of North America». In Haynes, Gary.
American Megafaunal Extinctions at the End of the Pleistocene
. Springer. pp. 55-75.
(неопр.)
. www.dopotopa.com. Дата обращения: 15 апреля 2020.
19 февраля 2020 года.
↑
Rabanus-Wallace, M. Timothy; Wooller, Matthew J.; Zazula, Grant D.; Shute, Elen; Jahren, A. Hope; Kosintsev, Pavel; Burns, James A.; Breen, James; Llamas, Bastien; Cooper, Alan (2017). «Megafaunal isotopes reveal role of increased moisture on rangeland during late Pleistocene extinctions».
Nature Ecology & Evolution
.
1
(5): 0125.
Birks, H.H. (1973). «Modern macrofossil assemblages in lake sediments in Minnesota». In Birks, H.J.B.; West, R.G.
Quaternary plant ecology: the 14thsymposium of the British Ecological Society, University of Cambridge, 28-30 March 1972
. Oxford: Blackwell Scientific
Davis, M. B. (1976). «Pleistocene biogeography of temperate deciduous forests».
Geoscience and man: Ecology of the Pleistocene
.
13
. Baton Rouge: School of Geoscience, Louisiana State Univ
Guthrie, R. D. (1988).
Frozen Fauna of the Mammoth Steppe: The Story of Blue Babe
. University Of Chicago Press.
Guthrie, R. D. (1989). «Mosaics, allochemics, and nutrients: an ecological theory of Late Pleistocene megafaunal extinctions». In Martin, P.S.; Klein, R.G.
Quaternary extinctions: A prehistoric revolution
. Tucson, AZ: Univ. Arizona Press. pp. 259-99.
Hoppe, P.P. (1978). «Rumen fermentation in African ruminants».
Proceedings of the 13thAnnual Congress of Game Biologists
. Atlanta.
(неопр.)
elementy.ru. Дата обращения: 22 мая 2020.
4 июня 2020 года.
Bryson, R.A., Baerreis, D.A., Wendland, W.M. (1970). «The character of late-glacial and post-glacial climatic changes». In Dort Jr., W.; Jones, Jr. J.K.
Pleistocene and recent environments of the central Great Plains
. Lawrence: Univ. Press Kan.
Graham, R.W., Lundelius, E.L. (1989). «Coevolutionary disequilibrium and Pleistocene extinctions». In Martin, P.S.; Klein, R.G.
Quaternary extinctions: A prehistoric revolution
. Tucson, AZ: Univ. Arizona Press. pp. 354—404.
King, J.E., Saunders, J.J. (1989). «Environmental insularity and the extinction of the American mastodont». In Martin, P.S.; Klein R.G.
Quaternary extinctions: A prehistoric revolution
. Tucson, AZ: Univ. Arizona Press. pp. 354—404.
Axelrod, D. I. (1967). «Quaternary extinctions of large mammals».
University of California Publications in Geological Sciences
.
74
: 1-42. ASIN B0006BX8LG.
Slaughter, B. H. (1967). «Animal ranges as a clue to late-Pleistocene extinction». In Martin, P.S.; Wright H.E.
Pleistocene extinctions: The search for a cause
. New Haven: Yale Univ. Press
Kilti, R. A. (1988). «Seasonality, gestation time, and large mammal extinctions». In Martin, P.S.; Klein R.G.
Quaternary extinctions: A prehistoric revolution
. Tucson, AZ: Univ. Arizona Press. pp. 354—404. I
Andersen, S. T (1973). «The differential pollen productivity of trees and its significance for the interpretation of a pollen diagram from a forested region». In Birks, H.J.B.; West, R.G.
Quaternary plant ecology: the 14thsymposium of the British Ecological society, University of Cambridge, 28-30 March 1972
. Oxford: Blackwell Scientific.
Ashworth, C.A. (1980). «Environmental implications of a beetle assemblage from the Gervais formation (Early Wisconsinian?), Minnesota».
Quat. Res
.
13
(2): 200-12.
Birks, H.H. (1973). «Modern macrofossil assemblages in lake sediments in Minnesota». In Birks, H.J.B.; West, R.G.
Quaternary plant ecology: the 14thsymposium of the British Ecological Society, University of Cambridge, 28-30 March 1972
. Oxford: Blackwell Scientific.
Bradley, R. S. (1985).
Quaternary Paleoclimatology: Methods of Paleoclimatic Reconstruction
. Winchester, MA: Allen & Unwin.
Davis, M. B. (1976). «Pleistocene biogeography of temperate deciduous forests».
Geoscience and man: Ecology of the Pleistocene
.
13
. Baton Rouge: School of Geoscience, Louisiana State Univ.
(рус.)
.
Газета.Ru
. Дата обращения: 1 декабря 2019.
20 июня 2022 года.
Flereov, C.C. (1967). «On the origin of the mammalian fauna of Canada». In Hopkins, D.M.
The Bering Land Bridge
. Palo Alto: Stanford Univ. Press. pp. 271-80
Frenzel, B. (1968). «The Pleistocene vegetation of northern Eurasia».
Science
.
161
(3842): 637-49.
↑
McDonald, J. (1989). «The reordered North American selection regime and late Quaternary megafaunal extinctions». In Martin, P.S.; Klein, R.G.
Quaternary extinctions: A prehistoric revolution
. Tucson, AZ: Univ. Arizona Press. pp. 354—404.
Birks, H.J.B., West, R.G. (1973).
Quaternary plant ecology: the 14th symposium of the British Ecological society, University of Cambridge, 28-30 March 1972
. Oxford: Blackwell Scientific.
McDonald, J. (1981).
North American Bison: Their classification and evolution
. Berkeley: Univ. Calif. Press.
Burney, D. A. (1993). «Recent animal extinctions: recipes for disaster».
American Scientist
.
81
(6): 530-41.
Vartanyan, S.L., Garutt, V. E. and Sher, A.V. (1993). «Holocene dwarf mammoths from Wangel Island in the Siberian Arctic».
Nature
.
362
(6418): 337-40.
Pennycuick, C.J. (1979). «Energy costs of locomotion and the concept of „Foraging radius“». In Sinclair A.R.E.; Norton-Griffiths M.
Serengetti: Dynamics of an Ecosystem
. Chicago: Univ. Chicago Press. pp. 164-85. I
Owen-Smith, R.N. (1992).
Megaherbivores: The influence of very large body size on ecology. Cambridge studies in ecology
. Cambridge: Cambridge Univ.
Kershaw, G.P. (1984). «Tundra plant communities of the Mackenzie mountains, Northwest Territories; floristic characteristics of long term surface disturbances». In Olson, R.; Hastings, R.; Geddes, F.
Northern Ecology and Resource Management: Memorial Essays honoring Don Gill
. Edmonton, Canada: Univ. Alberta Press. pp. 239—311
Webber, P.J., Miller, P.C., Chapin, F.S. III, MacCown, B.H. (1980). «The vegetation: pattern and succession». In Brown, J.; Miller, P.C.; Tieszen, L.L.; Bunnell, F.L.
An Arctic ecosystem: the coastal tundra at Barrow, Alaska
. US/IBP Synthesis. Stroudsburg, PA: Dowden Hutchinson & Ross. pp. 186—219. 12.
MacPhee, Ross D.E.; Preston Marx (1997). «The 40,000-year Plague: Humans, Hyperdisease, and First-Contact Extinctions».
Natural Change and Human Impact in Madagascar
. Washington, D.C.: Smithsonian Institution Press. pp. 169—217.
MacFee, R.D.E. & Marx, P.A. (1997). «Humans, hyperdisease and first-contact extinctions». In Goodman, S. & Patterson, B.D.
Natural Change and Human Impact in Madagascar
. Washington D.C.: Smithsonian Press. pp. 169—217. ISBN 1-56098-683-2.
S. Kathleen Lyons, Felisa A. Smith, Peter J. Wagner, Ethan P. White, James H. Brown.
(англ.)
// Ecology Letters. — 2004. —
Vol. 7
,
iss. 9
. —
P. 859–868
. —
ISSN
. —
doi
:
.
15 января 2022 года.
Lyons, K, Smith, F.A., Wagner, P.J., White, E.P., and Brown, J.H. (2004). «Was a 'hyperdisease' responsible for the late Pleistocene megafaunal extinction?» (PDF).
Ecology Letters
.
7
Whitney-Smith, E. (2004). «Late Pleistocene extinctions through second-order predation». In Barton, C.M.; Clark, G.A.; Yesner, D.R.
Settlement of the American Continents: A Multidisciplinary Approach to Human Biogeography
. Tucson, AZ: University of Arizona Press
Whitney-Smith, E. (2009). The Second-Order Predation Hypothesis of Pleistocene Extinctions: A System Dynamics Model. Saarbruken, Germany:
Whitney-Smith, E. (2006).
Clovis and Extinctions — Overkill, Second Order Predation, Environmental Degradation in a Non-equilibrium Ecosystem «Clovis Age Continent»
. University of New Mexico Press.
Scott, E. (2010). «Extinctions, scenarios, and assumptions: Changes in latest Pleistocene large herbivore abundance and distribution in western North America». Quat. Int.
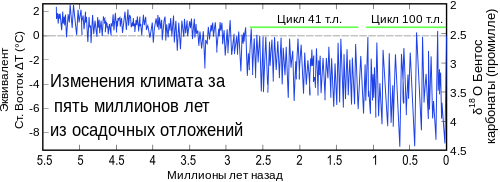


_in_each_TDWG_country_during_the_last_132_000_years,_only_counting_extinctions_earlier_than_1000_years_BP.jpg)




























.jpg)