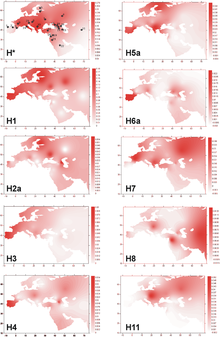
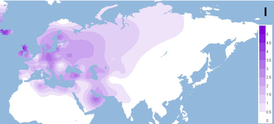
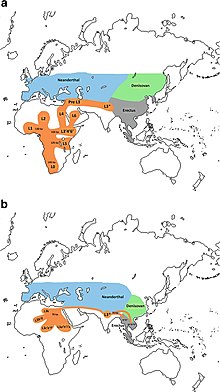
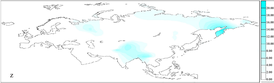
Interested Article - Гаплогруппа U (мтДНК)

- 2020-10-05
- 1
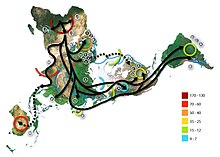
В популяционной генетике человека гаплогруппой U называют одну из гаплогрупп, выявленных при анализе последовательности мутаций в митохондриальной ДНК (mtDNA). Эта гаплогруппа широко распространена в Европе, её носители отделились от макрогруппы R, подгруппы в гаплогруппе N , около 60 тыс. лет назад. За прошедшее с тех пор время гаплогруппа U, кроме Европы, распространилась также в Северной Африке, Аравии, Индии и на Ближнем Востоке .
Разделилась на 8 подгрупп, от гаплогруппы U8 отделилась также гаплогруппа К .
Гаплогруппу U определили у образца MA-1 со стоянки Мальта́ (23 891—24 423 л. н.) .
Гаплогруппу U, содержащую семь «диагностических» позиций, которые отличают её от последовательностей мтДНК других людей ( Дольни-Вестонице -14, Лошбур, Оберкассель, саамы), определили у образца DCP1 из Денисовой пещеры (от 19 000 до 25 000 л. н.) .
Гаплогруппа U1
В основном встречается на Ближнем Востоке, хотя с более низкой частотой может быть также обнаружена во всём Средиземноморье. Например, U1a распространена на большом пространстве от Индии до Атлантики, но очень редко встречается на западных и северных берегах Европы, в том числе на Британских островах и в Скандинавии. Несколько носителей обнаружено в Италии (в Тоскане). В Индии U1a найдена в западной части. U1b распространена в тех же регионах, но с ещё меньшей частотой. Немногочисленные носители обнаружены в еврейской диаспоре. В Восточной Европе U1a и U1b распространены с одинаковой частотой . U1a1a определена у образцов из римско-христианского кладбища Келлис 2 (K2) в египетском оазисе Дахла . U1b2 определена у образца из карельской Хийтолы (1200—1500 гг.) в Лахденпохском районе .
Гаплогруппа U5
Наряду с U8a, U5 — старейшая гаплогруппа в Европе. Возраст — от 50 до 60 тыс. лет . Чаще всего U5 встречается на севере, среди саамов , финнов и эстонцев (преимущественно U5b), но с низкой частотой может быть обнаружена и в остальных частях Европы, на Ближнем Востоке и в Африке . Возраст финской линии U5b1b2 составляет около 6000 лет .
U5 определили у палеолитических обитателей Дольни-Вестонице ( граветтская культура ), живших 31 тыс. лет назад .
U5b2b определили у двух поздних эпиграветтских образцов из сицилийской пещеры (San Teodoro 3 и San Teodoro 5), живших после последнего ледникового максимума (LGM) 15 322—14 432 лет назад .
U5b2b определили у образца Villabruna 1 (ок. 14 тыс. л. н.) из локации Рипари Виллабруна в Северной Италии (коммуна Соврамонте ) .
U5b1h определили у палеолитического обитателя в Швейцарии, жившего 13 560—13 770 лет назад .
U5b1 определили у представителя мадленской культуры из ( ), жившего 13,4 тыс. лет назад .
U5a2 определили у образца Sidelkino (11,55 тыс. л. н.) со стоянки Гора Маяк у села Сиделькино Самарской области .
U5b2 определили у образца HG1 из погребения 37 (Vasilyevka 3, 11 143—10 591 л. н.) в Васильевке-на-Днепре .
U5b1 определили у образца Arene Candide 16 (10 810±65 л. н. ) из пещеры в Италии .
U5a2 определили у образца I1737 (9000—7500 лет до н. э.) из Василевки-на-Днепре .
U5b2b у определили мезолитического образца AVH-1 (10,211—9,91 тыс. л. н.) из пещеры Арма Вейрана (Arma Veirana) в Лигурийских Альпах ( Италия ) .
U5a2d определили у трёх человек с мезолитической стоянки Гусиный Клюв (Huseby Klev) на западе Швеции (о. Оруст), живших около 9880—9540 лет назад. ДНК выделена из слюны, извлечённой из фрагментов дёгтя .

Эта гаплогруппа обнаружена у темнокожего и голубоглазого Чеддарского человека (U5b1) — мумии эпохи мезолита из Англии, а также у ископаемых скелетов охотников-собирателей из Германии, Польши, Литвы и Испании .
U5b2 определили у трёх мезолитических образцов из Василевки-на-Днепре: I1819 (8825—8561 лет до н. э.), I1763 (8280—7967 лет до н. э.), I1734 (7446—7058 лет до н. э.) .
U5a2 определили у мезолитического образца Lec2 (10,39 тыс. л. н.) из крымской пещеры Лесника (Lesnik Cave), известной также как Медвежья пещера (Bear Cave) .
U5 определили у темнокожего мезолитического (5998 лет до н. э.) гаплогруппу .
U5b2c1 определили у темнокожего и голубоглазого мезолитического охотника-собирателя из пещеры , жившего около 7 тыс. лет назад на северо-западе Иберийского полуострова, и у его брата La Braña 2 .
U5a1 определили у неолитического образца I1958 (4723—4558 гг. до н. э.) со стоянки со стоянки Мергень 6 на озере Мергень в Тюменской области .
U5b1b1 (U5b1b-a* ) определили у неолитического образца I4893 (4446—4347 лет до н. э.) из пражского ( Чехия ) .
U5a1i определили у представителя хвалынской культуры .
U5 определили у представителя культуры кардиальной керамики .
U5b определили у трёх представителей неолитической сернийской культуры (север Франции) .
U5a2+16362 определили у представителя трипольской культуры (3789—3650 лет до н. э.) из пещеры Вертеба .
U5a1 (U5a1i ) обнаружена у представителя катакомбной культуры RK4001 (2451—2203 лет до н. э.) .
U5 определили у представителей ямной культуры .
U5a1a обнаружили у представителей днепро-донецкой культуры .
U5a1d1 определили у образцов RV 1852 и 2018/1 со стоянки Риннюкалнс (Riņņukalns) на реке Салаца в Буртниекском крае на севере Латвии, U5a2b2 определили у образца RV 2039 (5300—5050 л. н.). Кроме того, у RV 2039 удалось зафиксировать следы возбудителя чумы грамотрицательной бактерии Yersinia pestis .
U5 определили у представителей межовской культуры .
U5a1a2a, U5a1b1 и U5a2c определили представителей унетицкой культуры из Чехии (Czech_EBA_Unetice 3850—4100 л. н.) .
U5b2b5 обнаружили у 4000-летней головы египетской мумии из гробницы в коптской деревне Дейр эль-Берша .
U5b2b5 обнаружили у 4000-летней мумии Kadruka 1 с севера Судана (зуб KDR001.A) .
U5a1a2a определили у образца I4773 (1618—1513 гг. до н. э.) из среднего-позднего бронзового века Казахстана (Aktogai_MLBA) .
Редкий субклад U5b2c1 обнаружен у из пунического погребального склепа на холме Бирса недалеко от входа в Национальный музей Карфагена в Тунисе возрастом около 2500 лет назад (конец VI века до н. э.) . Предполагается, что эта линия унаследована из Испании, колонизированной финикийцами .
В финском были определены U5a, U5b и характерная для саамов U5b1b1a, у двух образцов из Холлолы (955—1390 гг.) определена гаплогруппа U5a2a1e .
U5a1a определили у одного из викингов с языческого захоронения Галгедил (Galgedil) на датском острове Фюн (700—1100 гг. н. э.) .
U5b1g определили у средневекового (X век) жителя Исландии с озера Миватн .
U5a1g выявлена у крестоносца из Сидона (Ливан), жившего примерно в XI—XIII веках .
U5a1b1, U5a2, U5a2a1c определили у трёх женщин из Рубленого города в Ярославле (массовое захоронение № 76, 1238 год) .
U5a2a1 определили у человека, жившего 700 л. н. и найденного на восточном Памире ( Ташкурган ), что указывает на происхождение от доисторического скотоводческого населения Волго-Уральского региона . U5a2a1 сохранился у некоторых представителей памирских народов .
U5a1a1 определили у пятимесячного эмбриона win002_all (Y-хромосомная гаплогруппа R1b-DF17>R1b1a1b1a1a2a1a2~-BY1806 ), обнаруженного в гробу епископа Лундского Педера Винструпа , похороненного в 1679 году .
Подгруппа U5a считается особо восприимчивой к синдрому приобретённого иммунного дефицита .
Гаплогруппа U6

На уровне 10 % распространена в Северной Африке, особенно среди алжирских берберов (29 %) , а также на Пиренейском полуострове, Канарских островах и в Восточной Африке. Хотя на Пиренеях распространение гаплогруппы U6 достигает максимума, считается, что возникла она не здесь, а либо в Леванте 40 тыс. лет назад , либо в Восточной Африке между 25 и 66 тыс. лет назад .
Примерно 45—40 тыс. л. н. преобладающие ныне североафриканские гаплогруппы M1 и U6 возникли в Юго-Западной Азии и вместе двинулись в Африку .
U6 начинает расширяться ~ 26 000 л. н.
Среди трёх основных подгрупп U6 :
- U6a распространена от Канарских островов и Пиренейского полуострова до Сирии, Эфиопии и Кении с наивысшей частотой в Восточной Африке. Возраст: 24 тыс. л. н. Выделяется также U6a1, распространённая в тех же регионах, но выделившаяся около 15 — 20 тыс. лет назад.
- U6b встречается чаще в западных регионах, в Испании — на севере страны, кроме неё — в Марокко, Алжире, Сенегале и Нигерии. Возраст: 8,5 — 12 тыс. лет. Подгруппа U6b1, выделившаяся около 6 тыс. лет назад, встречается только на Пиренейском полуострове и Канарских островах.
- U6c имеется только в Марокко и на Канарских островах. Возраст: 6 — 17,5 тыс. лет. Не имеет мутации (16219), общей для U6a и U6b.
Базальная гаплогруппа U6* была обнаружена у образца PM1 возрастом 35 тыс. л. н. из румынской «пещеры Женщин» ( ), что свидетельствует об обратной верхнепалеолитической миграции из Западной Евразии в Северную Африку .
U6 определена у обитателя грузинской пещеры (Dzudzuana Cave), жившего 26 тыс. лет назад .
Субклады U6d3, U6a1b, U6a6b, U6a7b определены у образцов иберо-мавританской культуры из марокканского Тафоральта (ок. 14 тыс. л. н.) .
U6d определена у энеолитического (4500—3900/3800 гг. до н. э.) образца I1187 из израильской пещеры Пкиин (Peqi’in Cave) .
U6a1a1 определили у образца UE2298/MS060 из исламского некрополя XI века в городе Сегорбе (Кастельон, Валенсия, Испания) .
U6a3 определили у образца VK116 (XII—XIII века) из Тронхейма .
U2'3'4'7'8'9
Субклады U2, U3, U4, U7, U8 и U9 теперь считаются монофилетическими , их общий предок "U2'3'4'7'8'9" определяется мутацией A1811G, возникшей примерно между 42 000 и 48 000 годами назад (Behar et al., 2012). В пределах U2'3'4'7'8'9, субклады U4 и U9 могут быть монофилетическими, как "U4'9" (мутации T195C!, G499A, T5999C), возникшие между 31 000 и 43 000 лет назад (Behar et al., 2012).
U2'3'4'7'8'9 был найден в останках двух древних северных сибиряков (Ancient North Siberians, ANS) возрастом 31 600 л. н. с Янской стоянки , палеолитическая стоянка человека современного типа в Арктике ( Якутия ) .
U2'3'4'7'8'9 был найден у образца Paglicci108 (28 тыс. л. н.) из итальянской пещеры Пальиччи . Граветтская культура археологическая культура кроманьонцев позднего палеолита. .
U2'3'4'7'8'9 был найден у образца Rigney1 (15,690-15,240 тыс. л. н.), Кластер Эль-Мирон (El Mirón Cluster), Мадленская культура во Франции
U2'3'4'7'8'9 был найден у образцов эпиграветтской культуры Ориенте С (14200-13800 кал. лет назад) с острова , у позднего эпиграветтского индивида San Teodoro 2 из на Сицилии , мезолитического индивида Oriente B (9 377 ± 25 л. н.) с острова Фавиньяна ( Эгадские острова ), у раннемезолитических особей из сицилийской пещеры (провинция Трапани ) .
U2'3'4'7'8'9 был найден у образца Balma Guilanya (~13 тыс. л. н., Испания). Географическое распространение U2'3'4'7'8'9 соответствует раннему распространению человеческих групп в Западную Европу и, как предполагается, пережило LGM в Иберийских и Апеннинских рефугиумах. .
U4'9 определили у образца NEO283 (23,685 л. н. до н.э.) из карстового грота Котиас Клде , Грузия .
U4'9 был найден у итальянского образца Tagliente 2 (коммуна Греццана ) возрастом 16 980 — 16 510 л. н. ( эпиграветтская культура ) .
Гаплогруппа U2
С низкой частотой встречается по всей Европе и на Ближнем Востоке .
Субклады U2a, U2b и U2c являются автохтонными гаплогруппами в Индии. В наборе данных по Центральной Азии (Comas et al. 2004) из 232 линий только 2 принадлежат к U2a, 1 — к U2c .
Гаплогруппа U2 (U2* ) определена у образцов Kostenki 12(33—31,8 тыс. л. н.), Kostenki 14(38,7—36,3 тыс. л. н.), найденного на стоянке Маркина Гора Костёнковского комплекса стоянок в России .
U2 (субклад U2f2 ) определили у трёх образцов со стоянки Сунгирь Sunghir 2(35,3—32,2 тыс. л. н.), Sunghir 3(35,1—32,0 тыс. л. н.), Sunghir 4(34,5—33,5 тыс. л. н.) в России .
U2 определили у образцов Ostuni 2(29,3 - 28,6 тыс. лет) из Италии, Goyet Q53-1(28,2 - 27,7 тыс. лет), Goyet Q375-19(27,7 - 27,3 тыс. лет), Goyet Q56-16(26,6 - 26 тыс. лет) из ( фр. Grottes de Goyet ) в Бельгии. Граветтская культура (Граветт, Ла-Граветт, Лаграветтская, Граветянская) — археологическая культура кроманьонцев позднего палеолита.
U2e3 определили у охотницы-собирательницы I1960 (Tyumen_HG, 8166 л. н.) со стоянки Мергень 6 на озере Мергень (озеро) в Тюменской области ( Россия ) .
U2 обнаружена у представителей синташтинской археологической культуры.
Гаплогруппа U2b2, характерная для Южной Азии, определена у представительницы хараппской (индской) культуры из Ракхигархи (2800—2300 гг. до н. э.) .
U2e3a определили у представителя катакомбной культуры SA6003 (2474—2211 лет до н. э.) .
U2e2 и U2e2a1a2 определили представителей унетицкой культуры из Чехии (Czech_EBA_Unetice 3850—4100 л. н.) .
U2e1a1 и U2e2a1a2 определили у участников битвы на реке Толлензе , произошедшей более 3200 лет назад .
U2b2 определили у образцов I12142 и I12446 (1000—800 лет до н. э., Katelai) из долины Сват (Пакистан) .
Один мужчина хунну из Duurlig Nars в Северо-Восточной Монголии, живший 2000 лет назад, оказался обладателем митохондриальной гаплогруппы U2e1 .
Гаплогруппа U3
В Европе и Средней Азии распространение зафиксировано на уровне 1 % населения, на Ближнем Востоке — 2,5 %. Носителями U3 является около 6 % населения Кавказа (у сванов наличие U3 составляет 4,2 %) и от 36 % до 56 % у цыган Польши, Литвы и Испании .
U3a2a1 определили у неолитического образца cay011 (ок. 10 тыс. л. н.) из Чайоню (Анатолия) .
U3a определили у образца Ash033 (7870—7595 лет до н. э.) из Ашиклы-Хююка (Анатолия) .
U3a1 определили у одного из 9 убитых около 5300 года до н. э. неолитических фермеров, найденных пещере ( Бисаурри , Арагон, Пиренеи) .
U3 определили у представителей днепро-донецкой культуры .
U1a'c, U3b, U3b1a, U3b3, U4a1 определили у образцов из пещеры B3 (Cave B3) в долине Бакаа (Baq҅ah Valley) в 20 км к северо-западу от Аммана (бронзовый век Иордании, 1424—1288 лет до н. э.) .
U3b1b определили у образца VK555 (VIII век) из корабля (волость Сальме , Эстония) .
U3a1 определили у женского образца VK193 (X—XII века) из Гренландии .
U3a1c определили у образца VK125 (XII—XIII века) из Тронхейма .
U3a* определили у образца KRA004 из Кракауэр-Берга (Krakauer Berg) в Восточной Германии, датирующегося возрастом примерно 612 л. н. (666—558 л. н.) .
Гаплогруппа U4
Субклады U4: U4a , U4b , U4c и U4d .
Широко распространилась в Европе примерно 25 тыс. лет назад. Предполагается ближневосточное (анатолийское) происхождение субгаплогруппы U4 . Гаплогруппы U4a1 , HV3, HV4, возможно, могут быть прослежены до неолита в Восточной Европе и на Кавказе. Эти линии мтДНК, вероятно, были вовлечены в позднеледниковую экспансию из восточноевропейских рефугиумов (мест выживания) после LGM (ледниковых периодов), (примерно за 12 000–19 000 лет до настоящего времени ) . U4a2a, U4a2* определенно моложе и датируются между 6400 и 8200 YBP, что позволяет предположить, что их расширение может быть связано с более поздними историческими событиями. Наиболее важным компонентом с точки зрения этнической истории славян является митохондриальный субкластер U4a2, скорее всего, центрально-восточноевропейского происхождения. Расширение этого подкластера можно объяснить рассредоточением культуры шнуровой керамики , которая процветала 5200–4300 л. н. в Восточной и Центральной Европе. Эта культура (также известная как культура боевых топоров) охватывала большую часть континентальной Северной Европы от реки Волги на востоке до реки Рейн на западе .
U4c1 определена у одного из образцов мехтоидов из Тафоральта , Марокко (19 800 лет до н. э.) .
U4a1 определена у образца PES001 (10 785—10 626 лет до н. э., Y-хромосомная гаплогруппа R1a5-YP1301 (под R1a1b~-YP1272)) с могильника Песчаница 1 на озере Лача .
U4a2b и U4c1 выявлены у представителей иберо-мавританской культуры на территории Северной Африки (10 120—8550 лет до н. э.) .
U4b определили у образца I1733 (9000—7500 лет до н. э.), Y - R1a из Василевки-на-Днепре .
U4 определили у образца NEO497 (10081, no 7, grave 14) (8705 лет до н. э.), Y- R1a из Василевки-на-Днепре .
U4a NEO537, Y-R-Y13202, U4a1, U4a2, U4d NEO536, Y-R-P297 (R1b) определили у образцов (от 8671±48 до 4600 лет до н. э.) с мезолитической стоянки культуры Веретье у озера Кубенское в деревне Минино (Вологодская область) .
U4a1 определили у образцов MN2002(ок. 8595 лет до н. э.) Y-хромосомная гаплогруппа R1a1 M459, MNN005 (ок. 8563 лет до н. э.), MNN003 (ок. 5610 лет до н. э.), Y-хромосомная гаплогруппа CT у озера Кубенское в деревне Минино , Вологодская область , Россия
U4 определили у образцов BVG1 (8112 лет до н. э.) и BVG2 (8988 лет до н. э.), найденным в многослойном торфяном участке Береговая 2, расположенном примерно в 150 км к северо-западу от Екатеринбурга .
U4a1 определили у образца SF12(9033–8757 лет до н. э. ), U4a2 у образца SF9 (9300–8988 лет до н. э. ), могильник Stora Förvar, Stora Karlsö , у образца SBj, могильник Stora Bjers, Stenkyrka parish, остров в Балтийском море Готланд , Швеция (8963–8579 лет до н. э. ) .
U4a2 определили у образца Lepe51(7,940-7,571 лет до н. э.) Лепенски-Вир , Сербия .
U4 определили у образца NEO17 (X82) (ок.7197 лет до н. э.), Y-I-PF6915 (I2) в образце, найденном в затопленной лодке, Hummervikholmen, Vest-Agder, Норвегия .
U4 определили у представителей эпохи неолита в Украине у образца Ukraine_N1=I1378 (6,469-6,293 лет до н. э.) днепро-донецкой культуры .
U4a2 определили у образца Spiginas4 (6440–6230 лет до н. э) из Литвы, относят к кундской культуре .
U4, U4a1 определили у представителей эпохи неолита в Латвии у образца Latvia_MN2 (6179-5750 лет до н. э.) и у образца Latvia_MN1 (6201-5926 лет до н. э.). Относят к культуре ямочно-гребенчатой керамики .
U4 определили у представителей эпохи мезолита или неолита в Карелии Оленеостровский могильник (Онежское озеро) (ок.7500 лет до н. э.) .
U4a определили у образца NEO671(F1586) (6709 лет до н. э.), Y-хромосомная гаплогруппа R-V88 (R1b), Железные Ворота , Schela Cladovei, Румыния .
U4a2 или U4d определили у представителя хвалынской культуры , жившего 6700 лет назад .
U4d определили у образца NEO557(No 4, 8625, burial 11) (5900 лет до н. э.), Y-хромосомная гаплогруппа R-Y13202 (R1b), U4a1 у образца NEO556(No 3, 8623, burial 7) (5900 лет до н. э.) , Караваиха, Россия .
U5b1d1, U4a, U2e1 и архаичная Y-хромосомная гаплогруппа R1a5 (R1a1b~-YP1272 ) у представителей культуры гребенчатой керамики из местонахождения ( эст. Kudruküla , устье реки Нарва 5600 лет назад) .
U4a2a определили у образца NEO212(ок. 5,443 лет до н. э.), Y-хромосомная гаплогруппа I2a, NEO204(ок. 5,278 лет до н. э.), Y-хромосомная гаплогруппа I-P220 (I2a), Днепро-донецкая культура , Голубая Криница, Россия .
U4a определили у образца I2074 (5,602–5,376 лет до н. э.) и I0992 (5,002–4,730 лет до н. э.) могильник Корчуган 1, погр. 1, Васьково 4, Межгорная котловина между отрогами Алтая и Саян; Новосибирская область , Россия
U4d определили у образца NEO270(ок. 5,343 лет до н. э.), Y-хромосомная гаплогруппа I-Y24472 (I2a) эпохи неолита, Мамай гора, Украина .
U4a1 определили у образца 2017/01(5,300—5,050 л. н.) со стоянки Riņņukalns на реке Салаца на севере Латвии .
U4a2 определили у образца NEO814(ок. 5,165 лет до н. э.), эпохи мезолита, Bodal K, Зеландия , Дания .
U4a1 определили у образца NEO185 (ок. 5,078 лет до н. э.), Y-Q-YP761 представителя льяловской культуры ; U4b1b1 у образца NEO194(ок. 3,647 лет до н. э.), Y-R-M479 (R2) со стоянки Сахтыш IIA, Ивановская область , Россия .
U4a2 определили у образца NEO746 (ок. 5,036 лет до н. э.) Henriksholm-Bøgebakken (Vedbæk), Y - I-L596 Зеландия , Дания .
U4a1 определили у образца NEO78 (ок. 5,003 лет до н. э.), Y-Q-YP1669 (Q1a) эпохи неолита Омская стоянка 2, Омская область , Россия .
U4a образец NEO302(N134, Ukraine; n7)(ок. 4,889 лет до н. э.), I-L699 (I2a) могильник Волненский и NEO305(N133, Ukraine; n15)(ок. 4,661 лет до н. э.), I-L1443 могильник Васильевский, Днепро-донецкая культура , Украина .
U4a1 определили у представителя эпохи неолита (GE83 (KOP002)(5,200 - 4,850 лет до н. э.) на острове Эланд в Швеции .
U4 определили у представителей эпохи неолита (Ajv4, Fri24, Ire5, Ire4) на острове Готланд в Швеции (5000 - 4400 лет до н. э.) .
U4a1 определили у образца NEO498(ок. 5,106 лет до н. э.), Y-R1b, эпохи мезолита и у NEO508(ок. 4,354 лет до н. э.) эпохи неолита, Вовниги2, Украина .
U4a1 определили у образцов I5875 (5,291-5,060 лет до н. э.) и I5892 (5301-4982 лет до н. э.) могильник Дереивка, Украина .
U4a2 определили у образцa C3349 (5,043-4,019 лет до н. э.), U4b1a1a1 определили у образцa C3350 (2722-2426 лет до н. э.) могильник Songshugou уезда Зимунай в округе Алтай Синьцзян-Уйгурского автономного района Китая. .
U4d3 определили у представителя чемурчекской культуры , образец C1707 (4,814-4,450 лет до н. э.) могильник Bolati уезда Бурчун в округе Алтай Или-Казахского автономного округа Синьцзян-Уйгурского автономного района Китая. .
U4a1 определили у представителя чемурчекской культуры, образец C1703 (4,200 лет до н. э.) могильник Tuoganbai уезда Каба в округе Алтай Синьцзян-Уйгурского автономного района КНР .
U4a1 определили у образцов Zv162 (4470 ± 72 лет до н. э.) и Zv317 (3,890 ± 67 лет до н. э.) у представителя кундской культуры из могильника Звейниеки, на севере Латвии, на северном берегу озера Буртниекс в устье реки Руя. .
U4b1b2 определили у представительницы трипольской культуры этапа BI-II KYT-SSX (4,049-3,820 лет до н. э.) из урочища Коломийцев Яр близ Копачова (Обуховский район Киевской области) .
U4d1 определили у представителя унетицкой культуры из Чехии (Czech_EBA_Unetice 3,850—4,100 л. н.) .
U4a1 определили у образцов могильника Большой в Карелии ( Мурманская область) (ок.3,500 лет до н. э.) .
U4a определили у образца NEO83(ок. 3,716 лет до н. э.), R-L1432(R1b) эпохи неолита Боровянка 17, Омская область , Россия .
U4a1 определили у женщины с позднетрипольской стоянки Gordinești в Молдавии (3,500—3,100 лет до н. э.) .
U4c1 определили у образцов SIJ003(BZNK-279/1)(3,349-3,033 лет до н. э.), SIJ002(BZNK-278/1)(3,349-3,033 лет до н. э.) Y-L, SIJ001(BZNK-278/1)(3,329-3,022 лет до н. э.), Синюха могильник поздней майкопской культуры в Адыгее , Россия .
U4a2 определили у образцов VEK006, VEK007 Y-J2, VEK008 (3,000-2,800 лет до н. э.) Курганы «Харман-тепе», Куро-араксская культура , село Великент в Дербентском районе Дагестана .
U4a2 определили у образца MK3003(BZNK-051/1)(2,577-2,476 лет до н. э.) из курганного могильника «Маринская-3», курган 1, м. 7, U4d3, Y DNA - R1b1a2 у образца RK4002(BZNK-304/2)(2,831-2,489 лет до н. э.) из курганного могильника «Расшеватская-4», Катакомбная культурно-историческая общность , Россия
U4a2f определили у образца NEO121(MAN 15, A, 101)(ок. 2,577 лет до н. э.), Y- I-Z27038 (I2a) эпохи палеолита, пещера Grotte Mandrin, Rhone-Alpes, Франция .
U4a1 определили у образца IVA001 (2,864–2,496 лет до н. э.) с могильника Ивановогорский фатьяновской культуры , Россия .
U4d3 определили у представителя катакомбной культуры RK4002 (2,831—2,489 лет до н. э.) .

- 3000 г. до н. э.: первоначальная миграция на восток, положившая начало афанасьевской культуре , возможно, прототохарской .
- 2900 г. до н. э.: миграции на северо-запад, несущие культуру шнуровой керамики, превращаясь в культуру колоколовидных кубков, возможно, предок итало-кельтов (оспаривается).
- 2700 г. до н. э.: миграция шнуровиков на восток, переходящая в Фатьяново - Баланово (2800 г. до н. э.) → Абашево (2200 г. до н. э.) → Синташта (2100—1900 гг. до н. э.) → Андроново (1900—1700 гг. до н. э.) .
U4a2 определили у образцa C4255 (2,678-2,356 лет до н. э.) могильник Eminhuojierte уезда Дурбульджин в округе Чугучак Синьцзян-Уйгурского автономного района КНР .
U4 определили у представителей ямной культуры , андроновской культуры .
U4d1 определили у образца NEO51(MHM 4555, Grave 105)(ок. 2,391 лет до н. э.), Баальбергская культура , Kastanjegården, Skåne Швеция .
U4a2 определили у образцa C3319 (-), U4b1a4 определили у образцa C3313 (2,345-2,159 лет до н. э.) могильник Caishichang уезда Нилки в составе Или-Казахского автономного округа Синьцзян-Уйгурского автономного района КНР .
U4a1 определили у образца I0231= SVP3 в могильнике Екатериновка, Самарская область Ямная культура, (2,310 год до н. э.) .
U4b1a1a1 определили у образца NEO737(individ X, PMD 206)(ок. 2,161 лет до н. э.), Y- R-P310 (R1b) и U4c1 определили у образца NEO735(Individ III, PMD 200)(ок. 2,045 лет до н. э.), Y-R-P310 (R1b), эпоха среднего неолита B/позднего неолита, Borreby, Зеландия , Дания .
U4a2 определили у образца NEO76(IPDN 48-1, pit 3)(ок. 2,117 лет до н. э.) эпохи Неолита - Халколита, Остров2, Тюменская область, Россия .
U4a2 определили у образца NEO878(Kyndeløse XIII, not in PMD)(ок. 2,088 лет до н. э.) эпоха среднего неолита/позднего неолита, Kyndeløse, Зеландия , Дания .
U4a1a определили у образца NEO946(NM 271/45, AS 30/45, Reol: 33:12)(ок. 1,195 лет до н. э.), Q-F1213 эпоха среднего неолита A Hove Å, Зеландия , Дания .
U4b1a4 определили у образцa C387 (1,727-1,575* лет до н. э.), U4a2 определили у образцa C401 (1,727-1,575* лет до н. э.) могильник Hetian уезда Лоп в округе Хотан Синьцзян-Уйгурского автономного района КНР. .
U4a1a определили у образцов ARZ-T25, ARZ-T29 (700 ± 25 н. э.) в курганах Аржан-1 и Аржан-2 у представителей ( части уюкской культуры ) скифо-сибиряков железного века .
U4d2 определили представителя Bartym3/Bartim16/B (428—591 гг., Phase II) из Бартыма (Пермский край) и у образца с озера Уелги (Челябинская область) . Носителей считают представителями древних венгров, которые произошли из Уральского региона России и мигрировали через Среднее Поволжье и восточноевропейские степи в Карпатский бассейн в девятом веке нашей эры. .
U4a1 определили представителя пазырыкской культуры Ak-Alakha-5, mound 4, burial 2 (400-300лет до н. э.), Алтай, плоскогорье Укок, курганы в долине реки Акалаха .
U4, U4a3 определили у представителей тагарской культуры .
U4a1 и U4a2 встречается в современной популяции Центральной и Восточной Европы (русские, белорусы, украинцы, поляки и словаки) - в среднем 4,6%; Северо-Восточная Европы (финны, карелы, эстонцы, латыши, литовцы) - в среднем 2,5%; Волго-Уральского региона( марийцы , коми-зыряне, коми-пермяки, мордва, удмурты, чуваши , татары, башкиры) - в среднем 6,7%. Центральной и Западной Европы (британцы, немцы, чехи, австрийцы, швейцарцы, боснийцы, словенцы, итальянцы, французы, испанцы и португальцы)- в среднем 1,1% .
U4 более 3% встречается в некоторых современных популяциях Испании: Кантабрия - 3,8%; Каталония - 3,8%; Франции: Бирон (Атлантические Пиренеи) - 6,2%; Сарта и Мен и Луара - 4,4%; Атлантическая Луара - 4%; Великобритания: Корнуолл - 4,3%; в некоторых частях Германии до 6,3%; в Австрии - 4%; в Швейцарии - 3,9%
Гаплогруппа U4a1 характеризуется также западно-сибирским распространением, встречающимся с самой высокой частотой (7–21%) в таких популяциях, как кеты и U4a, U4c у народа нганасаны . В другом исследовании указано на распространение гаплогруппы U4 с беспрецедентно высокой частотой - 28,9% у кетов, 20,8% у энцев/нганасан и 16,3% у манси.
На Кавказе и Турции U4 встречается в Грузии (18%), в Армении (6,8%), в Турции (1%) .
Из современных народов за пределами Европы и Кавказа U4 встречается особенно в Иране (3%) и по всей Центральной Азии, особенно в Кыргызстане (3%), Туркменистане (3%), Узбекистане (2,5%) и Казахстане (2%), а также в части Сибири, особенно в Республике Алтай (5%) и среди носителей языков ханты и манси (12%). U4 также встречается с высокой частотой у некоторых этнических групп в Пакистане и Афганистане, в том числе у белуджей (2,5%), хунза бурушо (4,5%), хазарейцев (8%), парсов (13,5%) и особенно у калашей Гиндукуша (34%) .
Описание Ульрике (Ulrike)(U4) от Брайана Сайкса : Клан Ульрике по-немецки «Хозяйка всего») не входит в число первоначальных кланов «Семи дочерей Евы» (семь главных митохондриальных европейских родословных), но, имея в своем составе чуть менее 2% европейцев, он претендует на то, чтобы быть включенным в число численно важных кланов. Ульрике жила около 18 000 лет назад в холодных убежищах Украины на северных границах человеческого обитания. Хотя потомки Ульрике редко встречаются, клан сегодня встречается в основном на востоке и севере Европы с особенно высокой концентрацией в Скандинавии и странах Балтии.
Гаплогруппа U7
Во многих европейских популяциях гаплогруппа U7 отсутствует, но на Ближнем Востоке частота её распространения превышает 4 %, достигая 10 % среди жителей Ирана и вновь опускаясь до 5 % в Пакистане. На западе Индии к носителям этой гаплогруппы относится 12 % населения, но в остальных регионах страны — только 2 %. Родиной U7 считают Иран или западную Индию .
U7 определили у неолитического образца Tutkaul 1 (8419—8026 лет до настоящего времени , Y-хромосомная гаплогруппа Q1b2a, ANE) со стоянки Туткаул в Таджикистане .
Одна из женщин, похороненных в Осебергском корабле , имела гаплогруппу U7.
Гаплогруппа U8
- U8 определена у позднепалеолитического образца BK-1653 возрастом ок. 34,5 тыс. л. н. из пещеры Бачо Киро в Болгарии
- U8c обнаружена у палеолитического образца Vestonice 13 из Дольни-Вестонице (Чехия), жившего 31 тыс. лет назад
- U8c обнаружена у образца Sunghir 1 (33,9—31,8 тыс. л. н.) со стоянки Сунгирь
- U8c определили у представителя граветтской культуры Paglicci 12 из Италии
- U8a определили у представителей мадленской культуры
- U8a специфична для басков . Поскольку в Северной Африке отсутствует, считается, что она занесена из Западной Азии
- U8b1a2 и U8b1b определили у представителей трипольской культуры (3700—3500 лет до н. э.)
- U8a1 обнаружен у двух представителей унетицкой культуры
- U8b найдена в Италии и Иордании
- U8b1a2 обнаружен у двух представителей майкопской культуры
Гаплогруппа UK
Считается, что носители гаплогруппы UK мало восприимчивы к синдрому приобретённого иммунного дефицита .
См. также
| Митохондриальная Ева | ||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||
| L0 | L3 | |||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||
| M | N | |||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| D | E | G | Q | R | A | S | X | Y | N1 | N2 | ||||||||||||||||
| | | | | | | | | |||||||||||||||||||||||
| C | Z | B | F | R0 | P | I | N1a | W | ||||||||||||||||||
| | | | | | | ||||||||||||||||||||||||
| HV | JT | K | ||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| H | V | J | T | Устаревшие кластеры | ||||||||||||||||||||||
Примечания
- ↑ от 5 апреля 2008 на Wayback Machine National Geographic
- ↑ González A. et al. . Дата обращения: 5 января 2009. 18 февраля 2009 года.
- Raghavan M. et al. от 29 октября 2018 на Wayback Machine , 2014
- Elena Essel et al. от 7 мая 2023 на Wayback Machine // Nature, 03 May 2023 ( от 7 мая 2023 на Wayback Machine )
- . Дата обращения: 5 января 2009. 22 января 2010 года.
- J. Eldon Molto et al. от 3 ноября 2018 на Wayback Machine , 2017
- ↑ Sanni Översti et al. от 17 ноября 2019 на Wayback Machine , 15 November 2019
- Barbujani G, Bertorelle G. «Genetics and the population history of Europe» // Proceedings of the National Academy of Sciences , 2001.
- Alessandro Achilli et al. (2005) от 19 января 2020 на Wayback Machine
- Sanni Översti, Päivi Onkamo, Monika Stoljarova, Bruce Budowle, Antti Sajantila, Jukka U. Palo . , 2017
- ↑ Fu Q. et al. A revised timescale for human evolution based on ancient mitochondrial genomes, Current Biology, 21 March 2013.
- Gabriele Scorrano et al. от 11 декабря 2021 на Wayback Machine , December 08, 2021
- ↑ Qiaomei Fu et al. , 2016
- Jones, E. R. et al. от 21 августа 2016 на Wayback Machine , 2015
- Peter de Barros Damgaard et al. от 24 июня 2018 на Wayback Machine // Supplementary Materials for: The first horse herders and the impact of early Bronze Age steppe expansions into Asia, 29 Jun 2018
- Eppie R. Jones et al. // Current Biology, February 02, 2017
- от 5 сентября 2021 на Wayback Machine , June 2018
- ↑ Alessandra Modi et al. от 5 сентября 2021 на Wayback Machine // Annals of Human Biology. Volume 48, 2021 - Issue 3
- ↑ Iain Mathieson et al. от 6 июня 2020 на Wayback Machine , 2017
- Jamie Hodgkins et al. от 20 декабря 2021 на Wayback Machine // Scientific Reports volume 11, Article number: 23735, 14 December 2021
- Natalija Kashuba et al. от 9 января 2020 на Wayback Machine , 2019
- . Дата обращения: 30 июня 2012. 2 июля 2012 года.
- Jens Blöcher . // Dissertation, Mainz, Johannes Gutenberg-Universität, 2019
- Zuzana Hofmanová et al. , June 28, 2022
- Iosif Lazaridis et al. от 12 февраля 2021 на Wayback Machine , 2013 ( от 16 марта 2021 на Wayback Machine )
- . Дата обращения: 9 февраля 2020. 4 марта 2016 года.
- . Дата обращения: 24 октября 2014. 25 мая 2018 года.
- Federico Sánchez-Quinto et al. от 16 мая 2019 на Wayback Machine , 2012 Aug 21
- ↑ Vagheesh M. Narasimhan et al. (англ.) // Science. — 2019-09-06. — Vol. 365 , iss. 6457 . — ISSN . — doi : . 4 апреля 2021 года. ( от 17 марта 2021 на Wayback Machine )
- Olalde, I., Brace, S., Allentoft, M. et al. . Nature (21 февраля 2018). Дата обращения: 18 марта 2021. 14 февраля 2021 года.
- Iain Mathieson et al. от 3 марта 2016 на Wayback Machine , 2015
- Inigo Olalde et al. от 5 сентября 2015 на Wayback Machine , 2015
- Maïté Rivollat et al. , April 21, 2022
- Pere Gelabert et al. от 5 января 2022 на Wayback Machine , November 2021
- ↑ Chuan-Chao Wang et al. от 15 июля 2021 на Wayback Machine , 04 February 2019 (Supplementary Data 1)
- ↑ Allentoft ME et al. (англ.) // Europe PMC. — 2015. — doi : . 9 мая 2020 года.
- Nikitin, A. G. , 2012
- Julian Susat et al. от 2 июля 2021 на Wayback Machine // Cell. Volume 35, ISSUE 13, 109278, June 29, 2021
- ↑ Nick Patterson et al. от 1 января 2022 на Wayback Machine // Nature, 22 December 2021
- Odile Loreille et al. от 21 апреля 2021 на Wayback Machine , Genes 2018, 9(3), 135; doi:10.3390/genes9030135
- , 03 December 2022
- Elizabeth A. Matisoo-Smith et al. от 8 марта 2020 на Wayback Machine , 2016
- . Дата обращения: 26 мая 2016. 3 июня 2016 года.
- Linea Melchior, Toomas Kivisild , Niels Lynnerup, Jørgen Dissing . от 10 апреля 2022 на Wayback Machine , May 28, 2008
- ↑ Ashot Margaryan et al. от 26 марта 2021 на Wayback Machine , 2020 ( от 12 февраля 2020 на Wayback Machine )
- Marc Haber et al. от 31 мая 2019 на Wayback Machine , 2019
- Мустафин Х. Х., Энговатова А. В., Альборова И. Э., Тарасова А. А. от 7 марта 2022 на Wayback Machine // Археология Подмосковья. Материалы научного семинара. Выпуск 18. М.: Институт археологии РАН, 2022. С. 111
- Chao Ning, Shizhu Gao, Boping Deng, Hongxiang Zheng, Dong Wei. (англ.) // Journal of Human Genetics. — 2015. — Vol. 61 , iss. 2 . — P. 103–108 . — ISSN . — doi : . 12 мая 2020 года.
- Min-Sheng Peng, Weifang Xu, Jiao-Jiao Song, Xing Chen, Xierzhatijiang Sulaiman. // European Journal of Human Genetics. — 2018-1. — Т. 26 , вып. 1 . — С. 124–136 . — ISSN . — doi : . 8 мая 2020 года.
- Maja Krzewińska et al. , 25 January 2024
- Maja Krzewińska et al. от 31 марта 2022 на Wayback Machine , 2021
- ↑ Дата обращения: 3 октября 2017. 5 марта 2016 года.
- Erwan Pennarun et al. от 20 сентября 2015 на Wayback Machine , 2012
- ↑ Maca-Mayer N. // BMC Genetics, 2003
- Александр Марков . Эволюция человека: обезьяны, кости и гены. 2011
- Olivieri A., Achilli A., Pala M., Battaglia V., Fornarino S.. et al. (2006) The mtDNA Legacy of the Levantine Early Upper Paleolithic in Africa. Science 314: 1767
- Adimoolam Chandrasekar et al. от 31 августа 2021 на Wayback Machine , October 20
- от 18 марта 2018 на Wayback Machine // Loosdrecht el al. Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations, 2018
- от 15 мая 2021 на Wayback Machine , 02.06.2016
- Iosif Lazaridis et al. от 22 сентября 2018 на Wayback Machine , 2018
- M. van de Loosdrecht el al. , Science (2018)
- . Дата обращения: 18 марта 2018. 17 марта 2018 года.
- Éadaoin Harney et al. от 20 августа 2018 на Wayback Machine , 2018
- Marina Silva et al. от 25 сентября 2021 на Wayback Machine , 13 September 2021
- Martin Sikora et al. от 24 октября 2018 на Wayback Machine
- Giulio Catalano от 5 марта 2021 на Wayback Machine , 2019
- Alessandra Modi et al. от 1 ноября 2020 на Wayback Machine , 2020
- Villalba-Mouco, V., van de Loosdrecht, M.S., Rohrlach, A.B. et al. , 2022
- ↑ Morten E. Allentoft et al. от 26 мая 2022 на Wayback Machine , 2022
- Eugenio Bortolini et al. , April 21, 2021 ( от 22 апреля 2021 на Wayback Machine , August 10, 2020)
- [www.familytreedna.com/public/mtDNA%20U2 FTDNA mtDNA U2 Haplogroup project.]
- Malliya gounder Palanichamy et al. от 5 мая 2021 на Wayback Machine , December 2004
- от 1 февраля 2010 на Wayback Machine
- от 1 октября 2022 на Wayback Machine // YFull MTree 1.02.16767.
- ↑ Sikora M. et al. от 22 декабря 2018 на Wayback Machine , Science 10.1126/science.aao1807 (2017).
- Vasant Shinde et al. от 17 сентября 2019 на Wayback Machine , 2019
- Joachim Burger, Vivian Link . от 30 октября 2020 на Wayback Machine // Current Biology, 9 Apr 2020
- . Дата обращения: 13 сентября 2020. 7 июня 2022 года.
- Vagheesh M Narasimhan et al. от 1 апреля 2018 на Wayback Machine , March 31, 2018
- Vagheesh M. Narasimhan et al. от 4 апреля 2021 на Wayback Machine , 06 Sep 2019
- . Дата обращения: 19 января 2015. 27 декабря 2014 года.
- . Дата обращения: 5 января 2009. 4 марта 2009 года.
- [www.blackwell-synergy.com/doi/pdf/10.1111/j.1529-8817.2005.00222.x Blackwell Synergy — Cookie Absent] (недоступная ссылка)
- N. Ezgi Altınışık et al. от 15 ноября 2022 на Wayback Machine , 9 Nov 2022 ( , February 01, 2022)
- Reyhan Yaka et al. , April 14, 2021
- Kurt W. Alt et al. от 7 февраля 2020 на Wayback Machine , 07 February 2020
- Newton J. (2011) от 31 декабря 2019 на Wayback Machine : The Presence of East Eurasian Haplogroups Provides Evidence of Interactions with South Siberians Across the Central Asian Steppe Belt
- Lily Agranat-Tamir et al. от 8 августа 2021 на Wayback Machine , May 28, 2020 (Table S1. Overview of the 73 Individuals Newly Reported in this Study, Related to Figure 1)
- Cody Parker et al. от 22 декабря 2021 на Wayback Machine , 26 October 2020 ( от 22 апреля 2021 на Wayback Machine , May 22, 2020)
- (2011).
- (2011). Дата обращения: 6 декабря 2022. 20 июля 2013 года.
- (2011). Дата обращения: 6 декабря 2022. 17 августа 2015 года.
- (2010). Дата обращения: 6 декабря 2022. 17 августа 2015 года.
- Boris Malyarchuk. // Russian Journal of Genetics 40(11):1281-1287. — 2004-01. 2 декабря 2022 года.
- ↑ Kristiina Tambets et al. [ The Topology of the Maternal Lineages of the Anatolian and Trans-Caucasus Populations and the Peopling of Europe: Some Preliminary Considerations]. — 2000-01. 2 декабря 2022 года.
- ↑ B. Malyarchuk et al. // Molecular Biology and Evolution, Volume 25, Issue 8, August 2008, Pages 1651–1658,. — 2008-08. 10 ноября 2022 года.
- Дата обращения: 30 декабря 2016. 20 октября 2019 года.
- ↑ Lehti Saag, Sergey V. Vasilyev, Svetlana V. Oshibkina et al. от 23 января 2021 на Wayback Machine (Table 1), July 03, 2020 ( от 30 января 2021 на Wayback Machine )
- Rym Kefi et al. от 20 октября 2019 на Wayback Machine , 2016
- ↑ Jens Blöcher . от 3 января 2021 на Wayback Machine , 2019
- Posth, C., Yu, H., Ghalichi, A; et al. , Nature, Published Online: March 22, 2023
- . Дата обращения: 28 января 2023. Архивировано 28 января 2023 года.
- Mikhail Zhilin al. // Cambridge University Press. — 2018-24-04. 14 сентября 2019 года.
- Günther T, Malmström H, Svensson EM, Omrak A, Sánchez-Quinto F, Kılınç GM et al. от 11 июня 2023 на Wayback Machine , PLOS Biology, Published Online: January 16, 2018
- Hofmanová, Z. et al.
- ↑ Eppie R. Jones et al. , Current Biology, Published Online: February 02, 2017
- Alissa Mittnik et al. от 25 апреля 2017 на Wayback Machine , 03.03.2017
- Alissa Mittniket al. // Nat Commun.. — 2018-30-01. 31 мая 2022 года.
- ↑ Clio Der Sarkissian et al. от 15 июня 2015 на Wayback Machine , 2013
- . Дата обращения: 11 октября 2015. 3 марта 2016 года.
- Lehti Saag et al. от 15 апреля 2017 на Wayback Machine , March 2, 2017
- ↑ Stanislav V. Dryomov . Stanislav V. Dryomov. // PLoS One. — 2021-01-28.
- Susat; et al. (2021). . . 35 (13). doi : .
- ↑ Helena Malmström et al. // Philos Trans R Soc Lond B Biol Sci.. — 2015-19-017. 9 ноября 2022 года.
- Mathieson et al. 2018 // Nature. — 2018-03-08. 24 июля 2023 года.
- ↑ Vikas Kumar et al. от 18 апреля 2022 на Wayback Machine , SCIENCE, Published Online: March 31, 2022
- Alexey G. Nikitin et al. , 2022
- Alexander Immel et al. от 19 марта 2022 на Wayback Machine , 2020 ( от 8 декабря 2019 на Wayback Machine )
- на Wayback Machine
- Choongwon Jeong et al. от 3 ноября 2020 на Wayback Machine , 2020
-
Mary, Laura (March 28, 2019). "Genetic kinship and admixture in Iron Age Scytho-Siberians".
.
138
(4): 411—423.
doi
:
.
PMID
.
S2CID
.
The absence of R1b lineages in the Scytho-Siberian individuals tested so far and their presence in the North Pontic Scythians suggest that these 2 groups had a completely different paternal lineage makeup with nearly no gene flow from male carriers between them
- Veronika Csáky et al. от 12 мая 2021 на Wayback Machine // Scientific Report, 22 January 2020
- Veronika Csáky al. // nature.com. — 2020-05-11. 12 апреля 2021 года.
- Gubina et al. Gubina, M.A., Kulikov, I.V., Babenko, V.N. et al. // Russian Journal of Genetics. — 2016-02-02.
- Pilipenko et al. Pilipenko AS, Trapezov RO, Cherdantsev SV, Babenko VN, Nesterova MS, Pozdnyakov DV, et al. // PLOS ONE. — 2018-09-20.
- O García and R Fregel et al. // Heredity volume 106, pages37–45 (2011). — 2010-04-21. 6 января 2023 года.
- Natalia V Volodko, Elena B Starikovskaya, Ilya Mazunin, N. P. Eltsov. // The American Journal of Human Genetics 82, 1084–1100, May 2008. — 2008-05. 2 декабря 2022 года.
- Дербенева О. А. от 18 мая 2019 на Wayback Machine , 2002
- Lluís Quintana-Murci et al. . — 2004-05. 9 ноября 2022 года.
- Qasim Ayub, Massimo Mezzavilla, Luca Pagani, Marc Haber, Aisha Mohyuddin. // American Journal of Human Genetics. — 2015-05-07. — Т. 96 , вып. 5 . — С. 775–783 . — ISSN . — doi : . 8 марта 2021 года.
- Olga A. Derbeneva, Elena B. Starikovskaya, Douglas C. Wallace, and Rem I. Sukernik. . — 2002-04. 9 ноября 2022 года.
- Брайан Сайкс , «Кровь Островов» (Bantam, 2006) страницы 118—119
- . Дата обращения: 5 января 2009. 11 июня 2011 года.
- Cosimo Posth et al. от 4 марта 2023 на Wayback Machine // Nature, 01 March 2023
- Jean-Jacques Hublin et al. от 19 июня 2020 на Wayback Machine , 11 May 2020
- Alexey G. Nikitin, Inna Potekhina, Nadin Rohland, Swapan Mallick, David Reich, Malcolm Lillie. Mitochondrial DNA analysis of eneolithic trypillians from Ukraine reveals neolithic farming genetic roots (англ.) // PLoS ONE. — 2017. — 24 February ( no. 12(2) ). — doi : .
- Brandt G. et al. (2013), Ancient DNA Reveals Key Stages in the Formation of Central European Mitochondrial Genetic Diversity, Science, vol. 342, no. 6155 (2013), pp. 257—261.
- Sokolov A. S. et al. от 11 мая 2017 на Wayback Machine
| Ключевые понятия |

|
|
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
|
Формирование славянских народов — генофонд, история
|
|||||||
|---|---|---|---|---|---|---|---|
| Основное | |||||||
| Генофонд славян |
|
||||||
|
Древнеславянские
археологические культуры |
|
||||||
Ссылки
- YFull MTree 1.02.00 (under construction)
- , from National Geographic
- (U5)
- [www.familytreedna.com/public/india The India(subcontinent)Genealogical DNA Project]
- (недоступная ссылка)
- [www.familytreedna.com/public/mtDNA%20U7 The U7 Haplogroup Mitochondrial DNA Project]
- (недоступная ссылка)
| Ключевые понятия |
|
|
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
- 2020-10-05
- 1