Interested Article - Рибулозобисфосфаткарбоксилаза

- 2020-01-25
- 2
Рибулозобисфосфаткарбоксилаза (Рибулозобисфосфаткарбоксилаза/оксигеназа, рибулёзобифосфаткарбоксилаза/оксигеназа, англ. Ribulose-1,5-bisphosphate carboxylase/oxygenase, RuBisCO ), рубиско — фермент ( КФ ), катализирующий присоединение углекислого газа к рибулозо-1,5-бисфосфату на первой стадии цикла Кальвина , а также реакцию окисления рибулозобифосфата на первой стадии процесса фотодыхания . Является одним из важнейших ферментов в природе, поскольку играет центральную роль в основном механизме поступления неорганического углерода в биологический круговорот. Рибулозобисфосфаткарбоксилаза является основным ферментом листьев растений и поэтому считается наиболее распространённым ферментом на Земле .
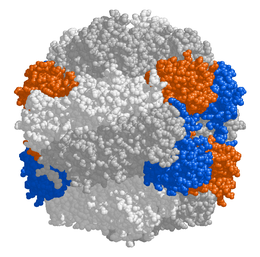
Структура
У растений , цианобактерий и хемоавтотрофных протеобактерий фермент обычно состоит из двух типов белковых субъединиц:
- Большая цепь ( L , массой 55 000 Да )
- Малая цепь ( S , массой 13 000 Да)
Активный центр фермента находится на больших цепях, объединённых в димеры. Все 8 димеров больших цепей и 8 малых цепи объединены в единый комплекс массой 540 000 Да. У некоторых протеобактерий малых цепей в составе рубиско не обнаружено. При этом большие цепи кодируются в ДНК хлоропласта , а малые — в ядре и транспортируются в хлоропласт перед сборкой всего белка .
Для работы фермента необходимы ионы Mg 2+ , которые размещаются в активном центре и способствуют присоединению CO 2 к остатку лизина , в ходе чего образуется карбамат . Образование карбамата протекает легче в щелочной среде: роль pH и ионов магния в регуляции работы фермента описано ниже.
Каталитическое действие
Субстратами для рибулозобифосфаткарбоксилазы являются рибулозо-1,5-бисфосфат , углекислый газ и вода, вместо углекислого газа может быть метаболизирован молекулярный кислород.
- В случае углекислого газа первым промежуточным продуктом выступает нестабильный шестиуглеродный фосфорилированный сахар, который сразу же распадается на две молекулы 3-фосфоглицериновой кислоты ( 3-фосфоглицерата ). Фосфоглицерат используется для синтеза глюкозы .
- В случае кислорода продуктами реакции являются 3-фосфоглицерат и фосфат гликолевой кислоты (ФГК). ФГК растения далее используют в процессе фотодыхания .
Реакция, проводимая ферментом, протекает относительно медленно (используются лишь несколько молекул углекислого газа в секунду) и является лимитирующей стадией всего цикла Кальвина. Константа Михаэлиса для карбоксилазной реакции рибулозобисфосфаткарбоксилазы равна 10±4 мкМ CO 2 , для оксигеназной 0,5 мМ O 2 , для рибулозобифосфата 1,5±0,5 мкМ.
Регуляция
У высших растений и некоторых водорослей фермент, активирующий рибулозобисфосфаткарбоксилазу (активаза) , необходим для формирования карбамата на активном центре. Активаза необходима, чтобы образованием карбамата уменьшить связь между рибулозобисфосфатом и активным центром и облегчить высвобождение продукта.
2-карбокси-D-арабитинол-1-фосфат связывается с активным центром рибулозобисфосфаткарбоксилазы и является её ингибитором. На свету фермент, активирующий рибулозобисфосфаткарбоксилазу вызывает диссоциацию ингибитора от активного центра. В диссоциированном состоянии ингибитор инактивируется CA1P-фосфатазой.
Фермент, активирующий рибулозобисфосфаткарбоксилазу, требует энергии АТФ и ингибируется при повышении концентрации АДФ . Активность рибулозобисфосфаткарбоксилазы косвенно регулируется соотношением концентрацией АТФ/AДФ.
Изменение конформации и снижение активности рибулозобисфосфаткарбоксилазы также наблюдется при наличии фосфат -аниона.
Сильно зависит активность фермента от концентрации CO 2 . Растения с C4-фотосинтезом и CAM-фотосинтезом выработали механизмы её увеличения и активации рибулозобисфосфаткарбоксилазы.
Генная инженерия
Оксигеназная активность рибулозобисфосфаткарбоксилазы приводит к потерям углерода из цикла Кальвина в ходе фотодыхания и снижает эффективность фотосинтеза . В этой связи проводились неоднократные попытки модифицировать гены, кодирующие синтез фермента, с тем, чтобы увеличить карбоксилазную и сократить оксигеназную активности. Одним из наиболее перспективных направлений здесь представляется трансплантация генов из красной водоросли Galdieria partita , рибулозобисфосфаткарбоксилаза которой обладает естественной высокой специфичностью к CO 2 , в высшие культурные растения. Это, предположительно, может увеличить их урожайность . В качестве донора генов в ряде исследований также выступает пурпурная бактерия Rhodospirillum rubrum .
Примечания
-
Cooper, Geoffrey M.
10.The Chloroplast Genome
//
. — 2nd. — Washington, D.C:
, 2000. —
ISBN 0-87893-106-6
.
. — «one of the subunits of ribulose bisphosphate carboxylase (rubisco) is encoded by chloroplast DNA. Rubisco is the critical enzyme that catalyzes the addition of CO2 to ribulose-1,5-bisphosphate during the Calvin cycle (see Figure 2.39). Not only is it the major protein component of the chloroplast stroma, but it is also thought to be the single most abundant protein on Earth, so it is noteworthy that one of its subunits is encoded by the chloroplast genome.».
(given that plants make up greater than 99% of the biomass on Earth.)
Dhingra A., Portis A.R., Daniell H. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2004. — April ( vol. 101 , no. 16 ). — P. 6315—6320 . — doi : . — . — PMC . . — «(Rubisco) is the most prevalent enzyme on this planet, accounting for 30–50% of total soluble protein in the chloroplast;». - См. Dhingra A, Portis AR Jr, Daniell H. Enhanced translation of a chloroplast-expressed RbcS gene restores small subunit levels and photosynthesis in nuclear RbcS antisense plants. 2004 Apr 5
- Harvey Lodish, Arnold Berk, S. Lawrence Zipursky, Paul Matsudaira, David Baltimore and James E. Darnell Molecular Cell Biology, 4th edition, Published by W. H. Freeman & Co. (2000) New York. в книге показывает структуру активного центра и место магния в нём
- A. R. Portis Rubisco activase—Rubisco’s catalytic chaperone // Jr in Photosynthesis Research (2003), volume 75, pages 11-27.
- Whitney S. M. , Andrews T. J. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2001. — Vol. 98, no. 25 . — P. 14738—14743. — doi : . — . John Andrews T. , Whitney S. M. (англ.) // Archives of biochemistry and biophysics. — 2003. — Vol. 414, no. 2 . — P. 159—169. — . Spreitzer R. J. , Salvucci M. E. (англ.) // Annual review of plant biology. — 2002. — Vol. 53. — P. 449—475. — doi : . — . Parry M. A. , Andralojc P. J. , Mitchell R. A. , Madgwick P. J. , Keys A. J. (англ.) // Journal of experimental botany. — 2003. — Vol. 54, no. 386 . — P. 1321—1333. — .
|
|
|
|---|
- 2020-01-25
- 2