Interested Article - IRES

- 2020-08-29
- 1
IRES ( англ. Internal Ribosome Entry Site — участок внутренней посадки рибосомы ) — регуляторный участок мРНК эукариот и их вирусов , который обеспечивает кэп -независимую, или внутреннюю инициацию трансляции . При таком механизме инициации рибосома связывается с мРНК непосредственно в районе IRES, которые чаще всего располагаются в 5'-нетранслируемой области (5'-НТО) недалеко от сайта инициации трансляции , минуя стадии узнавания кэпа и сканирования .
К 2016 году описано более 80 клеточных (у дрожжей , растений и других высших эукариот ) и 56 вирусных IRES. IRES были обнаружены у представителей следующих семейств вирусов: пикорнавирусы , флавивирусы , и лентивирусы . Кроме того, недавно была показана возможность трансляции, зависимой от IRES вируса эукариот, в клетках бактерий .
История
Участки внутренней посадки рибосомы были открыты в мРНК полиовируса и в 1988 году группами и соответственно. Они обнаружили, что в молекуле РНК есть участки, способные связывать рибосомы, тем самым инициируя трансляцию. Оказалось, что если поместить IRES между двумя репортёрными генами в мРНК, то второй (3'-концевой) цистрон также будет экспрессироваться . Однако использование данного метода (т. н. метода бицистронных конструкций) может привести к ряду артефактов, связанных со сплайсингом и потенциальной промоторной активностью участков внутренней посадки рибосомы .
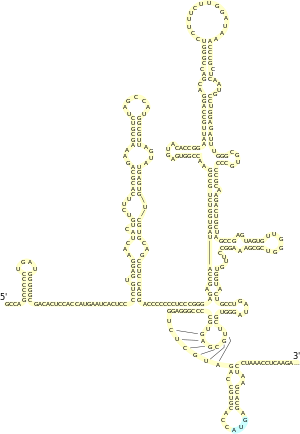
Структура

Для всех IRES пикорнавирусов характерно наличие мотива GRNA в центральном домене, который обеспечивает формирование четырёхпетельной конформации , а также небольшого пиримидинобогащённого участка, который расположен на 20—25 нуклеотидов выше кодона AUG . IRES флавивирусов содержит 4 домена, обозначаемых I—IV, причём у IRES вируса гепатита C (HCV), типового представителя IRES данной группы вирусов, последовательности, необходимые для активности IRES, находятся между доменами II, III и IV и захватывают первые 30 нуклеотидов открытой рамки считывания , начинающейся со старт-кодона AUG. Вторичная структура IRES HCV была тщательно изучена. В частности, установлено, что домен II представляет собой шпильку длиной 75 нуклеотидов с тремя внутренними и одной терминальной петлёй и что домен III обладает наиболее сложной вторичной структурой , которая включает несколько спиралей и шпилек. Вторичная и третичная структура IRES дицистровирусов довольно консервативна и включает 3 различных домена, каждый из которых содержит псевдоузел . IRES были выявлены почти у всех членов семейства лентивирусов (в число которых входит и вирус иммунодефицита человека ), причём IRES у них располагаются не только в 5'-НТО, но и в кодирующей области , где они участвуют в экспрессии некоторых изоформ основного структурного полипротеина .
Структуры эукариотических IRES очень разнообразны, и в них не выявлено никаких консервативных последовательностей и мотивов . В некоторых случаях для эффективной работы IRES должны быть стабильными структурами в мРНК, а в других, напротив, слишком жёсткая структура IRES ингибирует инициацию трансляции. Было высказано предположение, что IRES не являются жёстко зафиксированными структурами и способны к перестройкам, регулирующим их активность. IRES могут также обусловливать образование различных изоформ белка, тем самым дополнительно расширяя число возможных белковых продуктов, получаемых с одного гена .
Механизм
IRES обнаружены в 5'-нетранслируемых областях геномных мРНК вирусов и позволяют им транслироваться независимо от кэпа. Ряд клеточных мРНК также содержат IRES .
Клеточные IRES
В клетках IRES отвечают за посадку рибосом как на кэпированные, так и на некэпированные транскрипты в тех случаях, когда кэпзависимая инициация трансляции ингибирована (при стрессе , на определённой стадии клеточного цикла или при апоптозе ), и тем самым обеспечивают непрерывный синтез необходимых белков . Ряд генов, мРНК которых содержат IRES — c-Myc , APAF1 , Bcl-2 — при нормальных условиях экспрессируются мало, но в определённых ситуациях их экспрессия может существенно возрастать за счёт IRES-зависимой трансляции. Считается, что IRES могут также участвовать в поддержании низкого уровня трансляции ряда мРНК при нормальных условиях, связывая рибосомы и не позволяя им присоединяться к основным сайтам инициации. Такой механизм внутренней инициации в настоящее время плохо понятен, однако совершенно ясно, что эффективность IRES сильно зависит от транс-регуляторных белковых факторов, которые обеспечивают тонкую регуляцию IRES-зависимой трансляции на уровне отдельных клеток .
Структуры в 5'-НТО могут влиять на активность IRES, причём это влияние может быть опосредовано взаимодействиями как с различными транс-регуляторными факторами, так и непосредственно с рибосомами. Примерами генов, IRES которых находятся под контролем транс-регуляторных белков, являются протоонкогены Myc , участвующие в регуляции пролиферации клеток . Для IRES-зависимой трансляции этих мРНК необходимы особые факторы трансляции ITAF ( англ. IRES trans-acting factors ), которые выступают как шапероны РНК, заставляющие её принять правильную конформацию, подходящую для связывания с 40S субъединицей рибосомы .
Наличие IRES между AUG и другими старт-кодонами (отличными от AUG) свидетельствует о возможной роли IRES в инициации трансляции со слабых нестандартных старт-кодонов. IRES также могут взаимодействовать с короткими рамками считывания ( uORF ). Для доказательства существования IRES в структуре РНК недостаточно анализа её последовательности — все предположения должны быть подтверждены экспериментальными данными .
( англ. X-linked inhibitor of apoptosis, XIAP ) играет важную роль в регуляции апоптоза , поэтому его синтез должен точно контролироваться. 5’-НТО мРНК XIAP составляет 1700 нуклеотидов в длину и содержит IRES, облегчающий синтез XIAP в условиях стресса. Кроме описанной мРНК XIAP , в результате альтернативного сплайсинга образуется мРНК с более короткой 5'-НТО длиной 323 нуклеотида и не содержащая IRES. Количество мРНК XIAP с короткой 5'-НТО в клетках в 10 раз в больше, чем с длинной. Установлено, что мРНК с короткой 5’-НТО ответственна за синтез XIAP в нормальных условиях и транслируется по кэп-зависимому механизму, а мРНК с длинной 5’-НТО обеспечивает синтез XIAP в условиях стресса. Итак, комбинация альтернативных 5’-НТО и механизмов инициации трансляции обеспечивает постоянную экспрессию XIAP в любых условиях .
Другим примером контроля экспрессии генов через различные элементы 5’-НТО, является ген человеческого фактора роста фибробластов 2 ( англ. fibroblast growth factor 2, FGF-2 ). FGF-2 экспрессируется в 5 различных изоформах, образующихся при использовании альтернативных инициаторных кодонов в 5’-НТО, и трансляция его мРНК может идти не только кэпзависимо, но и IRES-зависимо. Интересно, что трансляция с четырёх из пяти инициаторных кодонов опосредована IRES. Предполагается, что IRES облегчает трансляцию с каждого из этих четырёх кодонов через модуляцию структуры мРНК при помощи транс-активирующих факторов .
Вирусные IRES

У многих РНК-содержащих вирусов инициация трансляции происходит по кэпнезависимому механизму при участии IRES, локализованных в 5'-НТО . Например, так происходит у ВИЧ, вирусов гепатита A и С . Такой механизм инициации трансляции удобен тем, что в его случае нет необходимости в сборке преинициаторного белкового комплекса, и вирус может быстро размножаться .
Некоторые вирусные мРНК имеют на 5'-конце ковалентно связанный белок ( ), поэтому использование кэпзависимой инициации для них исключено. IRES вируса гепатита C образует комплекс с 40S субъединицей рибосомы и привлекает фактор трансляции eIF3 клетки-хозяина . IRES многих пикорнавирусов не связывают 40S напрямую, а делают это через фактор инициации eIF4G, высокоаффинный сайт связывания которого находится в IRES . У многих вирусов (например, пикорнавирусов) для IRES-зависимой инициации одних канонических факторов инициации трансляции недостаточно: им также необходимы особые факторы, называемые ITAF . В зависимости от особенностей вторичной структуры и потребности в тех или иных факторах трансляции IRES делят на различные классы внутри отдельных семейств вирусов . Например, IRES пикорнавирусов делят на 4 класса (I—IV) .
Вирусный РНК - геном может содержать больше одного IRES. Например, , входящий в семейство , имеет РНК-геном с двумя IRES, один из которых локализован в 5'-НТО, а другой в . Оказалось, что функционирование этих элементов отличается: IRES, расположенный в 5'-НТО, инициирует трансляцию на всех этапах вирусной инфекции , а IRES, локализованный в межгенном участке, большую часть времени подавлен и активируется только через 2—3 часа после заражения. Временное разобщение двух IRES — эффективная вирусная стратегия для экспрессии определённых белков в строго определённую фазу инфекции .
Клиническое значение
Поскольку IRES играют важнейшую роль в регуляции ряда генов, мутации , затрагивающие IRES, приводят к развитию тех или иных заболеваний. В частности, к числу таких заболеваний у человека относят Х -сцепленную болезнь Шарко — Мари — Тута , множественную миелому и синдром ломкой Х-хромосомы .
Вирусные IRES могут выступать мишенями многих противовирусных препаратов . В настоящий момент разрабатываются препараты, которые непосредственно разрушают структуру IRES или же мешают взаимодействию IRES с рибосомой или белковыми факторами, например, факторами инициации трансляции. В частности, IRES вируса гепатита C, ввиду своей консервативности среди встречающихся в клинике образцов, может быть мишенью лекарств, блокирующих трансляцию у этого вируса . Среди препаратов, направленных против IRES, есть антисмысловые олигонуклеотиды , пептидо-нуклеиновые кислоты (RNA), (LNA), морфолиновые олигонуклеотиды , короткие шпилечные РНК , аптамеры РНК , рибозимы , , пептиды и малые молекулы . Показано, что ингибитор AZD1480 способен подавлять размножение вируса гепатита A, блокируя его IRES-зависимую трансляцию . Апигенин — препарат, применяющийся при лечении ящура — подавляет развитие вирусной инфекции, нарушая IRES-зависимую трансляцию у вируса ящура . Жизненный цикл энтеровирусов эффективно подавляет противоопухолевый препарат идарубицин , связывающийся с IRES .
Особый интерес представляют , жизненный цикл которых зависит от IRES, в связи с их возможным использованием в противораковой терапии .
IRES нашли широкое применение в создании векторов для генной терапии . Если локализовать IRES в 5'-НТО мРНК необходимых генов , то можно добиться их скоординированной экспрессии. В таких случаях чаще всего используется IRES вируса .
Методы
Для проверки того, обладает ли данная последовательность РНК свойствами IRES, можно использовать следующий метод. Создаётся бицистронная эукариотическая мРНК, в которой исследуемая последовательность находится между двумя репортёрными открытыми рамками считывания . Открытая рамка, находящаяся до исследуемой последовательности, будет транслироваться в белок по кэпзависимому механизму. Если изучаемая последовательность обладает свойствами IRES, то в клетке будет накапливаться и белок, соответствующий второй открытой рамке .
Разработаны web-серверы, позволяющие предсказывать наличие IRES в вирусных РНК (VIPS) или в вирусных и клеточных РНК (IRESPred) .
В 2016 году был предложен алгоритм RNAiFold2T для разработки особых РНК-термометров , содержащих IRES. Кэпнезависимая трансляция таких термо-IRES-элементов примерно на 50 % интенсивнее при 42 °С, чем при 30 °С. Впрочем, эффективность их трансляции всё равно меньше, чем у IRES дикого типа, которая не зависит от температуры .
Примечания
- ↑ , p. 14.
- ↑ Balvay L. , Soto Rifo R. , Ricci E. P. , Decimo D. , Ohlmann T. (англ.) // Biochimica et biophysica acta. — 2009. — Vol. 1789, no. 9-10 . — P. 542—557. — doi : . — .
- Colussi T. M. , Costantino D. A. , Zhu J. , Donohue J. P. , Korostelev A. A. , Jaafar Z. A. , Plank T. D. , Noller H. F. , Kieft J. S. (англ.) // Nature. — 2015. — Vol. 519, no. 7541 . — P. 110—113. — doi : . — .
- Pelletier J. , Sonenberg N. (англ.) // Nature. — 1988. — Vol. 334, no. 6180 . — P. 320—325. — doi : . — .
- Jang S. K. , Kräusslich H. G. , Nicklin M. J. , Duke G. M. , Palmenberg A. C. , Wimmer E. (англ.) // Journal of virology. — 1988. — Vol. 62, no. 8 . — P. 2636—2643. — .
- ↑ Kozak M. (англ.) // Nucleic acids research. — 2005. — Vol. 33, no. 20 . — P. 6593—6602. — doi : . — .
- ↑ , p. 15.
- Fitzgerald K. D. , Semler B. L. (англ.) // Biochimica et biophysica acta. — 2009. — Vol. 1789, no. 9-10 . — P. 518—528. — doi : . — .
- , p. 14—15.
- Cobbold L. C. , Spriggs K. A. , Haines S. J. , Dobbyn H. C. , Hayes C. , de Moor C. H. , Lilley K. S. , Bushell M. , Willis A. E. (англ.) // Molecular and cellular biology. — 2008. — Vol. 28, no. 1 . — P. 40—49. — doi : . — .
- Thompson S. R. (англ.) // Trends in microbiology. — 2012. — Vol. 20, no. 11 . — P. 558—566. — doi : . — .
- Kieft J. S. (англ.) // Trends in biochemical sciences. — 2008. — Vol. 33, no. 6 . — P. 274—283. — doi : . — .
- Brown, T.A. (неопр.) . — New York, New York: Garland Science Publishing, 2007. — С. . — ISBN 0 8153 4138 5 .
- Hellen C. U. , Sarnow P. (англ.) // Genes & development. — 2001. — Vol. 15, no. 13 . — P. 1593—1612. — doi : . — .
- Asnani M. , Pestova T. V. , Hellen C. U. (англ.) // Nucleic acids research. — 2016. — Vol. 44, no. 7 . — P. 3390—3407. — doi : . — .
- Filbin M. E. , Kieft J. S. (англ.) // Current opinion in structural biology. — 2009. — Vol. 19, no. 3 . — P. 267—276. — doi : . — .
- Niepmann M. (англ.) // Biochimica et biophysica acta. — 2009. — Vol. 1789, no. 9-10 . — P. 529—541. — doi : . — .
- Khong A. , Bonderoff J. M. , Spriggs R. V. , Tammpere E. , Kerr C. H. , Jackson T. J. , Willis A. E. , Jan E. (англ.) // Viruses. — 2016. — Vol. 8, no. 1 . — doi : . — .
- Chatterjee S. , Pal J. K. (англ.) // Biology of the cell / under the auspices of the European Cell Biology Organization. — 2009. — Vol. 101, no. 5 . — P. 251—262. — doi : . — .
- Dibrov S. M. , Parsons J. , Carnevali M. , Zhou S. , Rynearson K. D. , Ding K. , Garcia Sega E. , Brunn N. D. , Boerneke M. A. , Castaldi M. P. , Hermann T. (англ.) // Journal of medicinal chemistry. — 2014. — Vol. 57, no. 5 . — P. 1694—1707. — doi : . — .
- Komar A. A. , Hatzoglou M. (англ.) // Frontiers in oncology. — 2015. — Vol. 5. — P. 233. — doi : . — .
- Jiang X. , Kanda T. , Nakamoto S. , Saito K. , Nakamura M. , Wu S. , Haga Y. , Sasaki R. , Sakamoto N. , Shirasawa H. , Okamoto H. , Yokosuka O. (англ.) // Biochemical and biophysical research communications. — 2015. — Vol. 458, no. 4 . — P. 908—912. — doi : . — .
- Qian S. , Fan W. , Qian P. , Zhang D. , Wei Y. , Chen H. , Li X. (англ.) // Viruses. — 2015. — Vol. 7, no. 4 . — P. 1613—1626. — doi : . — .
- Hou H. Y. , Lu W. W. , Wu K. Y. , Lin C. W. , Kung S. H. (англ.) // The Journal of general virology. — 2016. — Vol. 97, no. 5 . — P. 1122—1133. — doi : . — .
- Buijs P. R. , Verhagen J. H. , van Eijck C. H. , van den Hoogen B. G. (англ.) // Human vaccines & immunotherapeutics. — 2015. — P. 0. — doi : . — .
- Ngoi S. M. , Chien A. C. , Lee C. G. (англ.) // Current gene therapy. — 2004. — Vol. 4, no. 1 . — P. 15—31. — .
- Kolekar P. , Pataskar A. , Kulkarni-Kale U. , Pal J. , Kulkarni A. (англ.) // Scientific reports. — 2016. — Vol. 6. — P. 27436. — doi : . — .
- Garcia-Martin J. A. , Dotu I. , Fernandez-Chamorro J. , Lozano G. , Ramajo J. , Martinez-Salas E. , Clote P. (англ.) // Bioinformatics. — 2016. — Vol. 32, no. 12 . — P. 360—368. — doi : . — .
Литература
- Lucy W. Barrett, Sue Fletcher, Steve D. Wilton. . — SpringerBriefs in Biochemistry and Molecular Biology, 2013. — 57 p. — ISBN 978-3-0348-0679-4 .
Ссылки
- .
- 2020-08-29
- 1