Хорошие статьи
- 1 year ago
- 0
- 0

Программи́руемая кле́точная ги́бель , или программи́руемая кле́точная смерть , или запрограмми́рованная ги́бель кле́ток ( ПКГ , англ. Programmed cell death ) — гибель клетки , которая происходит за счёт запрограммированных внутриклеточных процессов. Ко второму десятилетию XXI века насчитывается более десяти известных видов программируемой клеточной гибели. С 2005 года классификацией видов клеточной гибели занимается Комитет по номенклатуре видов клеточной гибели ( англ. Nomenclature Commitee on Cell Death ). Программируемая клеточная гибель описана для всех крупных групп эукариот : животных , растений , грибов , слизевиков и даже одноклеточных организмов (например, дрожжей ). ПКГ выполняет множество функций как на уровне клетки, так и на уровне целого организма: у животных она играет важнейшую роль в развитии , с её помощью элиминируются повреждённые клетки, у растений она задействована в образовании тканей , состоящих из мёртвых клеток, таких как ксилема . Программируемая клеточная гибель известна не только у эукариот: несколько видов программируемой гибели было описано у бактерий . Все виды программируемой клеточной гибели можно подразделить на внешние, которые запускаются сигналами извне клетки, и внутренние, вызванные нарушениями в функционировании клеток .
С точки зрения морфологии долгое время выделялось три основных вида программируемой клеточной смерти:
Позже была принята более сложная классификация видов программируемой клеточной гибели, которая построена не на морфологических деталях, а на генетических , биохимических , фармакологических и функциональных особенностях. Однако выделенные таким образом виды гибели далее делят на две группы по морфологии: в одну относят виды смерти, которые морфологически близки к апоптозу, а в другую — те, которые морфологически близки к некрозу. Таким образом, для каждого вида программируемой клеточной гибели присущ свой набор свойств, от полностью апоптотического до полностью некротического .
По состоянию на 2018 год выделяют следующие виды программируемой клеточной гибели :
Зависимая от лизосом клеточная гибель начинается с нарушений клеточного гомеостаза и пермеабилизации (изменения проницаемости) мембран лизосом. Она наблюдается при многих патофизиологических процессах: воспалении, перестройке тканей (например, перестройке ткани молочных желёз после лактации ), старении , нейродегенеративных заболеваниях , сердечно-сосудистых заболеваниях и ответе на внутриклеточные патогены .
После пермеабилизации мембран лизосом содержимое последних выходит в цитозоль , где в том числе оказываются в результате протеолитические ферменты семейства катепсинов , которые разрушают содержимое клетки. Процессы, предшествующие пермеабилизации лизосомальных мембран и запускающие её, не вполне ясны. В некоторых условиях она происходит после пермеабизизации внешней митохондриальной мембраны в ходе внутреннего апоптоза. В других случаях пермеабилизация мембран лизосом происходит раньше мембран митохондрий при участии белка . Важную роль в запуске повышения проницаемости лизосомальных мембран играют активные формы кислорода .

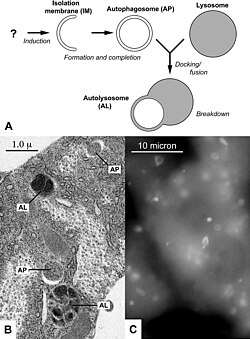
Зависимая от аутофагии клеточная гибель подразумевает активацию молекулярных механизмов аутофагии (всех или части), которые приводят к образованию аутофагосом — везикул с двойной мембраной . Аутофагия является важным процессом, составляющим часть клеточного ответа на стресс, поэтому её нарушения приводят к разнообразным дефектам развития и заболеваниям. У дрозофилы аутофагия задействована в обновлении выстилки средней кишки и деградации личиночных слюнных желёз . Зависимая от аутофагии клеточная гибель вносит свой вклад в патогенез ряда заболеваний и у человека . Например, при некоторых патологических состояниях по пути аутофагии погибают нейроны . Разновидность зависимой от аутофагии клеточной гибели, в которой задействована Na + /K + -АТФаза , известна как аутоз .
Иммунногенной клеточной гибелью называют те виды клеточной смерти, которые сопровождаются активацией адаптивного иммунного ответа , направленного против эндогенных (клеточных) или экзогенных ( вирусных ) антигенов , которые экспрессирует погибающая клетка. Иммунногенную клеточную гибель вызывают сравнительно немногие факторы: вирусные инфекции , некоторые противораковые препараты (например, антрациклины и ), некоторые виды радиотерапии , а также фотодинамическая терапия , основанная на гиперицине . Как правило, иммунный ответ запускают следующие ассоциированные с повреждениями молекулярные паттерны ( англ. Damage-associated molecular pattern, DAMP ), экспрессируемые умирающей клеткой: , АТФ , белок HMGB1 , , нуклеиновые кислоты , происходящие от раковых клеток , и аннексин A1 .

Внутренний апоптоз запускают разнообразные изменения окружающей среды клетки: отсутствие факторов роста , повреждения ДНК , стресс эндоплазматического ретикулума (ЭПР), активные формы кислорода , нарушения репликации ДНК , дефекты митоза и нарушения функционирования микротрубочек . Клетки, погибающие путём апоптоза, сохраняют целостность плазматической мембраны и некоторую метаболическую активность. Они распадаются на везикулы — апоптотические тельца, которые фагоцитируются другими клетками. Критический этап внутреннего апоптоза — необратимая пермеабилизация внешних митохондриальных мембран, которая контролируется различными белками семейства . В результате в цитозоль выходят проапоптотические факторы, которые в обычное время находятся в . Важнейшим из них является белок дыхательной цепи цитохром c . В цитозоле он связывается с белком APAF1 и про-каспазой 9, формируя комплекс, известный как апоптосома . В ней каспаза 9 активируется, формируя димеры , которые сами себя разрезают и тем самым активируют, и начинает активировать другие каспазы , внося в них разрезы. Каспазы — это протеазы , которые разрушают все белки клетки и вызывают смерть клетки .
Внешний апоптоз запускается изменениями в клетки. Ключевую роль в запуске внешнего апоптоза играют два типа рецепторов клеточной мембраны: рецепторы смерти , которые активируются при связывании с лигандом , а также рецепторы, которые активируются, когда концентрация их лиганда опускается ниже некоторого значения. К числу рецепторов смерти относятся, например, и ряд других рецепторов суперсемейства факторов некроза опухолей ( англ. tumor necrosis factor, TNF ). Когда рецептор смерти активируется, у его внутриклеточной части собирается особый многобелковый комплекс — DISC (от англ. death-inducing silencing complex ). Он регулирует активацию и функционирование каспазы 8 (или, в некоторых случаях, каспазы 10). Вслед за ней активируются и остальные каспазы, которые разрушают клеточные белки и приводят к её гибели .
MPT-зависимый некроз начинается при особых изменениях внутриклеточных условий, таких как сильный окислительный стресс и значительное повышение концентрации ионов кальция в цитозоле. Сокращение MPT происходит от англ. mitochondrial permeability transition — нарушение проницаемости митохондрий, так как при этом виде клеточной гибели внутренняя митохондриальная мембрана становится проницаемой для малых молекул , что приводит к исчезновению электрохимического градиента на ней, осмотическому разрушению обеих митохондриальных мембран и в конечном счёте гибели клетки в виде некроза .

Некроптоз вызывается различными изменениями во внутренней и внешней среде клетки, которые детектируются особыми рецепторами смерти (например, Fas), рецепторами распознавания патогенов (например, Toll-подобными рецепторами 3 и 4 ), а также белком , связывающимся с Z-ДНК . Морфологически смерть клетки происходит в форме некроза. Критически важную роль в запуске некроптоза играют протеинкиназы и , которые активируются рецепторами. Некроптоз не только связан с ответом организма на стресс, он обеспечивает гибель дефектных организмов до рождения и участвует в регуляции гомеостаза T-лимфоцитов во взрослом организме .
Параптоз представляет собой тип неапоптотической гибели клеток, который опосредован МАРК через активацию . Характеризуется внутриклеточным образованием вакуолей и набуханием митохондрий.
Ферроптозу, как правило, предшествует серьёзное повреждение клеточных липидов в результате образования активных форм кислорода и появления свободных ионов железа в клетке. Окисление липидов происходит именно из-за ионов железа, за что данный вид клеточной гибели и получил своё название . Морфологически ферроптоз — это некроз, при котором происходят серьёзные повреждения митохондрий: они сжимаются, в них исчезают кристы и разрушается внешняя мембрана. В ферроптозе не задействованы каспазы и белки, осуществляющие аутофагию. При этом виде клеточной гибели происходит окисление некоторых полиненасыщенных жирных кислот , например, арахидоновой кислоты , и образуются гидропероксиды липидов. Иногда окисление липидов может происходить под действием ферментов липоксигеназ и циклооксигеназ . Им противодействует глутатионпероксидаза 4 (GSH4). Ферроптозу также препятствуют ферростатин-1, липрокстатин-1, а также витамин E , кофермент Q 10 и подобные соединения с антиоксидантной активностью, которые отвлекают на себя активные формы кислорода и не дают им взаимодействовать с липидами .
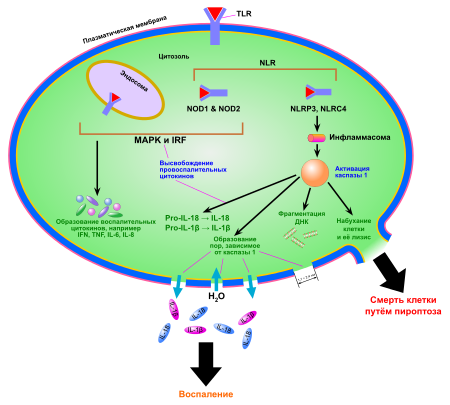
Пироптоз активируется в ходе реакций врождённого иммунитета . При пироптозе происходит особая конденсация хроматина, отличающаяся от конденсации хроматина при апоптозе. Клетка разбухает, происходит пермеабилизация мембраны. В пироптозе ведущую роль играет провоспалительная каспаза 1 , однако в некоторых случаях вместо неё выступают другие каспазы, например, каспаза 3 . Пироптоз задействован в развитии многих патологических состояний, например, смертельного септического шока , вызванного бактериальными липополисахаридами . Именно бактериальные липополисахариды, попадающие в цитоплазму клеток организма-хозяина, вероятно, играют ведущую роль в запуске пироптоза .
Партанатоз — это форма программируемой клеточной гибели, характеризующаяся гиперактивацией поли(АДФ-рибоза)полимеразы 1 ( ) — белка, участвующего в клеточном ответе на повреждение ДНК. Однако партанатоз может происходить не только при сильном повреждении ДНК путём алкилирования , но также при окислительном стрессе, гипоксии , гипогликемии или воспалении. Главную роль в гиперактивации PARP1, особенно в нейронах, играют активные формы азота , в том числе оксид азота NO . Гиперактивация PARP1 имеет цитотоксические эффекты, такие как истощение пула NAD + и АТФ (что приводит к нарушению биоэнергетического и окислительно-восстановительного равновесия клетки), а также накопление полимеров поли(АДФ-рибозы) и поли(АДФ-рибоз)илированных белков в митохондриях (из-за чего теряется мембранный потенциал и пермеабилизуется внешняя митохондриальная мембрана) .

Энтоз — это форма клеточного каннибализма, которая происходит в здоровых тканях и опухолях млекопитающих . Живая клетка поглощается другой клеткой, не обладающей способностью к фагоцитозу. Часто, но не всегда, поглощённая клетка погибает. Как правило, энтоз запускается, когда эпителиальная клетка теряет контакт с внеклеточным матриксом , хотя для этого могут быть и другие причины: неотрегулированная экспрессия миозинов при формировании межклеточных контактов , различия в механических свойствах соседних клеток и метаболический стресс. У раковых клеток энтоз может запускаться при митозе . Гибель поглощённой клетки не зависит от каспаз и белков BCL2, играющих важнейшую роль в апоптозе. По крайней мере в некоторых случаях гибель происходит в виде особой формы аутофагии .

Первоначально эта форма гибели была описана у нейтрофилов , которые, умирая, выбрасывают наружу сеть из волокон, содержащих хроматин и гистоны , связанные с цитозольными белками. Эти волокна были названы англ. neutrophil extracellulae traps (NET), и форма смерти получила название нетоз ( англ. NETosis ). Выброс NET может быть вызван микробами , активацией особых рецепторов (например, Toll-подобных). Существенная доля ДНК, входящей в состав этих волокон, имеет митохондриальное , а не ядерное происхождение. NET могут выбрасывать и другие клетки, отличные от нейтрофилов: тучные клетки , эозинофилы и базофилы , причём выброс NET не всегда приводит к гибели клетки. NET обладают не только антимикробным эффектом; показана их роль в таких заболеваниях, как диабет и рак .

Программируемая клеточная гибель зафиксирована у беспозвоночных животных, в частности, у губок . У губок экспрессируются такие белки ПКГ, как каспазы, белки с и Bcl-2 . Более того, Bcl-2 губок подавляет апоптоз и в клетках позвоночных . Первоначально молекулярные механизмы ПКГ были описаны у нематоды Caenorhabditis elegans . Поскольку количество клеток в теле взрослого червя жёстко фиксировано и одинаково для всех особей, количество актов ПКГ также фиксировано: в ходе развития червя клетки убивают сами себя ровно 131 раз. Ключевую роль в ПКГ у C. elegans играют белки Ced-4 и Ced-3 с каспазной активностью. В обычных условиях Ced-4 подавлен белком Ced-9, который локализован во внешней митохондриальной мембране. Когда клетка получает извне сигнал к началу ПКГ, Ced-9 инактивируется, а Ced-4 активируется и в свою очередь активирует Ced-3, который запускает работу протеаз и нуклеаз . У членистоногих ПКГ впервые происходит при образовании нервной системы , когда происходят дифференцировка и деление клеток эктодермы , причём одна из дочерних клеток становится нейробластом , а другая погибает . Более того, вследствие ПКГ у самцов и самок некоторые органы иннервируются по-разному . У плодовой мушки Drosophila melanogaster имеется несколько каспаз и ингибиторов апоптоза, кроме того, некоторые белки ПКГ, такие как REAPER, HID и GRIM, могут быть специфичны для насекомых .
У растений программируемая клеточная гибель наблюдается при образовании ксилемы и семян , старении , предотвращении самоопыления , а также под действием стрессов (солевого, температурного , окислительного ) и патогенов . Как и у животных, у растений существует несколько видов ПКГ, однако чаще всего она сходна с апоптозом и сопровождается фрагментацией ДНК, выходом цитохрома c из митохондрий, сжатием клетки, образованием активных форм кислорода и выходом фосфатидилсерина на внешний слой мембраны. В то же время, хотя у растений отсутствуют каспазы, известно, что ингибиторы каспаз животных могут подавлять программируемую клеточную смерть и у растений. Главная роль в программируемой клеточной гибели у растений принадлежит — серин -зависимым аспартат -специфичным протеазам. В здоровых тканях фитаспазы находятся в апопласте , а при индукции ПКГ входят в цитозоль .
У грибов программируемая клеточная гибель наблюдается при образовании спор полового и бесполого размножения , формировании плодового тела или склероция , в реакции вегетативной несовместимости, при патогенезе , стрессовых условиях и на заключительных этапах старения. Этим назначение ПКГ у грибов отличается от такового у животных, у которых она прежде всего важна для развития. В общем случае ПКГ грибов аналогична внутреннему апоптозу животных. ПКГ детально изучена у дрожжей Saccharomyces cerevisiae и может запускаться разнообразными внутренними факторами, причём внешний механизм активации ПКГ не обнаружен. У них нет и очевидных гомологов ключевых белков апоптоза животных, таких как Bcl-2, p53 , , PARP и даже каспазы. В то же время гомологи некоторых регуляторных апоптотических белков отсутствуют у дрожжей, но есть у мицелиальных грибов. У ПКГ проявляется при старении мицелия, которое обусловлено действием активных форм кислорода. В ходе ПКГ у P. anserina функционируют цистеиновые протеазы с каспазной активностью .
Плодовое тело слизевика Dictyostelium discoideum имеет ножку, образованную мёртвыми клетками. Эти клетки подверглись ПГК, похожей на аутофагию животных по степени развития вакуолей и конденсации хроматина , кроме того, в отличие апоптоза, фрагментации ДНК не происходит . Предки слизевиков отделились от остальных эукариот более миллиарда лет назад до отделения предков растений и грибов, что свидетельствует о древнем происхождении программируемой клеточной гибели .
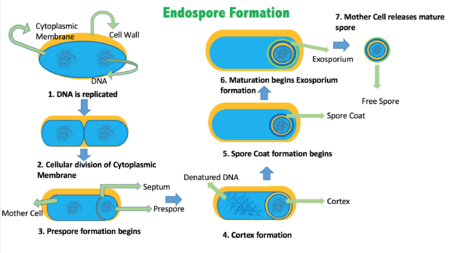
У бактерий известно несколько форм программируемой клеточной гибели. В условиях стресса (окислительного стресса, воздействия радиации , нехватки питательных веществ , фаговой инфекции ) часть клеток погибает на благо колонии . Чаще всего смерть происходит при участии систем токсин-антитоксин различных типов. Бактериофаги , геном которых представлен двуцепочечной ДНК, вызывают гибель заражённых клеток в конце литического цикла для высвобождения новых вирионов с помощью холин-эндолизиновой системы. Маленькие белки встраиваются в мембрану, давая возможность выйти наружу . Эндолизины гидролизуют пептидогликан , разрушают клеточную стенку и вызывают лизис клетки. Гибель бактериальных клеток наблюдается на разных этапах развитии колонии и при отсутствии стресса: при споруляции , генетической трансформации , образовании плодовых тел и формировании биоплёнок . Механизмы программируемой клеточной гибели во всех перечисленных случаях различны .
Физиологическое значение программируемой клеточной гибели огромно. У животных она играет важнейшую роль в развитии многих органов и тканей, а также старении. В ходе развития нервной системы множество клеток-предшественников нейронов погибают, так что количество нейронов в мозге взрослого животного существенно меньше, чем их было заложено в ходе эмбрионального развития . Апоптоз задействован в морфогенезе животных (в частности, апоптозом погибают клетки между пальцами, за счёт апоптоза исчезает хвост у головастика ). Иммуногенная клеточная смерть и пироптоз наряду с апоптозом задействованы в работе защитных систем организма. Подавление ПКГ очень часто связано со злокачественным перерождением клетки . У растений ПКГ участвует в образовании тканей, состоящих из мёртвых клеток, например, ксилемы. Кроме того, на ПКГ основана самонесовместимость при опылении : если на рыльце попадает пыльца от того же растения, то особые белки на рыльце запускают ПКГ клеток пыльцевого зерна . У грибов ПКГ обеспечивает вегетативную несовместимость, то есть не даёт сливаться гифам одного штамма , а также задействована в созревании спор полового и бесполого размножения .
.jpg)
Сама концепция программируемой клеточной гибели была предложена ( англ. Lockshin ) и Уильямсом в 1964 году по отношению к развитию некоторых тканей у насекомых . Примерно через 8 лет появился термин «апоптоз». Первые сведения о механизмах ПКГ появились при изучении белка Bcl-2 — продукта онкогена , экспрессия которого активируется при хромосомных транслокациях , которые часто наблюдаются при фолликулярной лимфоме . В отличие от других известных к этому моменту продуктов онкогенов, Bcl-2 вызывает злокачественное перерождение не за счёт непрерывной стимуляции деления, а за счёт предотвращения программируемой клеточной гибели . По сей день программируемая клеточная гибель интенсивно исследуется. В 2002 году Нобелевская премия по физиологии и медицине была присуждена за открытия в молекулярной биологии программируемой клеточной гибели Сиднею Бреннеру , Роберту Хорвицу и Джону Салстону , а в 2016 году этой награды был удостоен Ёсинори Осуми , исследовавший один из видов программируемой клеточной гибели — аутофагию .