

Сумчатые жабы
- 1 year ago
- 0
- 0

Жабы , или настоящие жабы ( лат. Bufonidae ) , — семейство бесхвостых земноводных , единственное, в котором все представители называются «жабами», хотя некоторые виды можно назвать лягушками из-за похожих внешних черт (например, ателопов ). На более высоком таксономическом уровне применение термина «жаба» не ограничивается только данным семейством, а также используется для видов из других семейств (например: жаба-повитуха , седлоносная жаба , лягушковидная жаба , носатая жаба ).
Первые представители семейства эволюционно возникли около 80 млн лет назад в Южной Америке и быстро распространились по всему миру. Самые крупные жабы ( ага ) могут достигать длины 25—50 см и весить более трёх килограммов; наименьшие представители семейства имеют длину не более 2 см. Рёбра отсутствуют. Кожа, как правило, сухая на ощупь, толстая, зачастую покрыта многочисленными гладкими или острыми бугорками-бородавками разного размера; позади глаз хорошо заметны околоушные железы ( паротиды ), которые у ряда тропических форм отсутствуют. На них содержится яд буфотоксин. Жабы — ночные наземные амфибии, входящие в воду лишь для откладывания яиц, однако существуют и полуводные виды (например, анзонии ) и древесные ( древесные жабы ). Зимуют в земле. Питаются исключительно мелкими беспозвоночными : насекомыми , червями и улитками . Способы размножения разные — от откладывания икры в воду и последующего выхода из неё свободноплавающих личинок, до яйцеживорождения и прямого наземного развития. Большинству видов свойственно внешнее оплодотворение , однако некоторым — внутреннее. На ноябрь 2018 года в семейство включают 52 рода и 609 видов.
Слово жаба происходит от праславянского * gēbā (ср. бел. жа́ба, укр. жа́ба , болг. жа́ба , сербохорв. жȁба , словен. žába , чеш. žába , слвц. žaba , польск. żaba , в.-луж., н.-луж. žaba , полаб. zobó ). На индоевропейском уровне родственно др.-прусск. gabawo «жаба», др.-сакс. quappa , ср.-нидерл. quappe «налим», ср.-нж.-н. quabbe , quobbe «влажная масса», возм., также лат. būfō (ср. научное название жаб) .
В разговорной речи животных из отряда бесхвостых земноводных называют « лягушками » или «жабами» (возможно провести параллель с латинскими терминами, используя в качестве основы лат. rana для слова «лягушка» и лат. bufo для слова «жаба»). Визуальное отличие одних от других состоит в том, что кожа жаб бородавчатая . При попытке наложить разговорные термины «лягушка» и «жаба» на принятую классификацию отряда бесхвостых оказывается, что практически все семейства бесхвостых включают и тех и других .
Считается, что первые представители семейства Bufonidae произошли из Южной Америки. Некоторые исследования датируют происхождение группы после распада Гондваны, около 78-98 миллионов лет назад в позднем меловом периоде . Напротив, другие исследования датируют происхождение группы ранним палеоценом . Первые настоящие жабы, вероятно, распространились из Южной Америки в течение эоцена, при этом всё расселение произошло в период от эоцена до олигоцена, что свидетельствует о чрезвычайно быстром расхождении, вероятно, которому способствовали изменяющиеся климатические условия палеогена .
Следующая филогения большинства родов семейства основана на исследованиях Portik and Papenfuss, 2015 , Chan et al . , 2016 , Chandramouli et al. , 2016 , и Kok et al. , 2017 :
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


Внешность и размеры жаб сильно варьируют. Длина тела колеблется от 20 мм ( гвианский арлекин ) до 550 мм ( жаба-ага ) . Масса — от нескольких граммов до 3,65 кг. Туловище обычно широкое, грузное, конечности короткие . Рёбра отсутствуют. Кожа, как правило, сухая на ощупь, толстая, зачастую покрыта многочисленными гладкими или острыми бугорками-бородавками разного размера . Язык узкий, большой. Зубы в большинстве случаев частично или полностью редуцированы . Зрачок горизонтальный, пальцы задних конечностей частично связаны плавательной перепонкой . Поперечные отростки крестцового позвонка не расширены в диски . Настоящие жабы широко распространены и обитают на всех континентах, кроме Австралии и Антарктиды, обитая в самых разных средах, от засушливых районов до тропических лесов. Большинство откладывает яйца парными цепочками, из которых вылупляются головастики, хотя у представителей рода Nectophrynoides прямо из яиц вылупляются миниатюрные жабы .
Позади глаз хорошо заметны околоушные железы ( паротиды ), которые у ряда тропических форм отсутствуют. Эти железы содержат алкалоидный яд — буфотоксин , который жабы выделяют при стрессе. Состав и пропорции яда у разных видов различаются . Яд некоторых жаб (например, колорадской жабы ) используется в качестве рекреационного психотропного средства .
У самцов семейства присутствует уникальный неотенический орган Биддера — рудиментарный яичник , который развивается на переднем крае личиночных семенников и может сохраняться у многих взрослых амфибий . При определённых условиях орган становится активным яичником, и самец, по сути, становится самкой .
Жабы — ночные наземные амфибии, входящие в воду лишь для откладывания яиц, однако существуют и полуводные виды (например, анзонии ) и древесные ( древесные жабы ). Зимуют в земле. Все виды жаб хищные; питаются исключительно мелкими беспозвоночными : насекомыми , червями и улитками . Существуют крупные виды, которые поедают других земноводных, небольших млекопитающих и птиц . Одни ловят свою быстро двигающуюся добычу клейким языком, другие запихивают пищу в рот передними конечностями . Полезны истреблением сельскохозяйственных вредителей :
Польза людям от жабьих ночных похождений огромна. В США попытались приблизительно оценить стоимость услуг, веками, ночь за ночью, приносимых жабами лесному и фермерскому делу: миллиарды долларов в год! Акимушкин И.И., Мир животных. Птицы, рыбы, земноводные и пресмыкающиеся. С. 391
В сумерках жабы вылезают из подвалов, нор, пещер, из-под половиц сараев и других тёмных, укромных, скрытных мест. Скачками или перебежками («жабьей рысью»), плотно прижавшись к земле, выходят эти земноводные за вредителями огородов, садов, полей и т. п.: поедают насекомых, слизней, червей . Если жабу перенести из водоёма, в котором она родилась и выросла, в другой, она отправится искать свой пруд и обычно находит, если амфибию занесли не слишком далеко. Жабы возвращаются и упорно ищут воду даже в тех водоёмах, которые пересохли или засыпаны, но в которых прежде они жили головастиками .

Большинству видов свойственно внешнее оплодотворение , однако некоторым — внутреннее ( Nectophrynoides , ). Внутреннее оплодотворение встречается у четырёх (по другим исследованиям, несколько больше) родов семейства настоящих жаб :
Ascaphus (все виды) и Eleutherodactylus (два вида, E. coqui и E. jasperi ) являются единственными другими родами лягушек, у которых есть внутреннее оплодотворение . Limnonectes larvaepartus также размножаются внутренним оплодотворением .
Способы размножения разные — от откладывания икры в воду и последующего выхода из неё свободноплавающих личинок, до яйцеживорождения и прямого наземного развития . Большинство представителей семейства нерестятся в стоячих или проточных водоёмах. Филиппинские жабы рода Pelophryne откладывают икру в пазухи листьев на высоте нескольких метров. У головастиков некоторых видов, живущих в быстротекущих ручьях ( Ansonia , Atelopus ) развиваются брюшные присоски, с помощью которых они прикрепляются к подводным предметам .
У жаб, как и у всех бесхвостых наблюдаются два типа брачного поведения: сезонный и спонтанный. При первом типе, характерном для большинства видов, в определённые периоды года взрослые особи собираются для размножения в прудах, озёрах или ручьях. Многие жабы возвращаются к тому водоёму, в котором они развивались головастиками. В результате можно наблюдать сезонные миграции , в которые вовлечены тысячи особей. При спонтанном типе размножения взрослые жабы прибывают на места размножения, подталкиваемые некой внешней причиной (например, выпадением осадков в засушливой местности). В данном случае размножение и развитие икринок и головастиков происходит с максимальной скоростью, до того, как недолговечные водоёмы (лужи) успеют высохнуть .

Самцы видов жаб с сезонным типом брачного поведения обычно первыми прибывают в места размножения и остаются там продолжительное время, призывая кваканьем самок и защищая свою территорию от других самцов. Самки появляются позже лишь на время спаривания и икрометания. В этой ситуации количество самцов у кромки воды всегда превышает количество самок, поэтому среди самцов существует жёсткая конкуренция. Они взбираются на бугорок или пук травы, высунувшись наполовину из воды; крупные и сильные самцы издают более низкие звуки и захватывают самые привлекательные для размножения места. Самцы повторяют этот звук около 35-40 раз в минуту. Наблюдения показали, что самки предпочитают именно таких самцов . Поскольку кваканье требует больших затрат энергии, иногда роли меняются и зовущий самец оставляет свою территорию и становится перехватчиком .
Многие жабы зимуют под землёй; в конце марта-апреле амфибии пробираются к воде для икрометания и спаривания. Во время дождя десятки тысяч жаб пытаются спариться. Самцы появляются первыми: они хорошо помнят свои прошлогодние пути (длиной до километра), которые ранее приводили их к водоёмам. Серые и прочие жабы запоминают также самые приметные ориентиры и могут видеть даже ультрафиолетовые лучи .
У видов со спонтанным типом брачного поведения основной упор делается не на выбор партнёра, а на скорость размножения. Соответственно меняется и тактика. Самец, первым обнаруживший подходящее место (лужу), начинает громко призывать сородичей, на его зов сбегаются все окрестные представители обоих полов, которые тут же подхватывают призыв, создавая хор, слышный издалека. Иногда подходящие условия отсутствуют годами, и, соответственно, эти виды не размножаются несколько лет подряд .
Как правило, процесс спаривания заключается в том, что самец осёдлывает самку в воде и обхватывает её туловище. Самка мечет икру, а самец покрывает её своей спермой (внешнее оплодотворение). У некоторых видов самцы могут задерживать икринки задними лапами на время, необходимое для оплодотворения . В этом плане уникальными среди бесхвостых являются виды западноафриканского рода Nimbaphrynoides , которые размножаются живорождением , и виды танзанийского рода Nectophrynoides , практикующие яйцеживорождение . В обоих случаях имеет место внутреннее оплодотворение, а самка рожает полностью развитых молодых лягушек (сеголеток), которые минуют стадию головастика .

В Африке и в Южной Азии живут живородящие жабы Nectophrynoides длиной около шести сантиметров. Обитают в горных лесах, на земле и в траве, на листьях деревьев. Учёные узнали об этом в 1905 году, когда была открыта . У самок расширенные концы яйцеводов образовали некое подобие матки, где развиваются оплодотворенные яйца, затем зародыши. Рождаются они вполне сформированными лягушатами — до 135 молодых амфибий. Второй вид этого рода, древесная жаба Торнира , описан в 1906 году зоологом Жаном Роуксом . Рост до трех сантиметров. Новорожденных детенышей — до 35 .
Третий вид, , был обнаружен лишь в 1942 году за пять тысяч километров к западу от места обитания двух первых, в Гвинее. Жабы размером до 2,5 см. Ареал достаточно невелик: несколько квадратных километров травяных лугов на склонах горы Нимба. В сезон дождей, от мая до августа, этих жаб-самцов до пятисот на ста квадратных метрах луга. Затем дожди кончаются, и амфибии скрываются в разных трещинах земли почти девять месяцев, дожидаясь, пока вновь польют дожди. Перед этим самцы успевают оплодотворить самок, и те вынашивают эмбрионов. Эмбрионы кормятся, как у акул или альпийских саламандр: питательными веществами, запасенными в яйцах или зародышах, развивающихся с запозданием; каинизма у этих амфибий нет — как у червяг и скатов-хвостоколов , матка матери производит творожистую массу. Зародыши быстро растут, проглатывая её: перед рождением они, как правило, лишь втрое меньше вынашивающей их матери. Детёнышей от двух до двадцати особей, однако чем больше зародышей развивается, тем меньше ростом рождающиеся жабята .
Жабы имеют короткие задние конечности и передвигаются шагом, а не прыжками . Быстрое передвижение у представителей этого семейства обеспечивают либо ускоренные движения конечностей (быстрая ходьба), либо быстрые короткие прыжки. В одном из экспериментов жаба была посажена на беговую дорожку, которая вращалась с разной скоростью. При замере количества кислорода, используемого жабой, было обнаружено, что для этого вида передвижение маленькими прыжками на длинные дистанции является энергетически неэффективным, но может быть оправдано для быстрого перемещения на короткие расстояния .

Жабы, обитающие в воде, адаптированы к плаванию; для строения их тела характерны сильные задние конечности и наличие плавательных межпальцевых перепонок. Перепонки увеличивают площадь поверхности ступни (наподобие ласт) и способствуют быстрому передвижению лягушек в воде. Головастики, как правило, имеют большие хвостовые плавники, придающие ускорение, направленное вперёд, когда хвост движется из стороны в сторону . В воде бесхвостые наиболее беспомощны во время метаморфоза, когда хвост уже атрофирован, а ноги ещё не полностью функциональны .
Кваканье — это звук, который производит проходящий через гортань воздух . Кваканье некоторых жаб слышно дальше чем за километр . Виды бесхвостых, обитающие преимущественно вдоль проточной воды, часто не имеют адаптаций для усиления звука, так как шумный фон делает звуковую коммуникацию неэффективной . Вместо неё эти виды используют визуальные способы общения («семафорят» друг другу). Как показывают наблюдения, основная доля коммуникации приходится на привлечение самок. Самцы могут квакать в одиночестве, а могут создавать хор , когда много самцов собирается в удобных для спаривания местах. Самки предпочитают самцов, издающих более громкие и низкие звуки, которые указывают на здорового и крепкого самца, способного производить наилучшее потомство .


В яйцах развиваются личинки жаб. Через несколько недель крохотные головастики вылезают из икринок. Обычно из всех разом. Их тело, как правило, имеет овальную форму, хвост длинный, сплющенный по вертикали, приспособлен для плаванья. У головастиков хрящевой скелет, глаза лишены век, есть боковая линия , для дыхания служат жабры . В начале у головастиков появляются внешние жабры, а позже внутренние (жаберный мешок закрывает жабры и передние ноги). Развивающиеся лёгкие служат дополнительным дыхательным органом. У головастиков нет настоящих зубов, но у многих видов челюсти покрыты параллельными рядами кератиновых зубчиков (два ряда на верхней челюсти, три ряда на нижней и роговой клюв). Количество рядов и точная морфология рта варьируют у разных видов и могут служить диагностическим признаком . Плавают личинки многотысячными стайками. Живыми лентами в метр шириной и несколько метров длиной вьются они в пруду. То у поверхности, то погружаются на дно. Все личинки разом совершают манёвры: плывут в одну сторону и в одном ритме .

Через несколько месяцев начинается метаморфоз, во время которого системы организма резко перестраиваются во взрослую форму. Как правило, метаморфоз длится около суток. Он начинается с выработки гормона тироксина , который влияет на развитие тканей. Так, изменение органов дыхания включает в себя развитие лёгких параллельно с исчезновением жабр и жаберного мешка. Передние конечности становятся видимыми. Нижняя челюсть приобретает характерную для хищника форму, кишечник укорачивается. Нервная система адаптируется под стереоскопическое зрение и слух, а также под новые способы передвижения и питания. Глаза сдвигаются выше, формируются веки и связанные с ними железы. Модифицируются органы слуха (появляется слуховая перепонка и среднее ухо). Кожа становится толще и крепче, боковая линия исчезает (у большинства видов), формируются кожные железы. На последнем этапе метаморфоза исчезает хвост, ткани которого идут на развитие конечностей . Постепенно жабята выходят на сушу. Заканчивается метаморфоз практически одновременно у всех головастиков .
В неволе зарегистрирована продолжительность жизни представителей семейства до 40 лет, этого возраста достигла обыкновенная жаба . Жаба-ага жила в неволе 24 года, жаба Бломберга — 28 лет, колорадская жаба — до 15 лет, а — 20 лет .
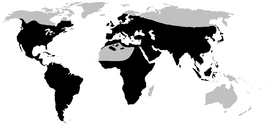
Распространены повсеместно, за исключением Арктики , Антарктики , Гренландии , Мадагаскара , Новой Зеландии и многих островов Тихого океана . В Австралии и Гвинее обитает ввезённая из Америки жаба-ага .
На территории России распространены лишь представители рода Bufo , всего 6 видов: обыкновенная , кавказская , дальневосточная , зелёная , камышовая и монгольская жабы .
На жаб охотятся многие хищники, в том числе цапли , ястребы , рыбы, крупные саламандры , змеи (в том числе ужи , полозы ), черепахи , более крупные лягушки, ежи , сороки , вороны , хищные жуки и сороконожки , еноты , скунсы , хорьки и другие. Опасны люди, посыпающие поля инсектицидами и ДДТ . В дельте Миссисипи эти химикаты практически полностью истребили, уничтожили обитающих там жаб; немногие уцелели и сумели приспособиться к ядовитым веществам .

Опасны также для жаб лягушкоедки, или люцилии. Лягушкоедка — это зелёная падальная муха, паразитирующая на лягушках и жабах. В период размножения летом мухи откладывают свои белые продолговатые яйца на спину или голову жаб и других амфибий. Хозяевами паразита являются: обыкновенная , камышовая , зелёная жабы . Вылупившиеся личинки проникают через носовые отверстия в голову хозяина, где въедаются в мягкие ткани ещё живущего животного. Через несколько дней передняя часть морды и головы может полностью деформироваться и исказиться. Когда личинки добираются до мозга или других жизненно важных органов, функции организма нарушаются и амфибия погибает. В своей последней фазе личинки почти полностью съедают труп животного, включая кожу и соединительные ткани, при определённых обстоятельствах — даже хрящи и кости .
Количество мутаций и генетических дефектов, наблюдаемых у жаб, как и у всех бесхвостых, возросло за время наблюдения с 1990-х по 2003 год. Одним из распространённых дефектов являются недостающие или лишние конечности. Различные гипотезы по поводу причин, вызывающих эти дефекты, включают в себя увеличение ультрафиолетового излучения, попадающего на икру, загрязнение сельскохозяйственными ядохимикатами и паразитические заболевания, как, например, заражение трематодами Ribeiroia ondatrae . Возможно, что все эти факторы действуют вместе (радиационный и химический стресс понижает сопротивление организмов паразитам). Дефекты конечностей ухудшают подвижность и, как следствие, шансы животного дожить до половой зрелости .
Самки комаров рода Corethrella питаются кровью бесхвостых амфибий, в том числе и жаб. Жертву они находят, ориентируясь по звуковым брачным сигналам. Самцы жаб издают сложные и простые позывные сигналы. Сложные сигналы предпочитаются, как самками комаров, так и самками лягушек. Если самцы издают простые звуки, они меньше подвергаются нападению кровососов, но и вероятность спаривания таких самцов уменьшается. Комары служат переносчиками трипаносомозов лягушек .
Специализированными хищниками на многих жаб являются жуки и их личинки рода в составе семейства жужелиц .

Исследования, начатые в 1950-х годах, указывают на значительное сокращение численности многих видов бесхвостых, в том числе и жаб. Среди этих видов числится и оранжевая жаба из Коста Рики . Её исчезновение вызвало серьёзную озабоченность исследователей, связанную с тем, что эта жаба обитала в заповеднике « » и её вымирание не могло быть вызвано напрямую человеческим вмешательством. В других местах снижение численности обусловлено разрушением среды обитания, поллютантами , климатическими изменениями и внедрением чужеродных хищников, паразитов и конкурентов. Особенно разрушительными для популяций жаб и вообще всех бесхвостых считаются инфекционные заболевания хитридиомикоз и .
В европейском фольклоре жабы ассоциировались с ведьмами как фамилиары , источники ингредиентов для магических зелий, а иногда имели собственные магические свойства . В иконографии жаба является атрибутом смерти или алчности . Кроме того, жаба, пожирающая гениталии обнаженной женской фигуры, символизирует похоть . Легенда Панамы гласит, что удача сопутствует тому, кто увидит панамскую золотую лягушку ( Atelopus zeteki ). В варианте этой легенды говорится, что, умирая, эти лягушки превращаются в золотой талисман уаку . В мультфильмах « Шрек 2 » и « Шрек 3 » жаба — истинный облик короля Гарольда.
На февраль 2023 года в семейство включают 52 рода и 642 видов :
Виды incertae sedis




| Жабы | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Обыкновенная жаба |
||||||||||||
| Научная классификация | ||||||||||||
|
Домен:
Царство:
Подцарство:
Без ранга:
Без ранга:
Тип:
Подтип:
Инфратип:
Надкласс:
Класс:
Подкласс:
Инфракласс:
Надотряд:
Отряд:
Подотряд:
Надсемейство:
Семейство:
Жабы
|
||||||||||||
| Международное научное название | ||||||||||||
| Bufonidae J. E. Gray , 1825 | ||||||||||||
| Ареал | ||||||||||||
|
||||||||||||
|
||||||||||||


















