Interested Article - Смилодоны

- 2020-12-06
- 1
Смилодоны ( лат. Smilodon , от др.-греч. σμίλη — нож, и ὀδούς , род. п. ὀδόντος — зуб) — род вымерших саблезубых кошек , представители которого жили с 2,5 млн до 10 тыс. лет назад в Северной и Южной Америке. Вероятно, произошёл от мегантереона . Широко известное название « » на самом деле является некорректным.
По данным изучения митохондриальной ДНК саблезубых кошек из подсемейства Machairodontinae , эволюционные пути родов Smilodon и Homotherium разошлись около 22 млн лет назад . Одной из причин, приведших саблезубых кошек к вымиранию, могло быть их низкое генетическое разнообразие , по другой версии, на вымирание смилодонов могло повлиять истребление первобытными людьми плейстоценовой мегафауны в ходе заселения Америки .
Описание


Смилодон был размером со льва или тигра , хотя имел куда более крепкое сложение и весил от 160—280 кг ( Smilodon fatalis ) до 220—400 кг ( Smilodon populator ), имел короткий хвост. Один из видов смилодона, Smilodon populator , являлся самым крупным представителем подсемейства саблезубых кошек . Смилодоны отличались от других представителей кошачьих мощным телосложением. Верхние клыки имели длину с корнями до 28 см, которые достигали верхнего края глазниц . Они были уплощены с боков, изогнуты и заострены с вогнутой стороны. Коренные зубы напротив были редуцированы.
Чтобы воспользоваться верхними клыками, смилодон мог открыть пасть на 120°, — для этого суставное соединение нижней челюсти и черепа было опущено. Ещё одна особенность смилодонов — развитие сосцевидного отростка, к которому прикреплялись мышцы, опускающие голову при нанесении удара верхними клыками, что является приспособлением, усиливающим это движение. Специализировались в охоте на очень крупных животных, например, мамонтов (вероятно, в основном молодых или детёнышей), мастодонтов , степных зубров , западных лошадей . Среда обитания — леса, где было проще подкрасться к жертве, но могли обитать также на открытых пространствах . Вероятно, охотились в одиночку из засады, подобно современным тиграм, так как короткие лапы и крепкое телосложение не позволяли им долго преследовать быструю добычу . Отсутствие доказательств того, что самцы были значительно крупнее самок, также может свидетельствовать об одиночном образе жизни или нахождению лишь в неорганизованных группах, с равным количеством самцов и самок, в отличие от обитающих в тех же регионах гомотериев , охотившихся группами (прайдами) на открытых пространствах .

У смилодона была уменьшенная поясничная область, высокая лопатка, короткий хвост и широкие конечности с относительно короткими ступнями . Смилодон наиболее известен своими относительно длинными клыками, которые являются самыми длинными среди саблезубых кошек, около 28 см (11 дюймов) в длину у самого крупного вида S. populator . Клыки были тонкими и имели тонкие зазубрины на передней и задней стороне . Череп был крепко сложен, морда короткая и широкая. Скуловые кости (zygomata) глубокие и широко изогнутые, сагиттальный гребень выражен, лобная область слегка выпуклая. Нижняя челюсть имела фланец с каждой стороны спереди. Верхние резцы были большими, острыми и наклонёнными вперёд. Между резцами и молярами нижней челюсти была диастема (щель). Нижние резцы были широкими, изогнутыми и располагались по прямой линии. Премолярный зуб p3 нижней челюсти присутствовал у большинства ранних экземпляров, но утрачен у более поздних экземпляров; он присутствовал только в 6 % пробы из Ла-Бреа . Идут споры о том, был ли у смилодона половой диморфизм: некоторые исследования окаменелостей S. fatalis не выявили особых различий между полами , в то время как исследование 2012 года показало, что, хотя окаменелости S. Fatalis демонстрируют меньшие различия в размерах, чем современные Panthera , они, по-видимому, демонстрируют такое же различие между полами в некоторых чертах .

S. gracilis был самым мелким видом, его вес оценивался в 55 кг. и он был размером с ягуара. Он был похож на своего предшественника Megantereon того же размера, но его зубной ряд и череп были более развитыми, приближаясь к S. fatalis. S. fatalis был промежуточным по размеру между S. gracilis и S. populator . Его вес варьировался от 160 до 280 кг , он достигал высоты в плечах в 100 см и имел длину тела в 175 см . Он был похож на льва размерами, но был более крепким и мускулистым, а потому имел большую массу тела. Его череп также был похож на череп Мегантереона, но более массивный и с более крупными клыками . S. populator был одним из самых крупных известных кошачьих с диапазоном массы тела от 220 до 400 кг , и, по одной из оценок, до 470 кг . Особенно большой череп S. populator из Уругвая размером 39 см в длину указывает на то, что эта особь могла весить до 436 кг. и достигать высоты в плечах до 120 см . По сравнению с S. fatalis, S. populator был более крепким и имел более удлинённый и узкий череп с более прямым профилем верхней части, более высоко расположенными носовыми костями, более вертикальным затылком, более массивными метаподиями и немного более длинными передними конечностями по сравнению с задними . Большие следы из Аргентины (для которых было предложено название ихнотаксона Smilodonchium) были отнесены к S. populator и имеют размеры 17,6 см на 19,2 см . Они оказались больше следов бенгальского тигра , с которым и было решено произвести сравнение .

Традиционно саблезубые кошки были художественно восстановлены с внешними чертами, похожими на черты существующих кошачьих, художниками, такими как Чарльз Р. Найт, в сотрудничестве с различными палеонтологами в начале XX века . В 1969 году палеонтолог Дж. Дж. Миллер вместо этого предположил, что смилодон сильно отличался бы от типичной кошки и был бы похож на бульдога, с более низкой линией губ (чтобы его рот мог широко открываться, не разрывая ткани морды), более втянутым носом и низкорасположенные уши . Палеохудожник Маурисио Антон и его соавторы оспаривали это в 1998 году и утверждали, что черты морды смилодона в целом не сильно отличались от черт других кошек. Антон отметил, что современные животные, такие как гиппопотам, за счёт умеренного сгибания круговой мышцы рта способны широко раскрывать рот, не разрывая ткани, и такая конфигурация мышц существует у современных крупных кошачьих . Антон заявил, что существующий филогенетический брекетинг (где черты ближайших существующих родственников ископаемого таксона используются в качестве эталона) является наиболее надёжным способом восстановления внешнего вида доисторических животных, и поэтому реставрации кошачьих смилодонов Найтом являются по-прежнему точными .
Смилодон и другие саблезубые кошки были реконструированы как с однотонной шерстью, так и с пятнистыми узорами (что, по-видимому, является наследственным условием для кошачьих), так как оба варианта окраса считаются возможными . Исследования современных видов кошек показали, что виды, живущие на открытом воздухе, как правило, имеют однородную шерсть, в то время как те, которые живут в более растительных средах обитания, имеют больше отметин, за некоторыми исключениями . Некоторые черты шерсти, такие как гривы львов-самцов или полосы тигра, слишком необычны, чтобы их можно было предсказать по окаменелостям .
Таксономия

В 1830-х годах датский натуралист Петер Вильхельм Лунд и его помощники собирали окаменелости в известняковых пещерах недалеко от небольшого городка Лагоа-Санта , штат Минас-Жерайс , Бразилия. Среди тысяч найденных окаменелостей он признал несколько изолированных щёчных зубов принадлежащими гиене, которую он назвал Hyaena neogaea в 1839 году — отдельный род кошачьих, хотя и переходный к гиенам . Он заявил, что по размеру он мог бы соответствовать самым крупным современным хищникам и был бы более крепким, чем любая современная кошка. Первоначально Лунд хотел назвать новый род Hyaenodon , но, понимая, что это название недавно получил другой доисторический хищник, в 1842 году назвал его Smilodon populator . Он объяснил древнегреческое значение Smilodon как σμίλη (улыбка), «скальпель» или « обоюдоострый нож», и οδόντος (odontús), «зуб». Это также было переведено как «зуб в форме обоюдоострого ножа». Он объяснил название вида populator как «разрушитель», что также переводится как «несущий опустошение». Животное получило название из-за выступающих верхних клыков, достигающих у некоторых видов 28 сантиметров. Крупные представители этого вида кошачьих могли весить до 400 кг при длине туловища немного больше одного метра. Учёные предполагают, что клыки помогали кошкам перегрызать у крупной добычи основные кровеносные сосуды, что вызывало у жертвы обильную кровопотерю, она быстро теряла силы . К 1846 году Лунд приобрёл почти все части скелета (от разных людей), и в последующие годы другими коллекционерами в соседних странах было найдено больше образцов . Хотя некоторые более поздние авторы использовали исходное название вида Лунда neogaea вместо populator , теперь оно считается недействительным nomen nudum , поскольку не сопровождалось надлежащим описанием и не обозначало типовых экземпляров . Некоторые экземпляры из Южной Америки были отнесены к другим родам, подродам, видам и подвидам, таким как Smilodontidion riggii , Smilodon (Prosmilodon) ensenadensis и S. bonaeriensis , но теперь они считаются младшими синонимами S. populator .

Окаменелости смилодона находились в Северной Америке начиная со второй половины XIX века . В 1869 году американский палеонтолог Джозеф Лейди описал фрагмент верхней челюсти с моляром , который был обнаружен в нефтяном пласте в округе Хардин , штат Техас . Он отнёс образец к роду Felis (который затем использовался для обозначения большинства кошек, как существующих, так и вымерших), но нашёл его достаточно отличным, чтобы быть частью своего собственного подрода, как F. (Trucifelis) fatalis . Видовое название означает «смертоносный» . В статье 1880 года о вымерших американских кошках американский палеонтолог Эдвард Дринкер Коп указал, что коренной зуб F. fatalis идентичен таковому у смилодона, и предложил новую комбинацию S. fatalis . Большинство находок в Северной Америке было скудным, пока не начались раскопки в карьерах Ла-Брея в Лос-Анджелесе, где с 1875 года были обнаружены сотни особей S. fatalis . У S. fatalis есть младшие синонимы, такие как S. mercerii, S. floridanus и S. californicus . Американский палеонтолог Аннализа Берта считала голотип S. fatalis слишком неполным, чтобы быть адекватным типовым образцом, и иногда этот вид предлагался в качестве младшего синонима S. populator . Скандинавские палеонтологи и поддержали различие двух видов в статье 1990 года . В статье от 2018 года американским палеонтологом Джоном П. Бабиарцем и его коллегами, сделан вывод о том, что S. californicus, представленный образцами из смоляных ям Ла-Бреа , в конце концов, был видом, отличным от S. fatalis, и что для уточнения таксономии необходимы дополнительные исследования рода .
В своей статье 1880 года о вымерших кошках Коуп также назвал третий вид смилодонов — S. gracilis . Этот вид был основан на частичном клыке, который был добыт в пещере Порт-Кеннеди, недалеко от реки Скулкилл в Пенсильвании . Коуп обнаружил, что клык отличается от других видов Smilodon из-за его меньшего размера и более сжатого основания . Его конкретное название связано с более лёгким телосложением этого вида , который известен из меньшего количества и менее полных останков, чем другие представители рода . S. gracilis иногда считался частью таких родов, как Megantereon и Ischyrosmilus . S. populator , S. fatalis и S. gracilis в настоящее время считаются единственными допустимыми видами Smilodon, а признаки, используемые для определения большинства их младших синонимов, были отвергнуты как различия между особями одного и того же вида (внутривидовая изменчивость) . Смилодон — одно из самых известных доисторических млекопитающих, часто упоминается в популярных СМИ и является окаменелостью штата Калифорния .
Останки крупнейших в истории представителей предков смилодона находились в Музее естественной и культурной истории Орегонского Университета . Они были исследованы в 2021 году учёными, которые определили их как новый вид (Machairodus lahayishupup) .
Эволюция
Самый известный саблезубый кот смилодон является одним из наиболее известных членов группы кошачьих, однако саблезубые хищники существовали не только у кошачьих. Термин «саблезубый» относится к экоморфе, состоящей из различных групп вымерших хищных синапсид (млекопитающих и близких родственников), у которых конвергентно развились чрезвычайно длинные верхнечелюстные клыки, а также связанных с этим адаптациям черепа и скелета. Сюда входят представители Gorgonopsia , , , Nimravidae , Barbourofelidae и Machairodontinae . Внутри семейства Felidae (настоящие кошки) члены подсемейства Machairodontinae называются саблезубыми кошками, и сама эта группа делится на три племени: (ложные саблезубые); (кошачьи саблезубые); и Smilodontini (саблезубые кошки), к которым принадлежит Smilodon . Представители Smilodontini отличаются своими длинными тонкими клыками с мелкими зазубринами или без них, тогда как Homotherini характеризуются более короткими, широкими и более уплощёнными клыками с более грубыми зазубринами .

Самые ранние представители кошачьих известны из олигоцена Европы, такие как Proailurus , а самый ранний вид с саблезубыми чертами — миоценовый род псевдаилурус . Морфология черепа и нижней челюсти самых ранних саблезубых кошек была похожа на морфологию современных дымчатых леопардов ( Neofelis ). Линия далее адаптировалась к точному убийству крупных животных за счёт развития удлинённых клыков и более широких промежутков, при этом жертвуя высокой силой укуса . По мере того, как их клыки становились длиннее, тела кошек становились более прочными для обездвиживания добычи . У производных смилодонтинов и гомотеринов поясничный отдел позвоночника и хвост стали укороченными, как и задние конечности . Основываясь на последовательностях митохондриальной ДНК, извлечённых из окаменелостей, линии Homotherium и Smilodon, могли разойтись примерно 18 млн лет назад . Самым ранним видом Smilodon является S. gracilis, который существовал от 2,5 миллионов до 500 000 лет назад (от ранней до ирвингтонской эпох) и был преемником Megantereon в Северной Америке, от которого он, вероятно, произошёл. Сам Megantereon проник в Северную Америку из Евразии в плиоцене вместе с Homotherium. S. gracilis достиг северных регионов Южной Америки в раннем плейстоцене в рамках великого межамериканского обмена . S. fatalis существовал 1,6 млн. — 10 000 лет назад (от позднего ирвингтонского до ранчолабрийского веков) и заменил S. gracilis в Северной Америке. S. populator существовал 1 млн. — 10 000 лет назад (от энсендана до лужанской эпохи) в восточной части Южной Америки .
Несмотря на разговорное название «саблезубый тигр», смилодон не имеет близкого родства с современным тигром (который принадлежит к подсемейству Pantherinae ) или любым другим существующим кошачьим . Анализ древней ДНК 1992 года показал, что смилодона следует отнести к современным кошкам (подсемейства Felinae и Pantherinae ).) . Исследование 2005 года показало, что смилодон принадлежал к отдельной линии кошачьих . Исследование, опубликованное в 2006 году, подтвердило это, показав, что Machairodontinae рано отделились от предков современных кошек и не были тесно связаны с какими-либо живыми видами . Следующая кладограмма 2011 года, основанная на окаменелостях и анализе ДНК, показывает место смилодона среди вымерших и существующих кошачьих согласно Ринкону и его коллегам :

| Felidae |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Развитие организма
У смилодона начинали развиваться взрослые саблезубые клыки, когда животное достигало возраста от 12 до 19 месяцев, вскоре после завершения прорезывания кошачьих молочных зубов . И детские, и взрослые клыки находились во рту в течение примерно 11 месяцев, а мышцы, используемые для мощного укуса, развивались примерно в полтора года (восемью месяцами ранее, чем у современного льва). Когда зверь вырастал до 23-30 месяцев, молочные зубы выпадали, а взрослые клыки продолжали расти со средней скоростью 7 мм (0,3″) в месяц в течение 12-месячного периода. Животное достигало своего полного размера примерно к 3 годам, что позже, чем у современных видов больших кошек. Молодые и юные ископаемые особи смилодонов чрезвычайно редко встречались на ранчо Ла-Бреа, где проводилось исследование, что указывает на то, что они скрывались или жили в берлогах во время охоты и зависели от родительской заботы, пока их клыки развивались .
Исследование 2017 года показывает, что молодые особи рождались с крепким телосложением, подобным взрослым. Сравнение костей молодых особей S. fatalis из Ла-Бреа с костями современного американского льва показало, что у двух кошек была схожая кривая роста. Развитие передних конечностей кошачьих в онтогенезе (изменения в процессе роста) была строго ограничена. Кривая похожа на кривую современных кошек, таких как тигры и пумы, но больше смещена в сторону сильного направления осей, чем у современных кошачьих . Исследования 2021 года Рейнольдса, Сеймура и Эванса показывают, что смилодон имел уникальную и быструю скорость роста, подобную тигру, но с длительным периодом созревания, подобно льву, в ходе которого детёныши зависели от своих родителей .
Палеобиология
Рацион

Будучи высшим хищником , смилодон в основном охотился на крупных млекопитающих. Изотопный состав костей S. fatalis в ямах Ла-Бреа-Тар показывает, что там кошки чаще всего ловили жвачных животных, таких как степные зубры ( , которые были намного крупнее современных американских бизонов) и верблюдов ( Camelops ) . Кроме того, изотопы, сохранившиеся в зубной эмали образцов S. gracilis из Флориды, показывают, что этот вид питался пекари Platygonus и ламоподобной . Изотопные исследования костей ужасного волка ( Aenocyon dirus ) и американского льва ( Panthera atrox ) показывают совпадение с S. fatalis в добыче, что позволяет предположить, что они были конкурентами . Однако более подробный изотопный анализ показывает, что Smilodon fatalis предпочитал лесную добычу, такую как тапиры , олени и лесные бизоны, в отличие от ужасных волков, предпочитающих добычу, обитающую на открытых территориях, таких как луга . Доступность добычи в районе Ранчо Ла Бреа, вероятно, была сопоставима с современной Восточной Африкой .
Когда смилодон мигрировал в Южную Америку, его рацион изменился: бизоны отсутствовали, лошади и хоботные были другими, а местные копытные, такие как токсодоны и литоптерны , были совершенно незнакомы; тем не менее S. populator процветал там так же хорошо, как и его родственники в Северной Америке . Изотопный анализ находок смилодона предполагает, что его основными видами-жертвами были , , Holmesina , виды рода Panochthus , , , и крокодил Caiman latirostris . Этот анализ его рациона также указывает на то, что S. populator охотился как в открытых, так и в лесных местообитаниях . Различия между видами Северной и Южной Америки могут быть связаны с разницей в добыче между двумя континентами . Смилодон, вероятно, избегал есть кости и оставлял достаточно еды для падальщиков . Смилодон мог поедать убитых ужасных волков . Было высказано предположение, что смилодоны были чистыми падальщиками, которые использовали свои клыки для разделки туш и отпугивания конкурентов, но эта теория сегодня не поддерживается, поскольку ни одно из современных наземных млекопитающих не является чистым падальщиком .
Хищническое поведение

Как и у современных кошек, мозг смилодона имел борозды , что предполагает повышенную сложность областей, которые контролируют слух, зрение и координацию конечностей. У кошачьих саблезубых в целом были относительно маленькие глаза, которые не были обращены вперёд, как у современных кошек, у которых хорошее бинокулярное зрение , помогающее им передвигаться по деревьям Smilodon вероятно, был засадным хищником, который скрывался в густой растительности, поскольку пропорции его конечностей были похожи на современных лесных кошек , а его короткий хвост не помогал бы ему балансировать во время бега . В отличие от своего предка Megantereon, который был, по крайней мере, частично пальцеходящим и, следовательно, мог лазить по деревьям, Smilodon , вероятно, был полностью наземным из-за своего большего веса и отсутствия приспособлений для лазания . Следы из Аргентины под названием Felipeda miramarensis в 2019 году могли принадлежать Smilodon . При правильной идентификации следы указывают на то, что у животного были полностью втягивающиеся когти, стопоходящая походка, а особенно крепкие передние конечности по сравнению с задними, указывают на то, что вероятно, это был засадный хищник .
Пяточная кость смилодона была довольно длинной, что говорит о том, что он был хорошим прыгуном . Его хорошо развитые мышцы-сгибатели и разгибатели на предплечьях, вероятно, позволяли ему тянуть вниз и надёжно удерживать крупную добычу. Анализ поперечных сечений бедренных костей S. fatalis показал, что кости были усилены за счёт утолщения надкостницы до такой степени, что могли выдерживать большую нагрузку, чем у современных больших кошек или у вымершего американского льва . Утолщение бедренных костей S. fatalis было в пределах диапазона существующих кошачьих . Его клыки были хрупкими и не могли впиться в кость; из-за риска поломки этим кошкам приходилось подавлять и удерживать свою добычу своими мощными передними конечностями, прежде чем они могли использовать свои клыки, и, вероятно, использовали быстрые рубящие или колющие укусы, а не медленные удушающие укусы, которые обычно используют современные кошки . В редких случаях, о чём свидетельствуют окаменелости, смилодон был готов рискнуть и вгрызться в кость своими клыками. Это могло быть больше направлено на борьбу с конкурирующими смилодонами и потенциальную угрозу со стороны других хищники, чем на добычу .

Продолжаются споры о том, как смилодон убивал свою добычу. Согласно самой популярной версии, кошка наносила глубокий колющий укус или колющий удар с открытой челюстью в горло, очень быстро убивая добычу . Другая гипотеза предполагает, что смилодон нацеливался на живот своей добычи. Это оспаривается, поскольку кривизна живота их жертв, вероятно, помешала бы кошке нанести хороший укус или удар . Что касается того, как смилодон наносит свой укус, предпочтение отдаётся гипотезе «собачьего укуса ножницами», согласно которой сгибание шеи и вращение черепа помогают укусить добычу, но это может быть механически невозможно. Однако данные сравнения с Homotherium показывают, что Smilodon был способен и использовал собачий укус ножницами в качестве основного средства убийства добычи. Эта версия основана на том факте, что у животного был толстый череп и относительно небольшая трабекулярная кость, в то время как у гомотерия была большая трабекулярная кость и он использовал зажимающий укус в качестве основного средства нападения. Опубликованное в 2020 году открытие, сделанное Фигейридо, Лаутеншлагером и другими предполагает чрезвычайно разные экологические адаптации у обоих махайродонтов . Фланцы нижней челюсти, возможно, помогали сопротивляться изгибающим силам, когда нижняя челюсть прижималась к шкуре жертвы .
Выступающие резцы были расположены дугой и использовались, чтобы удерживать добычу неподвижно и стабилизировать её во время нанесения собачьего укуса. Контактная поверхность между коронкой клыка и десной была увеличена, что помогло стабилизировать зуб и помогло кошке почувствовать, когда зуб проник на максимальную глубину. Поскольку у саблезубых кошек обычно было относительно большое подглазничное отверстие в черепе, в котором находились нервы, связанные с усами, было высказано предположение, что улучшенные чувства помогли бы кошкам точнее кусать вне их поля зрения, и тем самым предотвратить поломку клыков. Лезвиевидные хищные зубы использовались для разрезания кожи, чтобы получить доступ к мясу, а уменьшенные коренные зубы предположительно были менее приспособлены для дробления костей, чем у современных кошек . Так как пища современных кошек попадает в рот через бок при разрезании хищными, а не передними резцами между клыками, животным не нужно широко зевать, поэтому клыки смилодона также не были бы помехой при кормлении .
Несмотря на более мощное телосложение, чем у других крупных кошек, у смилодона был более слабый прикус. У современных крупных кошек скуловые дуги более выражены, в то время как у смилодона они были меньше, что ограничивало толщину и, следовательно, силу височных мышц и, таким образом, уменьшало силу укуса. Анализ его узких челюстей показывает, что он мог укусить только на треть сильнее льва (чей коэффициент силы укуса равен 112) . Кажется, существует общее правило, что у саблезубых кошек с самыми большими клыками пропорционально были более слабые укусы. Анализ прочности клыков на изгиб (способность клыков сопротивляться изгибающим усилиям без разрушения) и силы укуса показывает, что зубы саблезубых кошек были сильнее по отношению к силе укуса, чем у современных крупных кошек . Кроме того, зев смиолодона достигал 110 градусов , в то время как у современного льва он достигает 65 градусов . то сделало зев достаточно широким, чтобы смилодон мог схватить крупную добычу, несмотря на длинные клыки . Исследование 2018 года сравнило поведение хищника Smilodon fatalis и Homotherium и обнаружило, что у первого был крепкий череп с небольшой трабекулярной костью для колющего укуса, в то время как у второго была большая трабекулярная кость и он больше использовал стиль зажима и удержания, что делало его похожим на львов. Таким образом, эти двое заняли бы разные экологические ниши .
Останки
_(18246516009).jpg)
Многие экземпляры смилодонов были извлечены из смоляных ям , которые служили естественными ловушками для хищников. Животные случайно попали в ловушку и становились приманкой для хищников, которые затем сами оказались в ловушке. Самая известная из таких ловушек находится в Ла-Брея в Лос-Анджелесе, где было добыто более 166 тыс. экземпляров костей Smilodon fatalis что составляет самую большую коллекцию в мире. Отложения карьеров здесь накопились 40-10 тыс. лет назад, в . Хотя пойманные в ловушку животные быстро тонули, хищникам часто удавалось отгрызть у них кости конечностей, но они сами часто попадали в ловушку, а затем съедались другими хищниками; 90 % раскопанных костей принадлежали хищникам .
The Talara Tar Seeps представляют собой аналогичный сценарий, и в них также были обнаружены окаменелости смилодона. В отличие от Ла-Бреа, многие кости были сломаны или имеют признаки выветривания. Это могло быть связано с тем, что слои были более мелкими, поэтому избиение пойманных животных повредило кости ранее пойманных животных. Многие хищники в Таларе были молодыми, что, возможно, указывает на то, что неопытные и менее подготовленные животные имели больше шансов попасть в ловушку. Хотя Лунд считал, что скопление окаменелостей смилодона и травоядных в пещерах Лагоа-Санта произошло из-за того, что кошки использовали пещеры в качестве логова, это, вероятно, результат того, что животные умирали на поверхности, а водные потоки впоследствии утаскивают их кости на дно пещеры. Таким же образом могли погибнуть некоторые люди, заблудившись в пещерах .
Социальная жизнь

Учёные спорят, был ли смилодон социальным животным . Исследование африканских хищников показало, что социальные хищники, такие как львы и пятнистые гиены, больше реагируют на крики добычи, чем одиночные виды. Поскольку окаменелости S. fatalis часто встречаются в ямах Ла-Бреа-Тар и, вероятно, были привлечены криками бедствия застрявшей добычи, это может означать, что этот вид также был социальным . В одном критическом исследовании утверждается, что в исследовании игнорируются другие факторы, такие как масса тела (более тяжёлые животные чаще застревают, чем более лёгкие), интеллект (некоторые социальные животные, такие как американский лев, возможно, избегали смолы, потому что они лучше для распознавания опасности), отсутствие визуальных и обонятельных приманок, тип звуковой приманки и продолжительность сигналов бедствия (фактические сигналы бедствия пойманных в ловушку животных-жертв длились бы дольше, чем сигналы, использованные в исследовании). Автор этого исследования размышляет, как бы отреагировали хищники, если бы записи были воспроизведены в Индии, где, как известно, одинокие тигры собираются вокруг одной туши . Авторы первоначального исследования ответили, что, хотя эффекты вызовов в битумных ямах и экспериментов с воспроизведением не будут идентичными, этого недостаточно, чтобы опровергнуть их выводы. Кроме того, они заявили, что вес и интеллект вряд ли повлияют на результаты, поскольку более лёгкие хищники гораздо более многочисленны, чем тяжёлые травоядные, а социальный (и, по-видимому, умный) ужасный волк также встречается в ямах .

Другой аргумент в пользу общительности основан на заживших травмах в нескольких окаменелостях смилодона, что позволяет предположить, что животные нуждались в других, чтобы обеспечить их пищей . Этот аргумент был подвергнут сомнению, поскольку кошки могут быстро восстанавливаться даже после серьёзного повреждения костей, а раненый смилодон мог бы выжить, если бы у него был доступ к воде . Однако обнаружение останков страдавшего дисплазией тазобедренного сустава в молодом возрасте и дожившего до зрелых лет смилодона предполагает, что особь не могла бы прожить так долго без помощи социальной группы, поскольку животное из-за своей болезни не могло охотиться или защищать свою территорию . Мозг смилодона был относительно небольшим по сравнению с другими видами кошек. Некоторые исследователи утверждают, что мозг смилодона был бы слишком мал, чтобы он мог быть социальным животным . Анализ размера мозга у современных крупных кошек не выявил корреляции между размером мозга и социальностью . Ещё один аргумент против социального статуса смилодона основан на том, что, будучи предпочитающим охотиться из засады существом в закрытой среде обитания, ему, как и большинству современных кошек было бы ненужно жить в стае . Тем не менее, было также высказано предположение, что будучи самым крупным хищником в среде, сравнимой с саванной Африки, смилодон мог иметь социальную структуру, аналогичную современным львам, которые, возможно, живут группами в первую очередь для защиты оптимальной территории от других львов (львы являются единственными крупными социальными животными среди кошек) .
Половой диморфизм смилодона имеет значение для его репродуктивного поведения. Основываясь на своих выводах об отсутствии полового диморфизма у Smilodon fatalis, Ван Валкенбург и Сакко предположили в 2002 году, что, если бы кошки были социальными животными, они, вероятно, жили бы моногамными парами (вместе с потомством) без интенсивной конкуренции между самцами за самок . Точно так же Мичен-Самуэльс и Биндер в исследовании 2010 года пришли к выводу, что агрессия между самцами была менее выражена у S. fatalis, чем у американского льва . Кристиансен и Харрис в 2012 году обнаружили, что, поскольку S. fatalis действительно проявлял некоторый половой диморфизм, должен был иметь место эволюционный отбор для конкуренции между самцами . На некоторых костях видны следы укусов других смилодонов, возможно, в результате территориальных сражений, борьбы за право размножения или из-за добычи . На двух черепах S. populator из Аргентины видны кажущиеся смертельными незаживающие раны, которые, по-видимому, были нанесены клыками другого смилодона (хотя нельзя исключать, что они были нанесены ногой жертвы). Если это вызвано внутривидовыми драками, это может также указывать на то, что их социальное поведение могло привести к смерти, как это наблюдается у некоторых современных кошачьих (а также указывает на то, что клыки могут проникать в кости) . Было высказано предположение, что преувеличенные клыки саблезубых кошек эволюционировали для сексуальной демонстрации и драк между самцами, но статистическое исследование корреляции между клыком и размером тела у S. populator не обнаружило разницы в масштабировании между размером тела и клыком. Вероятно, они эволюционировали исключительно для хищнической функции .
Набор из трёх связанных скелетов S. fatalis, найденных в Эквадоре и описанных в 2021 году Рейнольдсом, Сеймуром и Эвансом, предполагает, что смилодон находился под длительной родительской заботой. У двух обнаруженных неполовозрелых особей есть уникальная унаследованная черта в их зубных костях, что позволяет предположить, что они были братьями и сёстрами; это редкий случай семейных отношений, обнаруженный в летописи окаменелостей. Также предполагается, что неполовозрелые экземпляры могли быть мужского и женского пола, в то время как останки взрослого скелета предположительно принадлежали их матери. По оценкам, несовершеннолетним на момент смерти было около двух лет, но их организмы всё ещё росли . Строение подъязычных костей предполагает, что смилодон мог рычать , как современные большие кошки, что может иметь значение для их социальной жизни .
Палеопатология
Несколько окаменелостей Smilodon демонстрируют признаки анкилозирующий спондилоартрит , гиперостоза и травмы . Исследование 1000 черепов смилодона показало, что у 36 % из них были разрушены теменные кости, к которым прикрепляются самые большие мышцы челюсти. У них также были признаки микропереломов, а также ослабление и истончение костей, возможно, вызванное механическим напряжением из-за постоянной необходимости делать колющие движения клыками . Костные разрастания в местах прикрепления дельтовидной мышцы к плечевой кости являются распространённой патологией для экземпляра из Ла Бреа, что, вероятно, было связано с повторяющимся напряжением, когда смилодон пытался стянуть добычу передними конечностями. Также распространены травмы грудины, вероятно, из-за столкновения с добычей . Частота травм у особей S. fatalis составила 4,3 % по сравнению с 2,8 % у ужасного волка, что означает, что нападения из засад приводило к большему риску травм, чем преследования. Останки смилодона демонстрируют относительно больше повреждений плечевых и поясничных позвонков .
Распространение и среда обитания

Смилодон жил в эпоху плейстоцена (2,5 млн — 10 000 лет назад) и, возможно, был самым последним из саблезубых кошек . Вероятно, он жил в закрытой среде обитания, такой как лес или кустарник . Окаменелости этого рода были найдены по всей Америке . Самые северные остатки рода — это окаменелости S. fatalis из Альберты , Канада . Ареал в Северной Америке варьировался от субтропических лесов и саванн на юге до безлесных мамонтовых степей на севере. Мозаичная растительность из лесов, кустарников и трав на юго-западе Северной Америки поддерживала существование крупных травоядных, таких как лошади, бизоны, антилопы, олени, верблюды, мамонты, мастодонты и наземные ленивцы . Северная Америка также поддерживала других саблезубых кошек, таких как Homotherium и Xenosmilus, а также других крупных хищников, включая ужасных волков, короткомордых медведей (Arctodus simus) и американских львов . Конкуренция со стороны таких плотоядных могла помешать североамериканскому S. fatalis достичь размера популяции S. Южной Америки. Сходство в размерах S. fatalis и американского льва предполагает перекрытие ниш и прямую конкуренцию между этими видами, и, по-видимому, они питались добычей одинакового размера .
S. gracilis попал в Южную Америку в период от раннего до среднего плейстоцена, где, вероятно, дал начало S. populator , обитавшему в восточной части континента. S. fatalis также проник в западную часть Южной Америки в позднем плейстоцене, и считалось, что эти два вида разделены Андами . Однако в 2018 году сообщалось о черепе S. fatalis, найденном в Уругвае к востоку от Анд, что ставит под сомнение идею о том, что эти два вида были аллопатрическими (географически разделёнными) . Американский обмен привёл к смешению местных и инвазивных видов, разделяющих прерии и леса в Южной Америке; Североамериканские травоядные включали хоботных , лошадей, верблюдовых и оленей, южноамериканские травоядные включали токсодонов, литоптерн, наземных ленивцев и глиптодонтов . Местные метатерийные хищники (включая саблезубых тилакосмилов ) вымерли к плиоцену и были заменены североамериканскими хищниками, такими как псовые, медведи и крупные кошки .
S. populator был очень успешным, тогда как Homotherium так и не получил широкого распространения в Южной Америке. Вымирание тилакосмилов связывают с конкуренцией со смилодонами, но это, вероятно, неверно, поскольку они, по-видимому, исчезли до появления крупных кошек. Фороракосовые , возможно, доминировали в нише крупных хищников в Южной Америке до появления смилодона . S. populator, возможно, смог достичь большего размера, чем S. fatalis, из-за отсутствия конкуренции в плейстоценовой Южной Америке; S. populator появился после исчезновения одного из крупнейших когда-либо существовавших плотоядных Arctotherium angustidens , и поэтому мог занять нишу мегахищников . S. populator предпочитал крупную добычу из открытых мест обитания, таких как пастбища и равнины, на основании данных, полученных из соотношений изотопов, которые определяли рацион животного. Таким образом, южноамериканский вид Smilodon, вероятно, был похож на современного льва. S. populator, вероятно, конкурировал там с псовым , но не с ягуаром, который питался в основном более мелкой добычей .
Вымирание

Наряду с большей частью мегафауны плейстоцена, смилодон вымер 10 000 лет назад в результате четвертичного вымирания . Его исчезновение было связано с сокращением и вымиранием крупных травоядных, которые были заменены более мелкими и проворными, такими как олени. Следовательно, смилодон мог быть слишком специализирован на охоте на крупную и медленную добычу и, возможно, не смог адаптироваться . Исследование износа зубов смилодона, проведённое в 2012 году, не обнаружило доказательств того, что они были ограничены пищевыми ресурсами . Другие объяснения включают изменение климата и конкуренцию с Homo sapiens ( пришедшими в Америку около 15 000 лет назад, незадолго до исчезновения смилодона) или сочетание нескольких факторов, все из которых относятся к общему вымиранию плейстоценовой мегафауны , а не конкретно к исчезновению саблезубых кошек .
Авторы первой половины XX в. предполагали, что последние саблезубые кошки в лице смилодона и гомотерии вымерли из-за конкуренции с пришедшими им на смену более быстрыми и более универсальными кошачьими. Было даже высказано предположение, что саблезубые хищники уступают современным кошкам, поскольку считалось, что постоянно растущие клыки мешают их владельцам правильно питаться. Тем не менее, быстрые кошачьи, такие как американский лев и американский гепард (Miracinonyx), также вымерли в позднем плейстоцене. Тот факт, что саблезубые зубы много раз появлялись в неродственных линиях, также свидетельствует об успехе этой функции .
Поздний образец Smilodon fatalis, извлечённый из смоляных ям Ранчо Ла Бреа, датируется 13 025 годами назад . Останки последней популяции смилодонов, найденные в пещере Куэва-дель-Медио, недалеко от города Сориа, на северо-востоке провинции Ультима-Эсперанса, регион Магалланес на самом юге Чили, датируются 10 935-11 209 лет назад . Самая последняя дата по углероду-14 для S. fatalis, о которой сообщалось, составляла 10 200 лет назад для останков из Первой американской пещеры в 1971 году ; однако самая последняя «достоверная» дата была указана как 11 130 лет назад .
Виды
- Smilodon fatalis — существовал 1 600 000—10 000 лет назад на территории Северной Америки
- — подвид Smilodon fatalis
- — подвид Smilodon fatalis
- Smilodon gracilis — жил в плейстоцене в обеих Америках
- Smilodon populator typus — наиболее крупный из всех смилодонов, достигал высоты в холке 120 см, 210 см длины тела, без учёта 30 см хвоста, и массы до 400 кг, кондило-базальная длина черепа 369 мм, ширина в скулах 233 мм, обитал в Южной Америке 1 000 000—10 000 лет назад.
Примечания
- / О. Л. Россолимо, И. Я. Павлинов , С. В. Крускоп, А. А. Лисовский, Н. Н. Спасская, А. В. Борисенко, А. А. Панютина. — М. : Издательство КМК, 2004. — Ч. III. — С. 725. — 408 с. — (Разнообразие животных). — ISBN 5-87317-098-3 .
- . ТАСС . Дата обращения: 14 июня 2021. 14 июня 2021 года.
- от 4 ноября 2019 на Wayback Machine , 2017
- от 16 августа 2019 на Wayback Machine , 2017
- Буровский А. М. // Биосфера. — 2010. — Т. 2 , вып. 1 . — ISSN . 29 апреля 2021 года.
- ↑ . Дата обращения: 19 июля 2011. 21 октября 2012 года.
- . Дата обращения: 19 июля 2011. Архивировано из 19 августа 2014 года.
- William J. Gonyea. (англ.) // Paleobiology. — 1976/ed. — Vol. 2 , iss. 4 . — P. 332—342 . — ISSN . — doi : . 14 июня 2021 года.
- Екатерина Русакова. . nplus1.ru . Дата обращения: 14 июня 2021. 14 июня 2021 года.
- Larisa R. G. DeSantis, Jonathan M. Crites, Robert S. Feranec, Kena Fox-Dobbs, Aisling B. Farrell. (англ.) // Current Biology. — 2019-08-05. — Vol. 29 , iss. 15 . — P. 2488—2495.e2 . — ISSN . — doi : .
- Екатерина Русакова. . nplus1.ru . Дата обращения: 14 июня 2021. 14 июня 2021 года.
- Ross Barnett, Michael V. Westbury, Marcela Sandoval-Velasco, Filipe Garrett Vieira, Sungwon Jeon. (англ.) // Current Biology. — 2020-12-21. — Vol. 30 , iss. 24 . — P. 5018—5025.e5 . — ISSN . — doi : .
- ↑ Turner, A. / A. Turner, M. Antón. — Columbia University Press, 1997. — P. 57–58, 67–68. — ISBN 978-0-231-10229-2 .
- University of California Museum of Paleontology. Дата обращения: 8 апреля 2013. 16 мая 2020 года.
- Slater, G. J.; Valkenburgh, B. V. (2008). "Long in the tooth: evolution of sabertooth cat cranial shape". Paleobiology . 34 (3): 403—419. doi : . ISSN .
- ↑ Berta, A. (1985). (PDF) . Contributions in Science, Natural History Museum of Los Angeles County . 370 : 1—15. (PDF) из оригинала 28 мая 2014 .
- ↑ Van Valkenburgh, B.; Sacco, T. (2002). "Sexual dimorphism, social behavior and intrasexual competition in large Pleistocene carnivorans". Journal of Vertebrate Paleontology . 22 (1): 164—169. doi : . JSTOR .
- ↑ Meachen-Samuels, J.; Binder, W. (2010). "Sexual dimorphism and ontogenetic growth in the American lion and sabertoothed cat from Rancho La Brea". Journal of Zoology . 280 (3): 271—279. doi : .
- ↑ Christiansen, Per; Harris, John M. (2012). . PLOS ONE . 7 (10): e48352. Bibcode : . doi : . PMC . PMID .
- ↑ Christiansen, Per; Harris, John M. (2005). "Body size of Smilodon (Mammalia: Felidae)". Journal of Morphology . 266 (3): 369—84. doi : . PMID .
- . San Diego Zoo Global (январь 2009). Дата обращения: 7 мая 2013. Архивировано из 3 февраля 2013 года.
- ↑ , pp. 108–154.
- Sorkin, B. (2008). "A biomechanical constraint on body mass in terrestrial mammalian predators". . 41 (4): 333—347. doi : .
- Manzuetti, A.; Perea, D.; Jones, W.; Ubilla, M.; Rinderknecht, A. (2020). "An extremely large saber-tooth cat skull from Uruguay (late Pleistocene–early Holocene, Dolores Formation): body size and paleobiological implications". Alcheringa: An Australasian Journal of Palaeontology . 44 (2): 332—339. doi : .
- ↑ Kurtén, B.; Werdelin, L. (1990). "Relationships between North and South American Smilodon ". Journal of Vertebrate Paleontology . 10 (2): 158—169. doi : . JSTOR .
- . 0223.com.ar (исп.) . 0223. 2016-05-26. из оригинала 22 июня 2018 . Дата обращения: 28 мая 2016 .
- Perkins, Sid (June 10, 2016). . Science . doi : . из оригинала 15 июля 2021 . Дата обращения: 8 апреля 2022 .
- ↑ , pp. 157–176.
- Miller, G. J. (1969). "A new hypothesis to explain the method of food ingestion used by Smilodon californicus Bovard". Tebiwa . 12 : 9—19.
- ↑ Antón, M.; García-Perea, R.; Turner, A. (1998). . Zoological Journal of the Linnean Society . 124 (4): 369—386. doi : .
- Allen, W. L.; Cuthill, I. C.; Scott-Samuel, N. E.; Baddeley, R. (2010). . Proceedings of the Royal Society B . 278 (1710): 1373—1380. doi : . PMC . PMID .
- Екатерина Русакова. . N + 1 . Дата обращения: 20 апреля 2022. 14 июня 2021 года.
- Максим Абаев. . Портал журнала «Наука и жизнь» . Дата обращения: 20 апреля 2022. 9 мая 2021 года.
- ↑ , pp. 3–26.
- Lund, P. W. : [ датск. ] . — Copenhagen : Det Kongelige Danske Videnskabernes Selskabs Naturvidenskabelige og Matematiske Afhandlinger, 1842. — P. 54–57.
- Leidy, J. (1869). . Journal of the Academy of Natural Sciences of Philadelphia . 7 : 366—367. doi : . из оригинала 2 февраля 2021 . Дата обращения: 6 апреля 2022 .
- . National Geographic (21 сентября 2011). Дата обращения: 13 октября 2021. 29 октября 2021 года.
- ↑ Cope, E. D. (December 1880). . The American Naturalist . 14 (12): 833—858. doi : . JSTOR .
- Werdelin, L. : [ англ. ] / L. Werdelin, H. G. McDonald, C. A. Shaw. — Smilodon from South Carolina: Implications for the taxonomy of the genus. — Baltimore : Johns Hopkins University Press, 2018. — P. 76–95. — ISBN 978-1-4214-2557-3 .
- Kurtén, B. / B. Kurtén, E. Anderson. — New York : Columbia University Press, 1980. — P. 186–188. — ISBN 978-0-231-03733-4 .
- Berta, A. (1987). . Bulletin of the Florida State Museum . 31 : 1—63. из оригинала 26 ноября 2015 . Дата обращения: 6 апреля 2022 .
- Churcher, C. S. . — , 1984. — Vol. 140. — P. . — ISBN 978-0-88854-305-9 . — doi : .
- . Naked Science . Дата обращения: 20 апреля 2022. 19 мая 2022 года.
- Orcutt, J.D., Calede, J.J. (англ.) // Journal of Mammalian Evolution. — 2021. — No. 28 . — P. 729–751 . — doi : . 1 июня 2021 года.
- Meehan, T.; Martin, L. D. J. (2003). "Extinction and re-evolution of similar adaptive types (ecomorphs) in Cenozoic North American ungulates and carnivores reflect van der Hammen's cycles". Die Naturwissenschaften . 90 (3): 131—135. Bibcode : . doi : . PMID .
- ↑ , pp. 65–76.
-
Christiansen, P. (2008).
.
PLOS ONE
.
3
(7): e2807.
Bibcode
:
.
doi
:
.
PMC
.
PMID
.
- Paijmans, J. L. A.; Barnett, R.; Gilbert, M. T. P.; Zepeda-Mendoza, M. L.; Reumer, J. W. F.; de Vos, J.; Zazula, G.; Nagel, D.; Baryshnikov, G. F.; Leonard, J. A.; Rohland, N.; Westbury, M. V.; Barlow, A.; Hofreiter, M. (2017-10-19). . Current Biology . 27 (21): 3330—3336.e5. doi : . PMID .
- ↑ Rincón, A.; Prevosti, F.; Parra, G. (2011). "New saber-toothed cat records (Felidae: Machairodontinae) for the Pleistocene of Venezuela, and the Great American Biotic Interchange". Journal of Vertebrate Paleontology . 31 (2): 468—478. doi : . JSTOR .
- de Castro, Mariela Cordeiro; Langer, Max Cardoso (2008). . Revista Brasileira de Paleontologia . 11 (3): 199—206. doi : .
- University of California Museum of Paleontology (декабрь 2005). Дата обращения: 12 июня 2012. 16 мая 2020 года.
- Janczewski, D. N.; Yuhki, N.; Gilbert, D. A.; Jefferson, G. T.; O'Brien, S. J. (1992). . Proceedings of the National Academy of Sciences . 89 (20): 9769—9773. Bibcode : . doi : . PMC . PMID .
- Barnett, R.; Barnes, I.; Phillips, M. J.; Martin, L. D.; Harington, C. R.; Leonard, J. A.; Cooper, A. (2005). . Current Biology . 15 (15): R589—R590. doi : . PMID .
- van den Hoek Ostende, L. W.; Morlo, M.; Nagel, D. (2006). "Majestic killers: the sabre-toothed cats (Fossils explained 52)". Geology Today . 22 (4): 150—157. doi : .
- . (1 июля 2015). Дата обращения: 2 июля 2015. 2 июля 2015 года.
-
Mihlbachler, M. C.; Wysocki, M. A.; Feranec, R. S.; Tseng, Z. J.; Bjornsson, C. S. (2015-07-01).
.
PLOS ONE
.
10
(7): e0129847.
Bibcode
:
.
doi
:
.
PMC
.
PMID
.
- Feranec, R. C. (2004). "Isotopic evidence of saber-tooth development, growth rate, and diet from the adult canine of Smilodon fatalis from Rancho La Brea". Palaeogeography, Palaeoclimatology, Palaeoecology . 206 (3—4): 303—310. Bibcode : . doi : .
- Long, K.; Prothero, D.; Madan, M.; Syverson, V. J. P.; Smith, T. (2017). . PLOS ONE . 12 (9): e0183175. Bibcode : . doi : . PMC . PMID .
- ↑ Reynolds, Ashley R.; Seymour, Kevin L.; Evans, David C. (January 7, 2021). . iScience . 24 (1): 101916. Bibcode : . doi : . PMC . PMID .
- ↑ Coltrain, J. B.; Harris, J. M.; Cerling, T. E.; Ehleringer, J. R.; Dearing, M.-D.; Ward, J.; Allen, J. (2004). (PDF) . Palaeogeography, Palaeoclimatology, Palaeoecology . 205 (3—4): 199—219. Bibcode : . doi : . Архивировано из (PDF) 29 мая 2013 .
- Fennec, R. S. (2005). (PDF) . Bulletin of the Florida Museum of Natural History . 45 (4): 369—377. (PDF) из оригинала 14 апреля 2021 . Дата обращения: 7 апреля 2022 – via University of Florida.
- DeSantis, Larisa R. G.; Crites, Jonathan M.; Feranec, Robert S.; Fox-Dobbs, Kena; Farrell, Aisling B.; Harris, John M.; Takeuchi, Gary T.; Cerling, Thure E. (2019). . Current Biology . 29 (15): 2488—2495. doi : . PMID .
- Vanvalkenburgh, B.; Hertel, F. (1993). "Tough times at la brea: tooth breakage in large carnivores of the late Pleistocene". Science . 261 (5120): 456—459. Bibcode : . doi : . PMID .
- Dantas, Mário André Trindade; Cherkinsky, Alexander; Lessa, Carlos Micael Bonfim; Santos, Luciano Vilaboim; Cozzuol, Mario Alberto; Omena, Érica Cavalcante; Silva, Jorge Luiz Lopes; Sial, Alcides Nóbrega; Bocherens, Hervé (2020-07-14). . Revista Brasileira de Paleontologia (англ.) . 23 (2): 138—152. doi : . ISSN . Архивировано из 16 июля 2020 . Дата обращения: 7 апреля 2022 .
- Van Valkenburgh, B.; Teaford, M. F.; Walker, A. (1990). . Journal of Zoology . 222 (2): 319—340. doi : .
- Van Valkenburgh, B. (1991). . Paleobiology . 17 (4): 340—362. doi : . JSTOR .
- ↑ , pp. 176–216.
- Gonyea, W. J. (1976). . Paleobiology . 2 (4): 332—342. doi : . JSTOR .
- Berkeley.edu. Дата обращения: 1 июня 2017. 16 мая 2020 года.
- ↑ Sabertooth. — 2013.
- Agnolin, F. L.; Chimento, N. R.; Campo, D. H.; Magnussen, M.; Boh, D.; De Cianni, F. (2019). "Large Carnivore Footprints from the Late Pleistocene of Argentina". Ichnos . 26 (2): 119—126. doi : .
-
↑
Meachen-Samuels, J. A.; Van Valkenburgh, B. (2010).
.
PLOS ONE
.
5
(7): e11412.
Bibcode
:
.
doi
:
.
ISSN
.
PMC
.
PMID
.
- McHenry, C. R.; Wroe, S.; Clausen, P. D.; Moreno, K.; Cunningham, E. (2007). . PNAS . 104 (41): 16010—16015. Bibcode : . doi : . PMC . PMID .
- Anyonge, W. (1996). . Journal of Mammalogy . 77 (4): 1059—1067. doi : . JSTOR .
- Figueirido, B.; Lautenschlager, S.; Pérez-Ramos, A.; Van Valkenburgh, B. (2018). . Current Biology . 28 (20): 3260—3266.e3. doi : . PMID .
-
Macchiarelli, R.; Brown, J. G. (2014).
.
PLOS ONE
.
9
(10): e107456.
Bibcode
:
.
doi
:
.
ISSN
.
PMC
.
PMID
.
- Hecht, J. (1 October 2007). . New Scientist . из оригинала 23 мая 2015 . Дата обращения: 6 апреля 2022 .
-
Wroe, S.; McHenry2, C.; Thomason, J. (2004).
(PDF)
.
Proceedings of the Royal Society
.
272
(1563): 619—25.
doi
:
.
PMC
.
PMID
. Архивировано из
(PDF)
25 августа 2013
.
{{ cite journal }}: Википедия:Обслуживание CS1 (числовые имена: authors list) ( ссылка ) - Christiansen, P. (2007). . Zoological Journal of the Linnean Society . 151 (2): 423—437. doi : .
-
Andersson, K.; Norman, D.; Werdelin, L. (2011).
.
PLOS ONE
.
6
(10): e24971.
Bibcode
:
.
doi
:
.
PMC
.
PMID
.
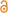
- Martin, L. D. (1980). . Transactions of the Nebraska Academy of Sciences . 8 : 141—154. из оригинала 3 декабря 2013 . Дата обращения: 6 апреля 2022 .
- Figueirido, B.; Lautenschlager, S.; Pérez-Ramos, A.; Van Valkenburgh, B. (2018). . Current Biology . 28 (20): 3260—3266.e3. doi : . PMID . из оригинала 20 апреля 2021 . Дата обращения: 6 апреля 2022 .
- Shaw, Christopher A.; Quinn, James P. (September 15, 2015). John M. Harris (ed.). (PDF) . Science Series 42: Contributions in Science . Natural History Museum of Los Angeles County (A special volume entitled La Brea and Beyond: the Paleontology of Asphalt–Preserved Biotas in commemoration of the 100th anniversary of the Natural History Museum of Los Angeles County's excavations at Rancho La Brea): 91—95. Архивировано из (PDF) 25 декабря 2016 . Дата обращения: 30 сентября 2017 .
- ↑ , pp. 30–33.
- Carbone, C.; Maddox, T.; Funston, P. J.; Mills, M. G. L.; Grether, G. F.; Van Valkenburgh, B. (2009). . Biology Letters . 5 (1): 81—85. doi : . PMC . PMID .
- Kiffner, C. (2009). . Biology Letters . 5 (4): 561—562. doi : . PMC . PMID .
- Van Valkenburgh, B.; Maddox, T.; Funston, P. J.; Mills, M. G. L.; Grether, G. F.; Carbone, C. (2009). . Biology Letters . 5 (4): 563—564. doi : . PMC .
- Heald, F. (1989). " "Injuries and diseases in Smilodon californicus Bovard, 1904, (Mammalia, Felidae) from Rancho La Brea, California". Journal of Vertebrate Paleontology . 9 (3): 24A.
- ↑ McCall, S.; Naples, V.; Martin, L. (2003). . Brain, Behavior and Evolution . 61 (3): 159—164. doi : . PMID . из оригинала 18 декабря 2019 . Дата обращения: 7 апреля 2022 .
- Balisi, Mairin A.; Sharma, Abhinav K.; Howard, Carrie M.; Shaw, Christopher A.; Klapper, Robert; Lindsey, Emily L. (2021). . bioRxiv . 11 (1): 21271. doi : . PMID . S2CID . из оригинала 7 апреля 2022 . Дата обращения: 7 апреля 2022 .
- Radinsky, L. B. (1975). "Evolution of the felid brain". Brain, Behavior and Evolution . 11 (3—4): 214—254. doi : . PMID .
- Yamaguchi, N.; Kitchener, A. C.; Gilissen, E.; MacDonald, D. W. (2009). . Biological Journal of the Linnean Society . 98 (1): 85—93. doi : .
- Chimento, N. R.; Agnolin, F. L.; Soibelzon, L.; Ochoa, J. G.; Buide, V. (2019). . Comptes Rendus Palevol . 18 (4): 449—454. doi : .
- O’Brien, D. M (2019). "Static scaling and the evolution of extreme canine size in a saber-toothed cat (Smilodon fatalis)". Integrative and Comparative Biology . 59 (5): 1303—1311. doi : . PMID .
- Hearld, F. The Great Cats / F. Hearld, C. Shaw. — Rodale Pr, 1991. — P. 26. — ISBN 978-1-875137-90-9 .
- Pickrell, J. . Scientific American (2018). Дата обращения: 29 октября 2018. 22 октября 2018 года.
- Bjorkengren, A. G.; Sartoris, D. J.; Shermis, S.; Resnick, D. (1987). . American Journal of Roentgenology . 148 (4): 779—782. doi : . PMID .
- Duckler, G. L. (1997). "Parietal depressions in skulls of the extinct saber-toothed felid Smilodon fatalis : evidence of mechanical strain". Journal of Vertebrate Paleontology . 17 (3): 600—609. doi : .
- Brown, C.; Balisi, M.; Shaw, C. A.; Van Valkenburgh, B. (2017). "Skeletal trauma reflects hunting behaviour in extinct sabre-tooth cats and dire wolves". Nature Ecology & Evolution . 1 (5): 0131. doi : . PMID .
- Meloro, C.; Elton, S.; Louys, J.; Bishop, L. C.; Ditchfield, P. (2013). (PDF) . Paleobiology . 39 (3): 323—344. doi : . (PDF) из оригинала 19 января 2022 . Дата обращения: 8 апреля 2022 .
- Reynolds, A. R.; Seymour, K. L.; Evans, D. C. (2019). "Late Pleistocene records of felids from Medicine Hat, Alberta, including the first Canadian record of the sabre-toothed cat Smilodon fatalis". Canadian Journal of Earth Sciences . 56 (10): 1052—1060. Bibcode : . doi : . : .
- DeSantis, L.R.G.; Schubert, B.W.; Schmitt-Linville, E.; Ungar, P.; Donohue, S.; Haupt, R.J. (September 15, 2015). John M. Harris (ed.). (PDF) . Science Series 42: Contributions in Science . Natural History Museum of Los Angeles County (A special volume entitled La Brea and Beyond: the Paleontology of Asphalt–Preserved Biotas in commemoration of the 100th anniversary of the Natural History Museum of Los Angeles County's excavations at Rancho La Brea): 37—52. Архивировано из (PDF) 20 декабря 2016 . Дата обращения: 6 февраля 2017 .
- ↑ Sherani, S. (2016). A new specimen-dependent method of estimating felid body mass (No. e2327v2) . PeerJ Preprints.
- Manzuetti, A.; Perea, D.; Ubilla, M.; Rinderknecht, A. (2018). "First record of Smilodon fatalis Leidy, 1868 (Felidae, Machairodontinae) in the extra-Andean region of South America (late Pleistocene, Sopas Formation), Uruguay: Taxonomic and paleobiogeographic implications". Quaternary Science Reviews . 180 : 57—62. Bibcode : . doi : .
- Bocherens, H.; Cotte, M.; Bonini, R.; Scian, D.; Straccia, P.; Soibelzon, L.; Prevosti, F. J. (2016-04-24). "Paleobiology of sabretooth cat Smilodon population in the Pampean Region (Buenos Aires Province, Argentina) around the Last Glacial Maximum: Insights from carbon and nitrogen stable isotopes in bone collagen". Palaeogeography, Palaeoclimatology, Palaeoecology . 449 : 463—474. Bibcode : . doi : .
- Hays, B. . UPI Science News . UPI (21 марта 2016). Дата обращения: 19 апреля 2016. 24 апреля 2016 года.
-
↑
DeSantis, L. R. G.; Schubert, B. W.; Scott, J. R.; Ungar, P. S. (2012).
.
PLOS ONE
.
7
(12): e52453.
Bibcode
:
.
doi
:
.
PMC
.
PMID
.
- ↑ , pp. 217–230.
- O'Keefe, F.R.; Fet, E.V.; Harris, J.M. (2009). "Compilation, calibration, and synthesis of faunal and floral radiocarbon dates, Rancho La Brea, California". Contributions in Science . 518 : 1—16. doi : .
- Prieto, Alfredo; Labarca, Rafael; Sierpe, Víctor (2010). . Revista Chilena de Historia Natural . 83 (2). doi : .
- (8 июля 2011). Дата обращения: 9 апреля 2022. 4 апреля 2022 года.
- Fiedel, Stuart. American Megafaunal Extinctions at the End of the Pleistocene. — Springer , 2009. — P. 21–37. — ISBN 978-1-4020-8792-9 . — doi : .
Литература
- Быстров А. П. // Природа. — М. : Издательство АН СССР, 1950. — № 12 — С. 30—37.
- Antón, M. . — 1st. — Bloomington : Indiana University Press, 2013. — ISBN 978-0-253-01042-1 .
- Christiansen P., Harris M. Body Size of Smilodon (Mammalia: Felidae) // Journal of Morphology. — 2005. — Vol. 266. — P. 369—384.
Ссылки
- .
|
Саблезубые хищники
|
|
|---|---|
| Горгонопсы | |
| Нимравиды | |
| Саблезубые кошки | |
| Другие саблезубые хищники | |
- 2020-12-06
- 1
| † Смилодоны | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

Smilodon fatalis , реконструкция |
||||||||||||||||||||||||||||
| Научная классификация | ||||||||||||||||||||||||||||
|
Домен:
Царство:
Подцарство:
Без ранга:
Без ранга:
Тип:
Подтип:
Инфратип:
Надкласс:
Клада:
Клада:
Класс:
Подкласс:
Клада:
Инфракласс:
Магнотряд:
Надотряд:
Клада:
Клада:
Грандотряд:
Отряд:
Подотряд:
Семейство:
Подсемейство:
Триба:
Род:
†
Смилодоны
|
||||||||||||||||||||||||||||
| Международное научное название | ||||||||||||||||||||||||||||
| Smilodon Lund, 1842 | ||||||||||||||||||||||||||||
| Виды | ||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||
|
Геохронология
2,5—0,01 млн лет
◄
Наше время
◄
Мел-палеогеновое вымирание
|
||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||

