Переходная новая кора
- 1 year ago
- 0
- 0


Зрительная кора является частью коры больших полушарий головного мозга , отвечающей за обработку визуальной информации. В основном, она сосредоточена в затылочной доле каждого из полушарий головного мозга .
Понятие зрительная кора включает первичную зрительную кору (также называемую стриарной корой или зрительной зоной V1 ) и экстрастриарную зрительную кору — зоны V2 , V3 , V4 , и V5 . Первичная зрительная кора анатомически эквивалентна полю Бродмана 17 , или BA17. Экстрастриарная зрительная кора включает и .
Именно со зрительной зоной V1 коры большого мозга непосредственно соединены нейроны, посылающие зрительные сигналы от глаз. Все остальные чувствительные к визуальным сигналам участки мозга (к 2000 году их было идентифицировано более 30) связаны с глазами через зону V1.
Зрительная кора присутствует в каждом из полушарий головного мозга . Области зрительной коры левого полушария получают сигналы от правой половины зрительного поля , правого полушария — от левой половины.
Далее в статье говорится об особенностях зрительной коры приматов (в основном, человека ).
Первичная зрительная кора (зона V1), по своему строению относящаяся к гранулярному (сенсорному) типу (иными словами, ), локализована в (зрительной) борозде ( англ. calcarine fissure ) затылочной доли ( англ. occipital lobe ) большого мозга. Первичная зрительная кора (зона V1) каждого полушария получает информацию непосредственно от наружного коленчатого тела .
Зрительные зоны V1 (правая и левая) передают визуальную информацию по двум первичным зрительным путям — дорсальному и вентральному (см. Рис. 3).
Разветвление потока зрительной информации на дорсальный ( «где?» ) и вентральный ( «что?» ) пути — иначе говоря, на каналы «действия» и « распознавания » — впервые описали и Мортимер Мишкин , и до сих пор эта гипотеза вызывает споры среди физиологов и учёных, изучающих феномен зрения . Возможно, она чрезмерно упрощает реальные процессы в зрительной коре. Она основана на обнаружении того, что оптические обманы зрения, такие как иллюзия Эббингауза , могут искажать правильное восприятие, но в случаях, когда субъект отвечает на визуальный стимул действием, таким как схватывание увиденного предмета, изображение воспринимается без искажений. Однако имеется публикация 2005 г. , утверждающая, что обе кортикальные системы обработки зрительной информации — и «канал действия» , и «канал распознавания» — в равной степени подвержены обману иллюзий.
Нейроны зрительной коры генерируют потенциал действия , когда визуальные стимулы воздействуют на их рецептивные поля . Рецептивное поле определяется как область , стимуляция которой приводит к генерации потенциала действия . Но иногда нейрон может лучше отвечать на какую-то определённую совокупность зрительных стимулов, представляемых в пределах его рецептивного поля. Это свойство называется . В более древних зрительных областях избирательность нейронов невысока. Так, нейрон зрительной зоны V1 может возбуждаться в ответ на любой вертикальный стимул в своём рецептивном поле . Нейронам интегративных зрительных областей свойственна сложная избирательность. Например, нейроны зрительной интегративной области верхней височной борозды (у обезьян) или вентральной поверхности на границе между затылочной и височной долями (у человека) могут возбуждаться лишь при стимуляции рецептивного поля изображениями лиц .
В обработку и восприятие визуальной информации о лице вовлечены несколько взаимодействующих друг с другом областей мозга. Ядро этой системы составляют: область в (OFA) , обеспечивающая начальный анализ отдельных частей лица; область в веретеновидной извилине (FFA), проводящая анализ инвариантных характеристик лица и узнавание человека по лицу ; область в задней части верхней височной борозды (pSTS), активирующаяся при анализе изменчивых аспектов — выражения лица, движений губ при речи и направления взгляда . В расширенной системе происходит дальнейший анализ направления взгляда ( — IPS), семантики ( нижняя лобная извилина — IFG, — ATC), эмоциональной составляющей ( миндалина — Amy, — Ins), биографической ( — PreCun, — pCiG) и другой информации. Связанная с восприятием объектов, (LOC), может включаться в ранний анализ структуры изображения лица. При этом выявление различных аспектов визуальной информации о лице осуществляется не автономной работой отдельных областей мозга, реализующих конкретные функции, а взаимосвязанной скоординированной их работой.
Кровоснабжение зрительной коры осуществляется, в основном, шпорной ветвью ( лат. ramus calcarinus ) — разветвления конечной (корковой) части ( лат. pars corticalis ) задней мозговой артерии ( лат. arteria cerebri posterior ). Шпорная ветвь залегает в шпорной борозде ( лат. sulcus calcarinus, fissura calcarina ) коры головного мозга .
При изучении первичной зрительной коры животных ( кошек , хорьков , крыс , мышей , обезьян ) регистрируются потенциалы , снимаемые с электродов , введённых в мозг, либо внутренние оптические сигналы ( коры мозга). Для исследования первичной зрительной коры человека и обезьян (зона V1) применяются электроэнцефалография (ЭЭГ), магнитоэнцефалография (МЭГ), функциональная магнитно-резонансная томография (фМРТ), позитронно-эмиссионная томография (ПЭТ).
Одно из современных открытий, касающихся зрительной зоны V1 человека, состоит в том, что внимание оказывает сильное модулирующее воздействие на обработку зрительной информации (сигналы были измерены посредством фМРТ ). В то же время, результаты физиологического исследования на макаках показали несущественное влияние процессов внимания на активность областей зрительной коры (очень малые изменения активности или их отсутствие). Эксперименты на макаках, как правило, выполняются путём регистрации пиковой активности единичных нейронов, а при использовании фМРТ , в основном, регистрируются постсинаптические потенциалы . Таким образом, контрастирующие результаты указанных экспериментов могут быть следствием неодинаковой методики исследований и вовсе не обязательно отражают реальные различия в физиологии человека и макаки.
Авторы другой современной публикации стремятся всецело охарактеризовать регуляторные свойства первичной зрительной коры (V1) и использовать её как модельную область, образец для характеристики кортикального канала передачи данных.
Повреждения первичной зрительной коры, как правило, приводят к возникновению скотомы , или «бреши» в зрительном поле. Интересно, что пациенты с такими дефектами зачастую могут воспринимать и использовать визуальную информацию, представленную по их скотомам. Это явление, называемое слепым ви́дением , или «слепым зрением», изучается многими учёными, проявляющими интерес к тому, какие именно структуры мозга и нейрофизиологические процессы являются «материальными носителями» сознания.

Первичная зрительная кора — наиболее изученная зрительная зона мозга . Исследования показали, что у млекопитающих она занимает задний полюс затылочной доли каждого полушария (эти доли ответственны за обработку зрительных стимулов). Это наиболее просто устроенная и филогенетически более «древняя» из кортикальных зон, связанных со зрением. Она приспособлена для обработки информации о статических и движущихся объектах, в особенности, для распознавания простых образов .
Составная часть функциональной архитектуры коры больших полушарий головного мозга — первичная зрительная кора — практически полностью соответствует анатомически определяемой стриарной коре . Название последней восходит к латинскому «полоса, полоска» ( лат. stria ) и во многом обусловлено тем, что здесь отчётливо видна невооружённым глазом полоска Дженнари (наружная полоска Байярже ), образованная конечными отделами покрытых миелиновой оболочкой аксонов , отходящих от нейронов латерального коленчатого тела и заканчивающихся в IV слое серого вещества .
Первичная зрительная кора подразделяется на шесть функционально различающихся горизонтальных цитоархитектонических слоёв , обозначаемых римскими цифрами от I до VI .
Слой IV ( внутренний зернистый слой ), к которому подходит наибольшее количество афферентных волокон, идущих от латеральных коленчатых тел (ЛКТ), в свою очередь, подразделяется на четыре подслоя, обозначаемых IVA , IVB , IVCα и IVCβ . Нервные клетки подслоя IVCα , в основном, получают сигналы, идущие от нейронов магноцеллюлярных («крупноклеточных», вентральных ) слоёв ЛКТ (« магноцеллюлярный зрительный путь »), подслоя IVCβ — от нейронов парвоцеллюлярных («мелкоклеточных», дорсальных ) слоёв ЛКТ (« парвоцеллюлярный зрительный путь »).
Полагают, что среднее число нейронов первичной зрительной коры взрослого человека составляет около 140 миллионов в каждом полушарии .
Зрительная область V2 , или вторичная зрительная кора , также называемая престриатной корой , — следующая крупная область зрительной коры и первый регион в пределах зрительной ассоциативной зоны . Зрительная область V2 имеет устойчивые упреждающие (англ.- feedforward) и обратные (англ.- feedback) связи с первичной зрительной корой V1. Область V2 также имеет устойчивую связь с областями V3, V4, и V5.
Что касается анатомии, V2 разделен на четыре квадранта: дорсальное и вентральное представление в левом и правом полушариях. Вместе эти четыре региона обеспечивают полную карту визуального мира. V2 имеет много общих свойств с V1: клетки настроены на простые свойства, такие как ориентация, пространственная частота и цвет. Ответы многих нейронов V2 также модулируются более сложными свойствами, такими как ориентация , диспаратность , и является ли стимул частью фигуры или фона. Недавние исследования показали, что клетки V2 демонстрируют небольшое количество модуляции внимания (больше, чем V1, меньше, чем V4), настроены на умеренно сложные структуры и могут управляться множественными ориентациями в разных субрегионах в пределах одного рецептивного поля.
Утверждается, что весь вентральный визуально-гиппокамповый путь важен для визуальной памяти. Эта теория, в отличие от доминирующей, предсказывает, что изменения памяти распознавания объектов (англ. object-recognition memory, ORM) могут возникнуть в результате манипуляции в V2, области, которая очень взаимосвязана с вентральным путем зрительной коры. В мозге обезьяны эта область получает устойчивые упреждающие связи из первичной зрительной коры (V1) и посылает устойчивые проекции на другие вторичные зрительные отделы (V3, V4 и V5). Большинство нейронов этой области настроены на простые визуальные характеристики, такие как ориентация, пространственная частота, размер, цвет и форма. Анатомические исследования подразумевают слой 3 области V2 в обработке визуальной информации. В отличие от слоя 3, слой 6 зрительной коры состоит из многих типов нейронов, и их реакция на зрительные стимулы более сложна.
В недавнем исследовании было обнаружено, что клетки слоя 6 коры V2 играют очень важную роль в хранении памяти распознавания объектов, а также преобразовании краткосрочных объектных воспоминаний в долговременные воспоминания.
|
|
|
|---|---|
| Словари и энциклопедии |
|
|
Лобная
доля |
|
||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Теменная
доля |
|
||||||||||||||
|
Затылочная
доля |
|
||||||||||||||
|
Височная
доля |
|
||||||||||||||
|
Междолевые
борозды |
|
||||||||||||||
|
|||||||||||||||
|
Островковая
доля |
|
||||||||||||||
| Прочее | |||||||||||||||
| Зрительные доли | |
|---|---|

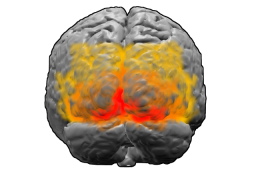
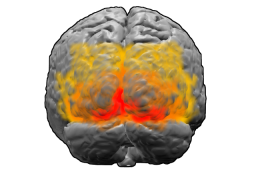
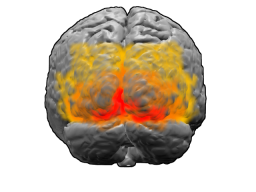
Мозг человека, вид сзади. Красным цветом обозначено поле Бродмана 17 (первичная зрительная кора); оранжевым — поле 18; жёлтым — поле 19 |
|

Мозг человека, вид слева. Вверху: латеральная поверхность, внизу: медиальная поверхность. Оранжевым цветом обозначено поле Бродмана 17 (первичная, или стриарная, зрительная кора) |
|
| Часть | Затылочная доля |
| Каталоги | |
|
|