
Физическая антропология
- 1 year ago
- 0
- 0

Физическая антропология и генетика русских — исследования комплекса наследственно определяемых признаков, характеризующих генотип и фенотип русских . Большинство антропологических и генетических показателей русских близко к средним значениям по Европе .
В 1654—1656 годах в России побывал путешественник архидиакон Павел Алеппский . Он оставил после себя подробные описания Царства Русского, быта местных жителей.
…в доме каждого человека по десяти и более детей с белыми волосами на голове; за большую белизну мы называли их старцами…
…Знай, что женщины в стране московитов красивы лицом и очень миловидны; их дети походят на детей франков, но более румяны…
Описания русских также оставили австрийский посол Сигизмунд Герберштейн , Адам Олеарий и другие.
Стоит выделить труды академика Императорской Академии наук и художеств Иоганна Готлиба Георги , который оставил первое подробное описание облика русского народа .

Антропологическое изучение русских началось вместе со становлением антропологии как самостоятельной науки в России . Первые работы, прежде всего по краниологии , в этой области принадлежат А. П. Богданову .
Начало обширных систематических исследований было положено трудом Д. Н. Анучина «О географическом распределении роста мужского населения России» (1889) . В своей работе он пользовался данными, полученными при осмотре лиц, призываемых на воинскую службу. Следующим шагом вперёд была монография Н. Ю. Зографа «Антропологические исследования мужского великорусского населения Владимирской, Ярославской и Костромской губерний» (1893) . В ней автор выделил два антропологических типа великорусского населения — высокорослый, светлый, мезокефальный и относительно более низкорослый, брахикефальный, темнопигметированный. Зограф выдвигал предположение, что второй тип сложился в результате смешения пришедших с запада славян с древним финноязычным населением Русской равнины .
Изучение антропологии восточных славян до середины XX века носило нерегулярный характер. Тем не менее, уже в это время был накоплен обширный научный материал, использованный и в более поздних работах и не потерявший актуальности до наших дней. Среди таких работ выделяются труды Д. Н. Анучина , Е. М. Чепурковского , В. В. Бунака , Г. Ф. Дебеца и Н. Н. Чебоксарова .
В работах Чепурковского впервые был последовательно применён разработанный им географический метод . В составе современного населения европейской части России и Украины автор выделил следующие варианты, имеющие чёткую географическую локализацию: западный великорус (валдайский тип), восточный великорус (северопонтийский или рязанский тип), широкоголовый шатен , северо-западный тип (ильменский тип) .
Наиболее масштабное антропологическое исследование русского народа было осуществлено в 1955 — 1959 годах Русской антропологической экспедицией под руководством В. В. Бунака . Целью ставилось выявление и описание основных антропологических элементов, вошедших в состав русских, и изучение путей их формирования. Исходя из этих целей, исследования начали проводить в зоне расселения предков русского народа в XI — XIV веках , а экспедиционные маршруты шли в соответствии с направлением колонизационных потоков восточнославянских племён средневековья ( вятичей , кривичей , северян и словен новгородских ). В течение пяти лет в 107 населённых пунктах по обширной антропологической программе было обследовано около 17 тысяч взрослых мужчин и женщин .
В 1953 — 1957 годах на северных территориях расселения русских работала экспедиция М. В. Витова . Маршрут разрабатывался в соответствии с колонизационными потоками из Новгородской и Ростово-Суздальской земель. В 80 территориальных группах Русского Севера было обследовано около 8 тысяч русских мужчин. Изучалось и расселённое на контактных территориях финно-угорское население .
В конце XX — начале XXI веков развитие молекулярной биологии и молекулярной генетики стимулировало геногеографические исследования народов мира, в том числе русских .
Продолжается обработка данных, собранных антропологическими экспедициями. Так, на основе этих данных был составлены портреты типичных представителей русского народа по географическим зонам . Антропологические данные анализируются совместно с геногеографическими, для получения более точной картины .
В ходе геногеографических экспедиций ГУ МГНЦ РАМН 2000 — 2008 годов было собрано и проанализировано более 10 000 образцов ДНК . Кроме того, исследовались частоты встречаемости русских фамилий , определена возможность использования этих данных совместно с геногеографическими .
В настоящее время геногеографические исследования продолжаются.
Русское население является довольно однородным в антропологическом отношении .
По трём антропологическим признакам (ширина головы, ширина носа, толщина губ) русские популяции достоверно не отличаются от западноевропейских. По остальным размерам головы и лица они близки к центральному европейскому варианту, характеризующемуся средними размерами. Средние величины группы или совпадают с центральными западноевропейскими величинами, или отклоняются от них, оставаясь, однако, в пределах колебаний западных групп.
Для русского населения характерно отсутствие эпикантуса . Из более чем 8,5 тысяч обследованных русских мужского пола эпикантус выявлен у 12 человек, к тому же только в зачаточном состоянии. Так же редок эпикантус у населения Центральной Европы .
Встречаемость прямого профиля носа у русских составляет 75 %, что выше среднего значения по Европе (70 %) . Встречаемость вогнутого профиля носа у русских составляет 9 %, что приближается к средним значениям по Западной и Центральной Европе (10 %) .
Сочетание признаков, характерных для русских популяций (в сравнении с западноевропейским центральным вариантом) :
Результаты краниологических исследований также демонстрируют исключительно высокое морфологическое сходство. Все локальные варианты проявляются в пределах основного единого и гомогенного краниологического типа русских .
Подобный невысокий уровень морфологической дифференциации в пределах весьма обширного ареала, занимаемого русскими, может иметь следующие причины :
Характер географической изменчивости антропологических признаков и их сочетания позволил выделить среди русского населения ряд локальных антропологических типов, получивших названия в соответствии с географической привязкой.
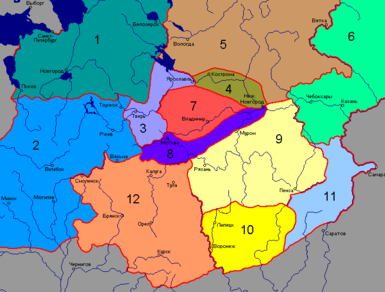




На основе данных, собранных Русской антропологической экспедицией , были выделены следующие антропологические типы:
На основе этой классификации были выделены и более крупные регионы: северо-запад, северо-восток, юго-запад, юго-восток, в пределах каждого из которых обнаруживается большое единство антропологических типов .
Морфогенез одонтологических признаков обусловлен главным образом генетическими факторами и слабо зависит от влияния внешней среды. Полиморфизм зубочелюстной системы широко используется в исследованиях проблем этногенеза .
Некоторые одонтологические характеристики в различных группах русских (%) :
| Группа |
Лопатообразность
резцов (баллы 2-3) |
Сумма
четырёхбугорковых М1 |
Сумма
шестибугорковых М1 |
Сумма
пятибугорковых М2 |
2med (II) |
|---|---|---|---|---|---|
| Кардымово ( Смоленская область ) | 7,4 | 6,3 | 3,5 | 14,6 | 40,0 |
| Починок ( Смоленская область ) | 9,4 | 18,6 | 3,0 | 13,7 | 50,5 |
| Смоленск | 8,9 | 12,9 | 5,5 | 10,8 | 42,6 |
| Старый Изборск ( Псковская область ) | 10,1 | 16,4 | 1,5 | 13,1 | 45,6 |
| Псков | 8,0 | 11,4 | 2,0 | 5,4 | 50,6 |
| Луга ( Ленинградская область ) | 2,4 | 8,8 | 3,3 | 12,6 | 65,4 |
| Мста ( Новгородская область ) | 4,7 | 18,6 | 1,4 | 5,6 | 54,1 |
| Валдай ( Новгородская область ) | 4,8 | 15,0 | 1,3 | 9,3 | 44,3 |
| Новгород | 5,6 | 16,9 | 1,1 | 8,2 | 58,1 |
| Западная Двина ( Калининская область ) | 8,5 | 6,5 | 2,2 | 8,1 | 57,8 |
| Калязин ( Калининская область ) | 4,4 | 11,6 | 3,5 | 6,6 | 46,3 |
| Вологодская область | 2,2 | 9,0 | 1,0 | 13,0 | 46,5 |
| Архангельская область | 9,3 | 20,5 | 2,0 | 15,3 | 34,1 |
| Рязанская область | 3,6 | 6,9 | 3,0 | 11,6 | 51,6 |
| Тамбовская область | 4,6 | 7,0 | 2,0 | 14,3 | 43,5 |
| Липецкая область | 5,2 | 13,6 | 2,3 | 8,3 | 45,3 |
| Воронежская область | 5,3 | 9,0 | 1,1 | 7,5 | 41,2 |
| Ростовская область | 7,4 | 6,9 | 4,6 | 18,3 | 40,0 |
| Тульская область | 3,3 | 10,0 | 2,2 | 10,5 | 56,5 |
| Орловская область | 1,7 | 8,4 | 1,1 | 17,6 | 47,4 |
| Курская область | 2,3 | 5,0 | 6,2 | 18,1 | 27,6 |
| Белгородская область | 1,7 | 3,0 | 5,0 | 7,1 | 42,4 |
| Калужская область | 8,3 | 10,1 | 3,8 | 6,6 | 37,7 |
| Брянская область | 10,7 | 16,9 | 2,4 | 16,4 | 30,0 |
Значения основных признаков одонтологии однозначно характеризуют русских как представителей западного круга форм . Одонтологические показатели русского населения наиболее близки к показателям среднеевропейского одонтологического типа .
Межгрупповые различия признаков у русских значительно меньше, чем между русскими и другими народами, что позволяет говорить о единстве русских . Тем не менее, на основании этих различий были выделены 2 основных одонтологических комплекса: северо-западный и южный, связанные между собой рядом переходных форм . Одонтологические типы в составе русского народа напрямую связаны с типами Восточной Европы в целом : среднеевропейским и северным грацильным . Выявлено, что северо-западный тип русских сформировался под влиянием северного грацильного типа Восточной Европы. В частности, для северо-западного одонтологического типа русских характерно некоторое повышение встречаемости лопатообразных резцов, что сближает эти популяции с соседними восточными латышами , эстонцами , финнами , карелами , вепсами и может объясняться влиянием общего субстрата . В целом встречаемость лопатообразных резцов, характерных для монголоидных популяций , у русских повсеместно низка .
Достоверных различий между городским и сельским населением областей, где такие исследования проводились, не выявлено .

Показано, что дерматоглифические признаки не подвержены влиянию направленного отбора и могут быть использованы в популяционных исследованиях наравне с генетическими .
Русские популяции являются достаточно однородными по признакам кожного рельефа . Тем не менее, в северных популяциях отмечаются черты северных европеоидов (пониженный дельтовый индекс сочетается с низким процентом добавочных межпальцевых трирадиусов), на юге — южноевропеоидные (относительно высокий процент добавочных межпальцевых трирадиусов сочетается с пониженной частотой осевого проксимального трирадиуса ладони у мужчин и относительно высоким дельтовым индексом у женщин) .
Европеоидно-монголоидный комплекс у русских, как правило, невысокий и в среднем составляет 41,9 у мужчин и 44,2 у женщин .
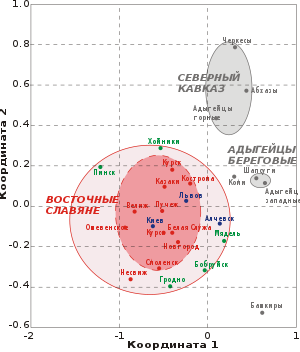
Восточные славяне обнаруживают высокое сходство друг с другом по дерматоглифическим данным. Ключевые признаки кожного рельефа у восточных славян варьируют незначительно, коэффициенты их вариации меньше, чем у европеоидов Европы и на территории Восточной Европы в целом . Дерматоглифический тип восточных славян является характерным для европеоидов Восточной Европы .
Минимальные обобщённые дерматоглифические расстояния отделяют восточных славян от мордвы , немцев , коми , молдаван , шведов . Обобщённые дерматоглифические расстояния между восточными славянами и литовцами , латышами , эстонцами , финнами , казанскими татарами , гагаузами выше, однако также находятся в пределах малых расстояний .)

Различное географическое распределение линий мтДНК и Y-хромосомы в русских популяциях генетики объясняют различным участием мужчин и женщин в освоении восточноевропейских территорий. Начальный этап колонизации осуществлялся как мужчинами, так и женщинами, в результате продвижения славянских племен на восток и север. В более поздний период ассимиляция местного населения проходила в основном через мужчин, которые женились на аборигенках .
По данным российских генетиков Елены Балановской и Олега Балановского , генофонд русского народа на территории Русской равнины — типично европейский . Процент восточноевразийских генетических маркеров в составе этой выборки не превышает средние показатели по Европе . Результаты изучения этой части русского генофонда показали его близость с населением практически всей Европы, при этом выявлены значительные отличия от населения Урала и Кавказа . Для генофонда выявлена широтная изменчивость (изменение характеристик по оси север — юг ). Этим русский генофонд отличается от общего генофонда Восточной Европы , для которого характерна долготная изменчивость. Таким образом, характер изменчивости русского генофонда не сводим к исходной изменчивости субстратов, на основе которых он формировался. Предполагается, что широтная изменчивость русского генофонда возникла или усилилась в ходе собственной истории русского народа .
Степень гетерогенности русского генофонда Восточно-Европейской равнины выше, чем у большинства остальных народов Европы. Это может быть связано с большой численностью русских и большой территорией расселения. Население Восточной Европы в целом более гетерогенно, чем население Западной Европы . При этом население Западной Сибири , наоборот, более гетерогенно, чем русские и иные народы Восточной Европы . Таким образом генофонды Восточной Европы, в том числе и русский генофонд по уровню гетерогенности занимают промежуточное положение между Западной Европой и генофондами народов Западной Сибири . В пределах самого русского генофонда существуют достаточно значительные отличия между северными популяциями русских, и существенно меньшие между южными и центральными, то есть население Русского Севера более гетерогенно .
Определённый интерес представляет генофонд северных русских . Северные популяции русских демонстрируют довольно сильные отличия от остальных русских популяций . До недавнего времени предполагалось, что такие отличия могут объясняться значительным финно-угорским компонентом, вошедшим в состав северных русских в ходе славянской колонизации этой территории . Последние исследования, включившие в себя комплексный анализ маркеров Y-хромосомы, мтДНК и аутосомных маркеров, позволяют сделать вывод о более выраженном сходстве северных русских с населением Северной Европы в целом . В частности, северные русские по комплексу признаков геногеографии наиболее схожи с балтами (латыши и литовцы) и шведами , достаточно малые генетические расстояния отделяют их от прибалтийско-финских ( финны , эстонцы , карелы , вепсы и ижорцы ) и некоторых пермско-финскими ( коми ) народами, а также от многих народов Центральной и Восточной Европы . При этом народы поволжско-финской группы удалены и от всего европейского массива, и от северных русских, что может свидетельствовать об отсутствии предполагавшегося ранее сильного вклада финно-угорского пласта в формирование населения Русского Севера . Таким образом, выявленное сходство генофонда северных русских с генофондами Центральной Европы и севера Восточной Европы позволяет выдвинуть гипотезу о сохранении на этих территориях древнего палеоевропейского генофонда, вошедшего в состав как северных русских, так и остальных народов Северной Европы .
По результатам исследования встречаемости гаплогрупп Y-хромосомы наиболее распространенные вариации гаплогрупп среди русских на 99,7 % совпадают с таковыми из Западной Евразии в рамках по крайней мере последних 3000 лет. Генетический пул русских сходен с остальными популяциями центральной и восточной Европы . В пределах русского генетического пула чётко выделяются две группы популяций. В северной группе (« Мезень », « Пинега », «Красноборск» ( Красноборский и Ленский районы)) обнаруживается близость с соседними финно-угорскими и балтскими популяциями. Южно-центральная группа, к которой относится подавляющее большинство русских популяций, входит в общий кластер с белорусами , украинцами и поляками .
Распределение гаплогрупп Y-хромосомы в некоторых исследованных русских популяциях :
| Популяция | N= | C | E1b1b1a1 | E1b1b1b2a1 | F * | G1 * | G2 | I * | I1 | I2a1 | I2a2a | J1 | J2 | K * | T | N * | N1a2b | N1a1 | Q | R1a | R1b1a1 | R1b1a2 | R2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M130 | M78 | M123 | M89 | M285 | P15 | M170 | M253 | P37 | M223 | M267 | M172 | M9 | M70 | M231 | P43 | TAT | M242 | SRY1532 | M73 | M269 | M124 | ||
| « Мезень » | 54 | 0 | 0 | 0 | 1,9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7,4 | 46,3 | 0 | 44,4 | 0 | 0 | 0 |
| « Пинега » | 114 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,9 | 4,4 | 0 | 0,9 | 0 | 0 | 0 | 0 | 15,8 | 23,7 | 0,9 | 39,5 | 0 | 14 | 0 |
| « Красноборск » | 91 | 0 | 0 | 0 | 1,1 | 0 | 2,2 | 3,3 | 12,1 | 9,9 | 0 | 0 | 5,5 | 0 | 0 | 0 | 3,3 | 36,3 | 0 | 19,8 | 0 | 6,6 | 0 |
| « Вологда » | 121 | 0 | 0,8 | 0 | 0 | 1,7 | 0,8 | 0 | 11,6 | 8,3 | 1,7 | 0 | 0,8 | 0 | 0 | 0 | 3,3 | 35,5 | 1,7 | 33,1 | 0 | 0,8 | 0 |
| Унжа | 52 | 0 | 5,8 | 0 | 1,9 | 0 | 0 | 0 | 11,5 | 11,5 | 3,8 | 3,8 | 3,8 | 0 | 0 | 0 | 0 | 13,5 | 0 | 32,7 | 1,9 | 9,6 | 0 |
| Кашин | 73 | 0 | 4,1 | 0 | 1,4 | 0 | 0 | 0 | 2,7 | 8,2 | 4,1 | 0 | 4,1 | 0 | 0 | 0 | 0 | 11 | 1,4 | 56,2 | 0 | 6,8 | 0 |
| Порхов | 57 | 0 | 1,8 | 1,8 | 0 | 0 | 0 | 1,8 | 3,5 | 10,5 | 0 | 0 | 0 | 3,5 | 0 | 1,8 | 1,8 | 15,8 | 0 | 52,6 | 0 | 5,3 | 0 |
| Остров | 75 | 0 | 4 | 0 | 0 | 0 | 0 | 1,3 | 6,7 | 9,3 | 0 | 1,3 | 1,3 | 0 | 0 | 0 | 0 | 28 | 0 | 45,3 | 0 | 2,7 | 0 |
| Рославль | 107 | 0,9 | 7,5 | 0 | 0,9 | 0 | 0 | 0 | 1,9 | 10,3 | 0 | 0 | 2,8 | 1,9 | 2,8 | 0 | 0,9 | 13,1 | 0 | 45,8 | 0 | 11,2 | 0 |
| Ливны | 110 | 0 | 0,9 | 0 | 0 | 0 | 0 | 0 | 8,2 | 13,6 | 0,9 | 0 | 0,9 | 0 | 2,7 | 0 | 0,9 | 4,5 | 0,9 | 62,7 | 0 | 3,6 | 0 |
| Пристень | 45 | 0 | 2,2 | 0 | 0 | 0 | 0 | 0 | 4,4 | 17,8 | 0 | 0 | 2,2 | 0 | 2,2 | 0 | 0 | 13,3 | 0 | 55,6 | 0 | 2,2 | 0 |
| Репьёвка | 96 | 0 | 1 | 0 | 1 | 0 | 2,1 | 1 | 3,1 | 16,7 | 0 | 2,1 | 1 | 0 | 0 | 0 | 0 | 6,3 | 0 | 59,4 | 0 | 5,2 | 1 |
| Белгород | 143 | 0,7 | 0,7 | 0 | 0 | 0 | 0,7 | 0 | 3,5 | 12,6 | 0,7 | 0 | 4,2 | 0,7 | 1,4 | 0 | 0,7 | 11,9 | 0 | 59,4 | 0 | 2,8 | 0 |
| Кубанские казаки | 90 | 0 | 3,3 | 0 | 0 | 0 | 1,1 | 1,1 | 4,4 | 16,5 | 2,2 | 0 | 4,4 | 0 | 1,1 | 0 | 1,1 | 6,6 | 1,1 | 47,3 | 0 | 8,8 | 0 |
По результатам многомерного анализа частот гаплогрупп митохондриальной ДНК русские входят в общеевропейский кластер популяций, включающий в себя все европейские популяции, за исключением саамов , удмуртов , башкир , ненцев , манси и народов Кавказа . Внутри данного кластера можно выделить более узкую группу популяций, в которую входят все европейские народы, в том числе русские, за исключением татар , чувашей и большинства финно-угорских популяций . Гомогенность популяций Европы в пределах данной «узко-европейской» группы высока . Тем не менее, в ней могут быть выделены отдельные кластеры, как правило, совпадающие с этно-лингвистическими группами . Русские входят в «славянский кластер», включающий в себя все исследованные народы славянской группы, а также венгров и эстонцев .
При сравнении митохондриального генофонда отдельных русских популяций с популяциями соседних народов было выделено 3 генетических кластера . В «восточноевропейский» кластер вошли южные и центральные русские, белорусы , украинцы , западнославянские и балтские народы . В «североевропейский» кластер вошли северные русские и западно-финские народы ( финны , карелы , мордва ) . Тюркские и восточно-финские народы Восточной Европы (татары, чуваши, коми , марийцы) образовали отдельный «приуральский» кластер .
При изучении внутрипопуляционной изменчивости митохондриальной ДНК в европейских популяциях была выявлена общая тенденция убывания разнообразия митохондриальной ДНК с юга на север, в которую укладываются и русские популяции . Этот факт может объясняться такими взаимосвязанными факторами, как меньшая плотность населения на севере, большая изолированность северных популяций, и как следствие, более высокая интенсивность дрейфа генов в них .
Митохондриальный геном русских в целом типично европейский , доля восточноевразийского компонента в русском митохондриальном геноме не превышает 2 % . Существуют определённые различия в доле восточноевразийского компонента в генофонде различных русских популяций. Среди русских южных и центральных регионов доля восточноевразийских гаплогрупп митохондриальной ДНК составляет 1,3 %, что сравнимо с показателями западных славян (1,6 %), среди северных русских — до 4 %, среди поморов — до 5 % .
Распределение гаплогрупп митохондриальной ДНК в некоторых исследованных русских популяциях :
| Популяция | N= | H | V | Pre*V | HV* | R1 | J | T* | T1 | K | U1 | U2 | U3 | U4 | U5a | U6b | U7 | U8 | I | N1a | N1b | W | X | C | D | G2a | M1 | L1b | L3b |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Тула | 73 | 58,9 | 1,4 | 1,4 | 1,4 | 0 | 5,5 | 8,2 | 1,4 | 2,7 | 0 | 0 | 2,7 | 4,1 | 2,7 | 2,7 | 0 | 0 | 2,7 | 1,4 | 1,4 | 0 | 0 | 0 | 0 | 0 | 0 | 1,4 | 0 |
| Калуга | 71 | 38,0 | 2,8 | 0 | 2,8 | 0 | 4,2 | 11,3 | 2,8 | 4,2 | 1,4 | 2,8 | 1,4 | 8,5 | 8,5 | 2,8 | 0 | 0 | 1,4 | 0 | 0 | 0 | 2,8 | 2,8 | 0 | 0 | 0 | 0 | 1,4 |
| Владимир | 72 | 38,5 | 0 | 0 | 1,4 | 0 | 8,3 | 8,3 | 11,1 | 6,9 | 0 | 1,4 | 1,4 | 2,8 | 6,9 | 0 | 1,4 | 0 | 5,6 | 1,4 | 1,4 | 4,2 | 1,4 | 0 | 0 | 0 | 1,4 | 0 | 0 |
| Ярославль | 41 | 48,8 | 0 | 2,4 | 4,9 | 0 | 4,9 | 2,4 | 0 | 0 | 7,3 | 0 | 0 | 0 | 4,9 | 9,8 | 0 | 0 | 0 | 2,4 | 2,4 | 2,4 | 0 | 4,9 | 0 | 2,4 | 0 | 0 | 0 |
| Псков | 68 | 42,6 | 5,9 | 0 | 0 | 1,5 | 11,8 | 2,9 | 2,9 | 0 | 0 | 2,9 | 0 | 2,9 | 17,6 | 2,9 | 0 | 1,5 | 0 | 0 | 0 | 2,9 | 0 | 0 | 0 | 1,5 | 0 | 0 | 0 |
Аутосомные маркеры — маркеры неполовых хромосом . Как правило, эти гены кодируют синтез определённых белков , ферментов и т. п.
Генетико-биохимические маркеры, как правило, представляют собой гены , кодирующие синтез ферментов . Полиморфизм некоторых иммунологических маркеров крови также может быть отнесён к биохимическому полиморфизму, так как эти гены кодируют ферменты, катализирующие соединение соответствующих сахаров с группоспецифическими гликопротеидами или гликолипидами . Показано, что полиморфизм многих биохимических маркеров может быть использован в популяционных исследованиях.
В таблице приведены частоты некоторых биохимических маркеров эритроцитарной фракции крови , сыворотки крови , иммунологических маркеров крови.
| Биохимический маркер | Геногеография | Аллельные частоты у русских | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Группа крови (система AB0) | Частота гена А — 0,05 среди населения Центральной и Южной Америки , 0,25—0,3 среди населения Центральной Европы, 0,4—0,45 среди населения Центральной Австралии и Северной Европы. Частота гена В — 0,0—0,05 среди населения американского континента, 0,05—0,1 среди населения Западной и Центральной Европы, 0,1—0,2 среди населения Восточной Европы, 0,2—0,3 среди азиатского населения. Частота гена 0 — 0,5—0,55 среди населения некоторых азиатских регионов, 0,6—0,65 среди населения Центральной и Восточной Европы, 0,65—0,7 среди населения Южной и Западной Европы, 0,95—1,0 среди населения Центральной и Южной Америки . | 0 | A | B | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0,598 | 0,247 | 0,156 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Резус-фактор | В странах Тихоокеанского бассейна ( Америка , Австралия , Океания , Восточная и Юго-Восточная Азия) концентрация варианта D, как правило, выше 0,8, в Южной и Юго-Западной Азии, а также в Африке (южнее Сахары ) до 0,8, в Европе и Северной Африке она ещё ниже, падая у берегов Атлантики ( Испания , Франция , Англия , Скандинавия ) ниже 0,6 . | Rh*D | Rh*d | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0,632 | 0,368 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Кислая эритроцитарная фосфатаза | Обнаружено шесть основных фенотипов , определяемых аллелями АСР1*А, АСР1*В, АСР1*С . Из них аллель АСР1*С является самым редким и встречается у европеоидов с частотой 0,03—0,07, у монголоидов, негроидов и аборигенов Австралии с частотой ещё меньшей, а в некоторых популяциях отсутствует полностью . | ACP1*A | ACP1*B | ACP1*C | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0,339 | 0,608 | 0,053 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Два аллеля GLO1*1 и GLO1*2, по крайней мере, среди европейского населения распределяются почти с одинаковой частотой . | GLO1*1 | GLO1*2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0,327 | 0,673 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Трансферрин | Два наиболее распространённых аллеля TF*C1 и TF*C2 присутствуют во всех популяциях. У европейцев и американских белых TF*C1 встречается с частотой 75—78 % . TF*C2 в европейских популяциях имеет частоту 13—19 %, в азиатских — 15—34 % . TF*C3 встречается европеоидных группах с частотой 4—7 %, в популяциях Индии — 1—4 %, отдельные случаи встречаемости в Восточной Азии, некоторых группах Тихоокеанского бассейна и у американских негров. В других популяциях он полностью отсутствует . По мнению ряда авторов, аллель С3 имеет европейское происхождение . TF*DChi является маркером монголоидных популяций и он широко распространён в Восточной Азии . | TF*B | TF*C1 | TF*C2 | TF*C3 | TF*DChi | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0,018 | 0,827 | 0,112 | 0,037 | 0,005 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Группоспецифический компонент | В европейских популяциях соотношение частот встречаемости аллелей GC*1F:GC*1S составляет от 2:10 до 3:10. Среди монголоидных популяций, в отличие от европейских, частота встречаемости GC*1F преобладает, ещё большая плотность этого гена свойственна негроидным популяциям . Частота встречаемости аллеля GC*2 составляет среди европеоидных популяций около 28 %, монголоидных — около 23 %, негроидных — около 8 % и среди аборигенов Австралии — около 12 % . | GC*1F | GC*1S | GC*2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0,119 | 0,615 | 0,262 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Фосфоглюкомутаза | Обнаружена направленная географическая изменчивость частот фосфоглюкомутазы в зависимости от климатических характеристик . | PGM1*1+ | PGM1*1- | PGM1*2+ | PGM1*2- | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0,591 | 0,241 | 0,090 | 0,079 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Эстераза D | Самыми высокими значениями аллеля EstD*1 характеризуются негроидные группы (0,89—0,95). Несколько ниже концентрация гена EstD*1 у европейцев (0,85—0,89). Таким образом, по уменьшению частоты гена EstD*1 популяции мира располагаются в следующем порядке: негроиды, европеоиды, группы Индии, монголоиды. Наиболее высокие частоты гена EstD*2 присущи монголоидам (до 0,408 у тибетцев). Остальные аллели встречаются чрезвычайно редко . | EstD*1 | EstD*2 | EstD*5 | EstD*7 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0,885 | 0,112 | 0,002 | 0,001 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Наиболее изученными в российских популяциях гипервариабельными минисателлитными локусами являются D1S80, D1S111, IL-1RN и 3’ApoB, так как они представляют наибольший интерес с точки зрения судебно-генетической экспертизы . Изучение полиморфизма локусов D1S80 и 3’ApoB также используется в популяционных исследованиях, так как данные маркеры обладают весьма высокой дифференцирующей способностью . Минисателлитный локус D1S80 локализован на коротком плече 1-й хромосомы . Минисателлитный локус 3’АроВ локализован на коротком плече 2-й хромосомы и представляет собой высокополиморфный участок на расстоянии 500 пар оснований после 3'-конца гена аполипопротеина В .
Исследования минисателлитных локусов D1S80 и 3’ApoB показали единство всех восточнославянских популяций и их отличие от коми-зырян , башкир , якутов и северокавказских народов, что может объясняться общим происхождением восточных славян и незначительностью влияния на их генофонд обозначенных этносов .
В 2011 году была исследована генетическая вариабельность 15 широко используемых в судебно-генетической экспертизе микросателлитных маркеров в 17 популяциях Восточной Европы, в том числе в 6 русских городских популяциях ( Москва , Белгород , Орёл , Ярославль , Оренбург , Томск ) .
Все русские группы, а также белорусы , украинцы и коми характеризуются общностью генного пула по указанным маркерам и отсутствием значимой межпопуляционной дифференциации . При этом группа славянских популяций (русские, белорусы, украинцы) значительно отличается от большинства других популяций, в частности, волго-уральского и северокавказского регионов . Все исследованные русские городские популяции, несмотря на значительные географические расстояния между ними, демонстрируют полное отсутствие межпопуляционных различий по исследованным 15 микросателлитным маркерам . По результатам многомерного шкалирования все европейские популяции, включая восточных славян и коми, вошли в общий кластер, что указывает на единство их генофонда . Народы волго-уральского региона и Северного Кавказа образуют отдельные кластеры .
Комплекс генов HLA ( главного комплекса гистосовместимости ) локализован на коротком плече 6-й хромосомы и занимает 3500 тысяч пар оснований , содержит более 220 генов . Он выполняет функции генетического контроля иммунного ответа и поддержания иммунного гомеостаза . Гены локуса HLA DP, DQ и DR кодируют молекулы II класса, выявленные на поверхности антиген-представляющих клеток (хелперов), в частности, В-лимфоцитов , Т-хелперов и некоторых других .
Система HLA является одной из наиболее полиморфных генетических систем. Показано, что, несмотря на влияние направленного естественного отбора , распределение генов HLA II класса адекватно отражает геногеографию этнических групп, и особенности полиморфизма генов HLA могут быть использованы в популяционных исследованиях .
Наибольший полиморфизм проявляет ген DRB1, его изучение наиболее информативно при популяционных исследованиях .
Частоты аллелей системы DRB1 в некоторых восточнославянских популяциях :
| Популяция |
Северные
белорусы ( Витебская область ) |
Южные
белорусы ( Брестская область ) |
Центральные
украинцы ( Хмельницкая область ) |
Западные
украинцы ( Львовская область ) |
Русские
( Вологодская область ) |
|---|---|---|---|---|---|
| Аллель DRB1 | N=100 | N=105 | N=137 | N=102 | N=121 |
| *01 | 10,0 % | 10,5 % | 9,5 % | 12,2 % | 12,4 % |
| *03 | 7,0 % | 9,0 % | 5,5 % | 7,8 % | 7,4 % |
| *04 | 7,5 % | 12,9 % | 10,9 % | 11,3 % | 14,0 |
| *07 | 10,5 | 10,5 | 14,6 | 15,2 | 14,9 |
| *08 | 5,5 % | 2,9 % | 3,6 % | 5,4 % | 5,4 % |
| *09 | 0 | 1,9 % | 0,4 % | 1,5 % | 2,5 % |
| *10 | 0 | 0,5 % | 1,1 % | 0 | 0,4 % |
| *11 | 15,0 % | 13,3 % | 18,6 % | 18,6 % | 9,5 % |
| *12 | 4,0 % | 3,8 % | 2,2 % | 0,5 % | 1,2 % |
| *13 | 16,5 % | 11,9 % | 12,4 % | 10,8 % | 14,9 % |
| *14 | 1,0 % | 2,9 % | 2,2 % | 2,0 % | 1,2 % |
| *15 | 18,0 % | 13,3 % | 10,2 % | 9,8 % | 14,5 % |
| *16 | 5,0 % | 6,7 % | 8,8 % | 5,9 % | 1,6 % |
Частоты аллелей гена DRB1 в популяциях восточных славян соответствуют типичному западноевразийскому варианту распределения . Встречаемость частот аллелей DRB1*09 и DRB1*12, характерных для монголоидных популяций , у русских низка .
По результатам многомерного анализа полиморфизма системы HLA в популяциях Европы выделяется три кластера: североевропейский, центральноевропейский и южноевропейский. Русские входят в североевропейский кластер, наряду с немцами , датчанами , голландцами , финнами , ирландцами и англичанами .
| Фольклор | |
|---|---|
| Культура | |
| Быт и обряды | |
| Религия | |
| Самосознание | |
| Политика | |
| Данные | |
| ФИО | |






















