Interested Article - РНК-термометр
- 2021-09-18
- 1
РНК-термо́метр , или РНК-термосе́нсор ( англ. RNA thermometer, RNA thermosensor, RNAT ) — температурочувствительная некодирующая РНК , которая принимает участие в регуляции экспрессии генов . РНК -термометры, как правило, регулируют гены , которые необходимы для ответа на или , однако показано их участие в регуляции длительного голодания и патогенности .
Принцип работы РНК-термометра заключается в изменении вторичной структуры этой молекулы в ответ на изменение температуры . В ходе этих структурных изменений важные участки этой РНК, например, , выставляются наружу или, наоборот, уходят вглубь молекулы, тем самым влияя на трансляцию близлежащего белоккодирующего гена.
РНК-термометры, наряду с рибопереключателями , служат доводами в поддержку гипотезы мира РНК . Согласно этой теории, сначала единственной нуклеиновой кислотой , представленной в клетках , была РНК, которая впоследствии была заменена современной системой ДНК → РНК → белок .
Примерами РНК-термометров могут служить , , , Hsp17-термометр .
История изучения
Об открытии первого температурочувствительного РНК-элемента было сообщено в 1989 году . Предшествующие исследования показали, что мутации , располагающиеся выше сайта начала трансляции в мРНК cIII фага лямбда (λ), оказывают влияние на уровень трансляции белка cIII . Этот белок участвует в выборе программы ( литический или лизогенный путь) жизненного цикла фага λ, причём высокая концентрация белка cIII соответствует лизогенному пути . Дальнейшие исследования показали, что у этого вышестоящего участка РНК имеются две альтернативные вторичные структуры. Оказалось, что эти структры не взаимозаменяемы и зависят от концентрации ионов Mg 2+ и температуры . Сейчас считается, что эти РНК-термометры запускают литический путь в условиях теплового шока, чтобы бактериофаг смог быстро реплицироваться и покинуть клетку-хозяина .
Термин «РНК-термометр» не использовался до 1999 года , когда так был назван РНК-элемент rpoH бактерии Escherichia coli . В недавнее время с помощью методов биоинформатики было выявлено несколько новых возможных РНК-термометров . В этом случае обычный поиск по последовательностям неэффективен, так как вторичная структура РНК-термометров гораздо более консервативна, чем их нуклеотидные последовательности .
Для изучения работы РНК-термометров применяют различные подходы. Для изучения динамики РНК-термометров можно заменять в них обычные нуклеотиды в определённых сайтах на флуоресцентные и, таким образом, наблюдать за их изменениями . Для определения положения РНК-термометра в исследуемой последовательности при определённых температурах был разработан специальный web-сервер RNAthermsw . Для идентификации бактериальных РНК-термометров используются и генетические методы, например, Tet-Trap .
Распространение
Большая часть известных сейчас РНК-термометров располагается в 5'-нетранслируемых областях (5'-UTR) прокариотических мРНК, кодирующих белки теплового шока . Возможно, такие результаты обусловлены и непреодолимыми сложностями в поиске коротких неконсервативных последовательностей в геномных данных .
Хотя большинство известных РНК-термометров обнаружены у прокариот (в том числе цианобактерий ), возможные РНК-термометры были выявлены у млекопитающих , в том числе и человека . У человека возможный термосенсор РНК теплового шока-1 (HSR1) активирует (HSF1) и запускает синтез защитных белков при температуре, превышающей 37 °C ( * ), и тем самым защищает клетки от перегревания . Цис -регуляторный элемент Hsp90 регулирует экспрессию шаперона hsp90 у дрозофилы , повышая его трансляцию при высоких температурах .
Структура
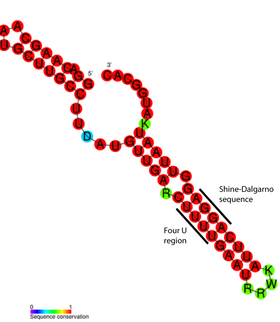
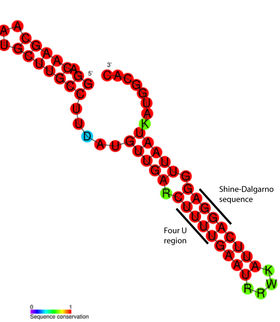
Структура РНК-термометров проста и может быть образована короткими последовательностями РНК. Длина наименьшего из известных РНК-термометров составляет 44 нуклеотида. Он располагается в мРНК белка теплового шока (hsp17) у цианобактерии sp. PCC 6803 . В общем случае длина РНК-термометров варьирует от 60 до 110 нуклеотидов , и они, как правило, содержат шпильку , в которой небольшая доля оснований не спарена . Они уменьшают стабильность структуры, благодаря чему она может легко расплавляться при повышении температуры .
Детальный структурный анализ РНК-термометра ROSE показал, что неспаренные основания на самом деле принимают участие в нестандартном спаривании оснований, которое поддерживает спиральную структуру РНК. Эти необычные пары представлены парами G -G, U -U и U C -U. Поскольку эти неканонические пары относительно нестабильны, повышение температуры вызывает локальное расплавление РНК в этой области, из-за чего последовательность Шайна — Дальгарно выставляется наружу .
Некоторые РНК-термометры имеют гораздо более сложную структуру, чем единственная шпилька, как в случае , где РНК-термометр содержит псевдоузел и множество шпилек .
Были разработаны искусственные РНК-термометры, содержащие одну лишь шпильку . Однако нуклеотидная последовательность столь коротких РНК-термометров может быть чувствительна к мутациям, и замена единственного основания может сделать этот РНК-термометр неактивным in vivo .
Механизм
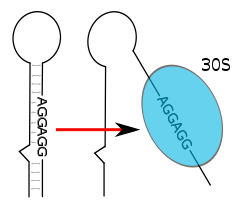
РНК-термометры располагаются в 5'-UTR мРНК, выше кодирующей последовательности . В отличие от рибопереключателей, действующих на уровне транскрипции , трансляции и регуляции стабильности мРНК, все известные на данный момент РНК-термометры действуют на уровне инициации трансляции . Структурные изменения в РНК-термометров могут убирать сайт связывания рибосомы в глубь молекулы и тем самым предотвращать трансляцию мРНК в белок . При повышении температуры шпилечная структура РНК-термометра может плавиться, выставляя наружу сайт связывания рибосомы или последовательность Шайна — Дальгарно (а в некоторых случаях старт-кодон AUG ), позволяя малой субъединице рибосомы ( ) связаться с мРНК, вслед за чем собирается и весь аппарат трансляции . Старт-кодон , располагающийся, как правило, на 8 нуклеотидов ниже последовательности Шайна — Дальгарно , отмечает начало белоккодирующей области , которую рибосома транслирует в пептид . Помимо таких цис -действующих РНК-термометров известен единственный транс -действующий РНК-термометр, располагающийся в мРНК , где он, как предполагается, регулирует ответ на длительное голодание .
В качестве примера можно рассмотреть РНК-термометр FourU Salmonella enterica . Под действием температур выше 45 °C шпилька, содержащая последовательность Шайна — Дальгарно , плавится, последовательность Шайна—Дальгарно становится неспаренной и трансляция мРНК становится возможной . Показано, что на стабильность FourU влияет концентрация Mg 2+ . Наиболее изучен РНК-термометр, расположенный в мРНК гена rpoH у E. coli . Этот термосенсор положительно регулирует трансляцию белков теплового шока при высоких температурах посредством специализированного сигма-фактора σ 32 .
У Bradyrhizobium japonicum и Rhizobium radiobacter , протеобактерий порядка Rhizobiales, описаны РНК-термометры ROSE 1 и ROSE AT2 соответственно. Они располагаются в 5'-UTR HspA и подавляют трансляцию белков теплового шока при физиологических температурах .
Хотя РНК-термометры обычно связаны с экспрессией белков теплового шока, они могут также регулировать экспрессию белков холодового шока . Например, у термофильной бактерии Thermus thermophilus экспрессия двух белков массой 7 кДа регулируется РНК-термометром , кроме того, похожий механизм был описан у Escherichia coli .
РНК-термометры, реагирующие на температуру 37 °C, могут использоваться патогенными микроорганизмами для активации генов, связанных с инфекцией . Например, путём пришивания гена, кодирующего зелёный флуоресцентный белок , к 5'-концу гена , кодирующего ключевой регулятор транскрипции генов вирулентности у , была продемонстрирована положительная регуляция экспрессии prfA : при транскрипции такого гибридного гена с промотора Т7 E. coli флуоресценция наблюдалась при 37 °C, но не при 30 °C . РНК-термометры вовлечены в регуляцию вирулентности таких болезнетворных бактерий, как и Vibrio cholerae . У болезнетворной бактерии Shigella dysenteriae и патогенных штаммов Escherichia coli РНК-термометры вовлечены в регуляцию процессов, влияющих на патогенез .
Иногда оперон может регулироваться несколькими РНК-термометрами. Предсказано, что оперон ibpAB E. coli содержит два кооперативных РНК-термометра: элемент ROSE и .
Стоит также отметить, что РНК-термометры могут использоваться не только для регуляции трансляции моно цистронных транскриптов, содержащих одну последовательность Шайна — Дальгарно , но и для полицистронных транскриптов, содержащих несколько последовательностей Шайна — Дальгарно . Например, у устойчивость к стрессу обеспечивается трицистронным опероном, консервативным среди многих свободноживущих бактерий. Первые два гена этого оперона регулируются РНК-термометрами .
РНК-термометры и гипотеза мира РНК
Гипотеза мира РНК утверждает, что изначально РНК выступала носителем наследственной информации и осуществляла ферментативные процессы, причём различные последовательности РНК выступали био катализаторами , регуляторами и сенсорами . Позже под действием отбора большая часть функций, осуществляемых РНК, стала выполняться другими биомолекулами , и на смену жизни, основанной исключительно на РНК, пришла жизнь, основанная на ДНК , РНК и белке .
Считается, что РНК-термометры и рибопереключатели являются эволюционно древними элементами, поскольку они широко распространены у самых эволюционно далёких организмов . Было высказано предположение, что в мире РНК РНК-термометры осуществляли температурозависимую регуляцию других РНК . У современных организмов РНК-термометры, возможно, являются « молекулярными ископаемыми », которые в ушедшем мире РНК были гораздо более распространены, чем сейчас .
Применение
Для температурного контроля экспрессии генов у бактерий разрабатываются искусственные РНК-термометры .
В 2013 году были разработаны «термозимы» — искусственные РНК-термометры с рибозимной активностью. Термосенсорная шпилька в расплавленном состоянии подавляет работу рибозима, который высвобождает последовательность связывания рибосомы. При повышенных температурах шпилька плавится, рибозим инактивируется и экспрессия гена подавляется. Таким образом, термозим реагирует на повышенные температуры противоположно природным РНК-термометрам .
В 2016 году было сообщено о создании «термопереключателей» — интеграции температурочувствительных РНК-термометров и аптамеров рибопереключателей в единую структуру. Термопереключатели функционируют как рибопереключатели при низких температурах и реагируют на связывание со своим лигандом изменением структуры, а при высокой температуре они переходят в постоянно «включённое» состояние. Таким образом, термопереключатели — первые РНК-термометры, действующие на уровне транскрипции . Подобные искусственные РНК-регуляторы могут широко применяться для регуляции экспрессии генов .
В 2016 году был предложен алгоритм RNAiFold2T для разработки особых РНК-термометров, содержащих IRES. Кэп -независимая трансляция таких термо-IRES-элементов примерно на 50 % интенсивнее при 42 °С, чем при 30 °С. Впрочем, эффективность их трансляции всё равно меньше, чем у IRES дикого типа, которая не зависит от температуры .
Примечания
- ↑ Narberhaus F. , Waldminghaus T. , Chowdhury S. (англ.) // FEMS microbiology reviews. — 2006. — Vol. 30, no. 1 . — P. 3—16. — doi : . — .
- ↑ Atkins, John F.; Gesteland, Raymond F.; Cech, Thomas. The RNA world: the nature of modern RNA suggests a prebiotic RNA world (англ.) . — Plainview, N.Y: , 2006. — ISBN 0-87969-739-3 .
- ↑ Waldminghaus T. , Heidrich N. , Brantl S. , Narberhaus F. (англ.) // Molecular microbiology. — 2007. — Vol. 65, no. 2 . — P. 413—424. — doi : . — .
- ↑ Ahmed R. , Duncan R. F. (англ.) // The Journal of biological chemistry. — 2004. — Vol. 279, no. 48 . — P. 49919—49930. — doi : . — .
- ↑ Nocker A. , Hausherr T. , Balsiger S. , Krstulovic N. P. , Hennecke H. , Narberhaus F. (англ.) // Nucleic acids research. — 2001. — Vol. 29, no. 23 . — P. 4800—4807. — .
- ↑ Kortmann J. , Sczodrok S. , Rinnenthal J. , Schwalbe H. , Narberhaus F. (англ.) // Nucleic acids research. — 2011. — Vol. 39, no. 7 . — P. 2855—2868. — doi : . — .
- ↑ Altuvia S. , Kornitzer D. , Teff D. , Oppenheim A. B. (англ.) // Journal of molecular biology. — 1989. — Vol. 210, no. 2 . — P. 265—280. — .
- ↑ Altuvia S. , Oppenheim A. B. (англ.) // Journal of bacteriology. — 1986. — Vol. 167, no. 1 . — P. 415—419. — .
- Altuvia S. , Kornitzer D. , Kobi S. , Oppenheim A. B. (англ.) // Journal of molecular biology. — 1991. — Vol. 218, no. 4 . — P. 723—733. — .
- ↑ Storz G. (англ.) // Genes & development. — 1999. — Vol. 13, no. 6 . — P. 633—636. — .
- Morita M. T. , Tanaka Y. , Kodama T. S. , Kyogoku Y. , Yanagi H. , Yura T. (англ.) // Genes & development. — 1999. — Vol. 13, no. 6 . — P. 655—665. — .
- ↑ Waldminghaus T. , Gaubig L. C. , Narberhaus F. (англ.) // Molecular genetics and genomics : MGG. — 2007. — Vol. 278, no. 5 . — P. 555—564. — doi : . — .
- Narayan S. , Kombrabail M. H. , Das S. , Singh H. , Chary K. V. , Rao B. J. , Krishnamoorthy G. (англ.) // Nucleic acids research. — 2015. — Vol. 43, no. 1 . — P. 493—503. — doi : . — .
- Churkin A. , Avihoo A. , Shapira M. , Barash D. (англ.) // Public Library of Science ONE. — 2014. — Vol. 9, no. 4 . — P. e94340. — doi : . — .
- Delvillani F. , Sciandrone B. , Peano C. , Petiti L. , Berens C. , Georgi C. , Ferrara S. , Bertoni G. , Pasini M. E. , Dehò G. , Briani F. (англ.) // RNA (New York, N.Y.). — 2014. — Vol. 20, no. 12 . — P. 1963—1976. — doi : . — .
- ↑ Narberhaus F. (англ.) // RNA biology. — 2010. — Vol. 7, no. 1 . — P. 84—89. — .
- Johansson J. (англ.) // Contributions to microbiology. — 2009. — Vol. 16. — P. 150—160. — doi : . — .
- ↑ Krajewski S. S. , Narberhaus F. (англ.) // Biochimica et biophysica acta. — 2014. — Vol. 1839, no. 10 . — P. 978—988. — doi : . — .
- ↑ Shamovsky I. , Ivannikov M. , Kandel E. S. , Gershon D. , Nudler E. (англ.) // Nature. — 2006. — Vol. 440, no. 7083 . — P. 556—560. — doi : . — .
- ↑ Chowdhury S. , Maris C. , Allain F. H. , Narberhaus F. (англ.) // The EMBO journal. — 2006. — Vol. 25, no. 11 . — P. 2487—2497. — doi : . — .
- Waldminghaus T. , Fippinger A. , Alfsmann J. , Narberhaus F. (англ.) // Biological chemistry. — 2005. — Vol. 386, no. 12 . — P. 1279—1286. — doi : . — .
- ↑ Breaker R. R. (англ.) // Molecular cell. — 2010. — Vol. 37, no. 1 . — P. 1—2. — doi : . — .
- ↑ Giuliodori A. M. , Di Pietro F. , Marzi S. , Masquida B. , Wagner R. , Romby P. , Gualerzi C. O. , Pon C. L. (англ.) // Molecular cell. — 2010. — Vol. 37, no. 1 . — P. 21—33. — doi : . — .
- ↑ Neupert J. , Karcher D. , Bock R. (англ.) // Nucleic acids research. — 2008. — Vol. 36, no. 19 . — P. e124. — doi : . — .
- ↑ Nikolova E. N. , Al-Hashimi H. M. (англ.) // RNA (New York, N.Y.). — 2010. — Vol. 16, no. 9 . — P. 1687—1691. — doi : . — .
- ↑ Roßmanith J. , Narberhaus F. (англ.) // Nucleic acids research. — 2016. — doi : . — .
- Rinnenthal J. , Klinkert B. , Narberhaus F. , Schwalbe H. (англ.) // Nucleic acids research. — 2011. — Vol. 39, no. 18 . — P. 8258—8270. — doi : . — .
- Shah P. , Gilchrist M. A. (англ.) // Public Library of Science ONE. — 2010. — Vol. 5, no. 7 . — P. e11308. — doi : . — .
- Balsiger S. , Ragaz C. , Baron C. , Narberhaus F. (англ.) // Journal of bacteriology. — 2004. — Vol. 186, no. 20 . — P. 6824—6829. — doi : . — .
- Mega R. , Manzoku M. , Shinkai A. , Nakagawa N. , Kuramitsu S. , Masui R. (англ.) // Biochemical and biophysical research communications. — 2010. — Vol. 399, no. 3 . — P. 336—340. — doi : . — .
- Johansson J. , Mandin P. , Renzoni A. , Chiaruttini C. , Springer M. , Cossart P. (англ.) // Cell. — 2002. — Vol. 110, no. 5 . — P. 551—561. — .
- Weber G. G. , Kortmann J. , Narberhaus F. , Klose K. E. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2014. — Vol. 111, no. 39 . — P. 14241—14246. — doi : . — .
- Kouse A. B. , Righetti F. , Kortmann J. , Narberhaus F. , Murphy E. R. (англ.) // Public Library of Science ONE. — 2013. — Vol. 8, no. 5 . — P. e63781. — doi : . — .
- Viswanathan V. K. (англ.) // Gut microbes. — 2013. — Vol. 4, no. 4 . — P. 267—268. — doi : . — .
- Gaubig L. C. , Waldminghaus T. , Narberhaus F. (англ.) // Microbiology (Reading, England). — 2011. — Vol. 157, no. Pt 1 . — P. 66—76. — doi : . — .
- Krajewski S. S. , Joswig M. , Nagel M. , Narberhaus F. (англ.) // Environmental microbiology. — 2014. — Vol. 16, no. 6 . — P. 1835—1853. — doi : . — .
- Walter Gilbert. The RNA World (англ.) // Nature. — 1986. — February ( vol. 319 , no. 6055 ). — P. 618—618 . — doi : . — .
- Serganov A. , Patel D. J. (англ.) // Nature reviews. Genetics. — 2007. — Vol. 8, no. 10 . — P. 776—790. — doi : . — .
- Bocobza S. E. , Aharoni A. (англ.) // Trends in plant science. — 2008. — Vol. 13, no. 10 . — P. 526—533. — doi : . — .
- Neupert J. , Bock R. (англ.) // Nature protocols. — 2009. — Vol. 4, no. 9 . — P. 1262—1273. — doi : . — .
- Saragliadis A. , Krajewski S. S. , Rehm C. , Narberhaus F. , Hartig J. S. (англ.) // RNA biology. — 2013. — Vol. 10, no. 6 . — P. 1010—1016. — doi : . — .
- Garcia-Martin J. A. , Dotu I. , Fernandez-Chamorro J. , Lozano G. , Ramajo J. , Martinez-Salas E. , Clote P. (англ.) // Bioinformatics. — 2016. — Vol. 32, no. 12 . — P. 360—368. — doi : . — .
Литература
- O. Ю. Лиманская, Л. А. Муртазаева, А. П. Лиманский. // Микробиология. — 2013. — Т. 82 , № 1 . — С. 69—78 .
- Righetti F. , Narberhaus F. (англ.) // Frontiers in cellular and infection microbiology. — 2014. — Vol. 4. — P. 132. — doi : . — .
- Krajewski S. S. , Narberhaus F. (англ.) // Biochimica et biophysica acta. — 2014. — Vol. 1839, no. 10 . — P. 978—988. — doi : . — .
|
Виды
РНК
|
|
|---|---|
| Биосинтез белка | |
| Процессинг РНК | |
| Регуляция экспрессии генов | |
| Цис-регуляторные элементы | |
| Паразитические элементы | |
| Прочее | |
- 2021-09-18
- 1
