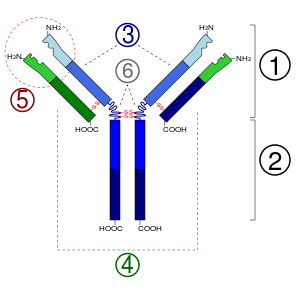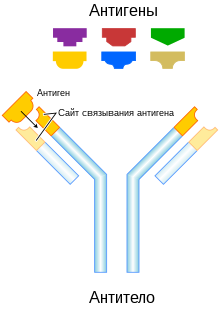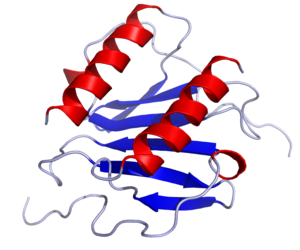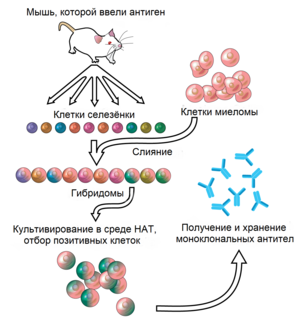
Антитела
- 1 year ago
- 0
- 0
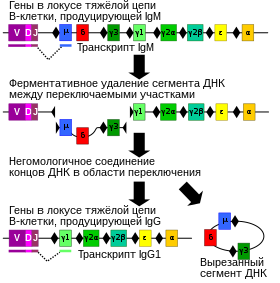
Сомати́ческая гипермута́ция ( англ. Somatic hypermutation ) — один из молекулярных механизмов, обеспечивающих разнообразие антител . За счёт соматической гипермутации создаётся разнообразие B-клеточных рецепторов и антител, благодаря чему увеличивается спектр антигенов , которые может распознать адаптивная иммунная система . В ходе соматической гипермутации в вариабельных участках генов иммуноглобулинов происходит множество точечных мутаций . В отличие от мутаций в клетках зародышевой линии , мутации, появившиеся в результате соматической гипермутации, не передаются потомкам и имеются только в геномах B-клеток . Если соматическая гипермутация затрагивает участки, отличные от тех, которые кодируют вариабельные домены иммуноглобулинов, развиваются и другие формы онкологических заболеваний .
После распознавания антигена B-клетка приступает к пролиферации . В ходе клеточных делений локус , кодирующий B-клеточный рецептор, характеризуется повышенной частотой точечных мутаций, которая в 10 5 —10 6 раз превышает частоту мутаций в других участках генома . Как правило, в ходе соматической гипермутации изменяются азотистые основания , вставки и делеции происходят реже. Мутации особенно часто затрагивают гипервариабельные участки в составе последовательности, кодирующей , в составе вариабельных доменов иммуноглобулинов, взаимодействующих с антигеном . Контекст, который наиболее благоприятен для внесения мутации, зависит от основания: G чаще всего мутирует в контексте RGYW, C — WRCY, A — WA, T — TW . Конечный итог соматической гипермутации зависит от действия систем репарации . Направленная гипермутация позволяет отбирать B-клетки, которые обладают повышенным сродством к данному антигену .

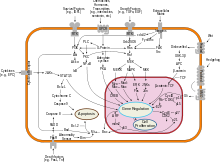
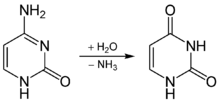
В основе соматических гипермутаций лежит реакция дезаминирования цитозина в составе ДНК до урацила , который имеется в РНК вместо тимина. Эту реакцию катализирует фермент , известный как . В результате действия фермента пара гуанин: цитозин заменяется на гуанин: урацил. Поскольку урацил в норме не встречается в ДНК, такие замены репарируются по пути эксцизионной репарацией оснований . Остатки урацила удаляет фермент репарации . Для заполнения образовавшейся бреши привлекаются ДНК-полимеразы , склонные к ошибкам, и в результате их работы возникают точечные мутации .
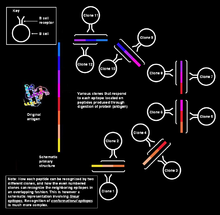
Появление точечных мутаций в быстро пролиферирующих B-клетках приводит к образованию тысяч новых B-клеток, несущих слегка отличающиеся последовательности, кодирующие вариабельные домены. Эти клетки имеют немного отличающиеся B-клеточные рецепторы с разной специфичностью к антигенам, и в ходе отбора остаются те B-клетки, рецепторы которых имеют наибольшее сродство к данному антигену. B-клетки, несущие рецепторы с наибольшим сродством к антигену, дифференцируются в плазматические клетки , секретирующие антитела, и , которые обеспечивают быстрый адаптивный иммунный ответ при повторном заражении тем же патогеном .
Помимо мутаций, связанных с превращением цитозина в урацил, свой вклад в увеличение разнообразия антител вносит редактирование РНК иммуноглобулинов, при котором аденозин переходит в инозин (I) .
Существуют свидетельства в пользу другого механизма соматической гипермутации, который подразумевает синтез кДНК с пре-мРНК иммуноглобулинов, синтезированной с ошибками, и интеграцию мутантной кДНК в соответствующий локус в хромосоме вместо немутантного фрагмента (то есть обратную транскрипцию ). Было высказано предположение, что синтез кДНК начинается после A → I редактирования пре-мРНК, и после интеграции мутантной кДНК в хромосому в изменённом куске в парах A-T соотношение аденозина и тимина в кодирующей цепи смещается в сторону тимина, что и характерно для локусов, подвергающихся соматической гипермутации и гипермутации при раке .
| Органы | |||||
|---|---|---|---|---|---|
|
Врождённый
иммунитет |
|
||||
|
Приобретённый
иммунитет |
|
||||
| Медиаторы | |||||
| Иммунный ответ | |||||
|
Иммунологическая
толерантность |
|||||
| Нарушения | |||||
|
Лимфоцитарная
адаптивная иммунная система
и
комплемент
|
|||||||||
|---|---|---|---|---|---|---|---|---|---|
| Лимфоидные |
|
||||||||
| Лимфоциты | |||||||||
| Вещества | |||||||||