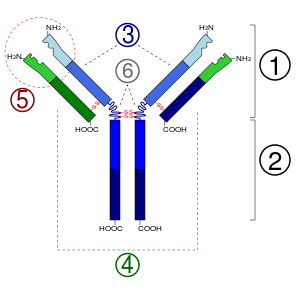
Антитела
- 1 year ago
- 0
- 0
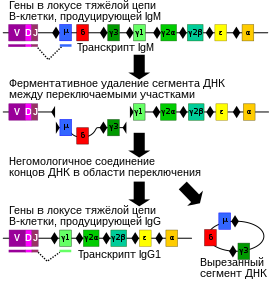

Переключе́ние кла́ссов антите́л , или переключе́ние кла́ссов иммуноглобули́нов , или переключе́ние изоти́пов ( англ. immunoglobulin class switching ), — процесс переключения B-лимфоцита (B-клетки) с синтеза антител одного класса на синтез антител другого класса, например, с иммуноглобулинов M ( IgM ) на иммуноглобулины G ( IgG ). В ходе переключения классов константная часть локуса IGH , кодирующего тяжёлую цепь антител, претерпевает изменения, в то время как вариабельный участок остаётся неизменным. Так как вариабельный участок антитела не меняется, его антигенная специфичность остаётся прежней, и антитело продолжает распознавать тот же эпитоп .
В кластере генов IGH присутствует несколько генов константных доменов антител, которые экспрессируются в определённой последовательности в комбинации с одним и тем же геном, кодирующим вариабельный домен. У человека гены в кластере IGH располагаются в последовательности: Cμ, Cδ, Cγ3, Cγ1, Cα1, Cγ2, Cγ4, Cε, Cα2, а у мыши — Сμ, Сδ, Сγ3, Сγ1, Сγ2b, Cγ2a, Cε, Cα. Между Cγ1 и Сα1 у человека располагается псевдоген , гомологичный Cε мыши .
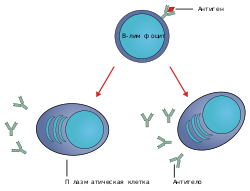
Переключение классов происходит в зрелой B-клетке после её активации при участии мембраносвязанной молекулы антитела или B-клеточного рецептора . В результате B-клетка начинает производить антитела другого класса, то есть с другой константной частью тяжёлой цепи, но с тем же вариабельным доменом, который был сформирован за счёт V(D)J-рекомбинации в незрелой B-клетке .
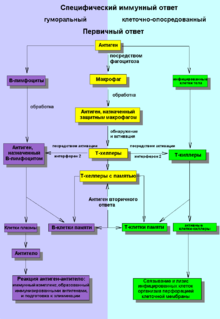
производят антитела классов IgM и IgD , которым соответствуют два первых сегмента тяжёлых цепей в локусе. После активации антигеном B-клетки приступают к пролиферации . Если их поверхностные молекулы CD40 и CD154 или свяжутся с определёнными сигнальными молекулами (при участии T-хелперов ), то в B-клетках происходит переключение классов антител, и они начинают вырабатывать антитела классов IgG, IgA или IgE . Благодаря переключению классов дочерние клетки , происходящие от одной B-клетки, секретируют антитела разных или подтипов (например, IgG1, IgG2 и другие) .
Процесс, ответственный за переключение классов антител, называется рекомбинационным переключением классов ( англ. class switch recombination, CSR ). В ходе CSR некоторые гены локуса тяжёлой цепи антитела удаляются, и образовавшиеся на месте разрывов концы соединяются, в результате чего образуется функциональный участок, кодирующий антитело другого изотипа. , образующиеся при CSR, происходят в районе консервативных нуклеотидных мотивов , называемых участками переключения (S-участками). S-участки имеют последовательности G A G C T или GGGGGT. Переключению предшествует их полимеризация (до 150 повторов). В начале процесса переключения происходит сближение полимерных S-участков и формируется петля, в которую попадают С-гены, расположенные между V-геном и C-геном, который в дальнейшем будет экспрессироваться. По краям петли вносятся два двуцепочечных разрыва по двум S-участкам при участии разнообразных ферментов , среди которых: ( англ. activation-induced (cytidine) deaminase, AID ), , ; они получают доступ к ДНК благодаря предшествующему ремоделированию хроматина . Фрагмент ДНК между двумя S-участками удаляется из хромосомы , в результате чего гены константных доменов тяжёлых цепей μ и δ вырезаются и замещаются соответствующими генами тяжёлых цепей γ-, ε- или α-типа. Сшивание свободных концов ДНК происходит в ходе негомологичного соединения концов , благодаря чему ген вариабельного домена соединяется с геном нужного типа константного домена тяжёлой цепи . Концы ДНК могут быть соединены и без негомологичного соединения концов — путём микрогомологичного соединения концов . За исключением IgD и IgM, в определённый момент времени B-клетка образует антитела только одного класса. Хотя переключение классов в большинстве случаев является результатом перестройки одной хромосомы, в 10—20 % случаев (в зависимости от классов антител) происходят межхромосомные транслокации между гомологичными хромосомами , при которых перемешиваются участки генов тяжёлых цепей с разных аллелей .
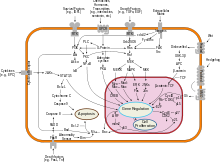
Ремоделирование хроматина , доступность S-участков для аппарата транскрипции и , а также сшивание ДНК после внесения разрывов в S-участки находятся под контролем супер энхансера , известного как 3'-регуляторный участок (3'-RR) . В некоторых случаях 3'-RR может сам становиться мишенью AID, и в него вносятся двуцепочечные разрывы, что в итоге приводит к делеции локуса тяжёлой цепи. Этот процесс известен как ( англ. locus suicide recombination, LSR ) .
Как правило, переключение — необратимый процесс, так как ненужные гены C-доменов удаляются в процессе переключения, однако из этого правила известны единичные исключения .
Важнейшую роль в регуляции переключения классов антител играют цитокины , которые секретируют T-хелперы и регуляторные T-клетки . В таблицах ниже представлены сведения о том, как различные цитокины влияют на переключение синтеза определённых классов антител у человека и у мыши, а также указаны T-клетки, которые выделяют эти цитокины .
| T-клетки | Цитокины | Классы антител | |||||
|---|---|---|---|---|---|---|---|
| IgG1 | IgG2a | IgG2b | IgG3 | IgG4 | IgE | ||
| T h 2 | IL-4 | ↑ | ↓ | ↓ | ↓ | ↓ | ↑ |
| IL-5 | ↑ | ||||||
| T h 1 | IFNγ | ↓ | ↑ | ↓ | ↑ | ↓ | ↓ |
| Treg | TGFβ | ↑ | ↓ | ↑ | |||
| IL-10 | ↑ | ||||||
| T-клетки | Цитокины | Классы антител | |||||
|---|---|---|---|---|---|---|---|
| IgG1 | IgG2 | IgG3 | IgG4 | IgA | IgE | ||
| T h 2 | IL-4 | ↑ | ↓ | ↑ | ↑ | ||
| IL-5 | ↑ | ||||||
| T h 1 | IFNγ | ↓ | ↑ | ↓ | |||
| Treg | TGFβ | ↓ | ↑ | ||||
| IL-10 | ↑ | ↑ | |||||
| Термины | |
|---|---|
| Классы антител | |
| Органы | |||||
|---|---|---|---|---|---|
|
Врождённый
иммунитет |
|
||||
|
Приобретённый
иммунитет |
|
||||
| Медиаторы | |||||
| Иммунный ответ | |||||
|
Иммунологическая
толерантность |
|||||
| Нарушения | |||||
|
Лимфоцитарная
адаптивная иммунная система
и
комплемент
|
|||||||||
|---|---|---|---|---|---|---|---|---|---|
| Лимфоидные |
|
||||||||
| Лимфоциты | |||||||||
| Вещества | |||||||||