Interested Article - Гаплогруппа H (мтДНК)
- 2020-08-29
- 1
Гаплогруппа H — гаплогруппа митохондриальной ДНК человека. Является потомком гаплогруппы HV . « Кембриджская эталонная последовательность » — человеческая митохондриальная последовательность, с которой сравниваются все прочие, принадлежит гаплогруппе H.
Происхождение
Ряд независимых исследований показал, что гаплогруппа H, предположительно, возникла в западной Азии около 30 тыс. лет назад, прибыла в Европу около 20—25 тысяч лет назад и быстро распространилась на юго-запад континента во франко-кантабрийский регион . В период последнего ледникового максимума (последнего оледенения) 20—13 тысяч лет назад большинство палеолитических поселений Северной и Центральной Европы вымерло, в связи с чем представители гаплогруппы Н в большей степени выжили лишь на севере Испании (поэтому в настоящее время данная гаплогруппа хоть и распространена по всей Европе, но с наибольшей частотой, более 50 %, встречается среди басков) . По мнению некоторых генетиков, на Пиренейском полуострове данная гаплогруппа встречалась у 70 % населения; оттуда носители гаплогрупы Н, будучи носителями культуры колоколовидных кубков , заселили оставшуюся часть Европы .
Предполагается, что распространение подклассов H1, H3, а также сестринской гаплогруппы V связано с внутриевропейской экспансией во франко-кантабрийском регионе после последнего ледникового максимума около 13 000 лет назад . Данные молекулярной генетики говорят о том, что франко-кантабрийский регион был колыбелью большей части населения Европы, по крайней мере, по женской линии (через гаплогруппу H) .
Палеогенетика
Древняя митохондриальная ДНК останков, известных как «Пальиччи 23» ( ) возрастом 28 тысяч лет из пещеры Пальиччи ( Апулия , Италия ) соответствует эталонной кембриджской последовательности HVR1, указывающей на то, что индивидуум имел либо митохондриальную гаплогруппу R , либо митохондриальную гаплогруппу H. Свидетельством достоверности данного открытия является то, что гаплотип данных останков отличался от гаплотипа всех лиц, работавших с данными останками со времени их обнаружения .
Субклада H13c была обнаружена у мезолитического охотника из карстового грота в известняках плато Мандаэти в Западной Грузии, жившего 9529—9895 лет назад .
Гаплогруппа H была обнаружена у представителя старчевской культуры , жившего ок. 7600 лет назад и у представителей днепро-донецкой культуры , живших ок. 7500 лет назад .
Субклады H3 и H4a1 были обнаружены у представителей неолитической культуры кардиальной керамики , живших 7400 лет назад .
Носителем митохондриальной гаплогруппы H1 был обитатель раннего неолита (7200 — 7000 л. н.) из Карсдорфа (Саксония-Анхальт, Германия) .
Гаплогруппа H была обнаружена у представителей культуры линейно-ленточной керамики , живших ок. 7000 лет назад .
H2a1 обнаружеили у представителя хвалынской культуры , жившего 6700 лет назад .
H2a3 обнаружили у представительниц анатолийского неолитического поселения Кумтепе , живших 6700 лет назад .
H2a1a определили у представителя среднестоговской культуры из Александрии (Alexandria) (Украина), жившего 6200 лет до настоящего времени .
H4 определили у обитательницы пещеры близ израильской деревни Пкиин , жившей ок. 4000 лет до нашей эры .
H2 определили у образца PG2004 с Северного Кавказа (Progress 2, 6090 лет до настоящего времени, Eneolithic steppe) .
H2b, H6a1b, H13a1a1a определили у представителей ямной культуры .
H2a определили у представителя культуры Ремеделло , жившего ок. 5300 лет назад .
Гаплогруппа H определена у представителей культуры воронковидных кубков .
Митохондриальная гаплогруппа H прослежена у обитателей верховий Западной Двины с 5120±120 лет назад ( ) до VIII—X веков нашей эры (представитель культуры длинных курганов — H2) .
H2a, H2a5, H5a1, H5e1a1, H6a1a, H7a, H10b, H13a2b5, H16, H17, H40 определили у представителей культуры Злота (2900—2500 лет до н. э.) из Польши .
H4a1 и H5b определили у неолитических обитателей болгарской Дзулюницы (2800 лет до н. э.) .
H6a1a определили у представителя фатьяновской культуры эпохи бронзы HAN004 (2835—2471 лет до н. э., Ханево , Московская область) .
H2a определили у обитателя пещеры (Бадахшан, Афганистан), жившего ок. 4,5 тыс. лет назад .
H2b определили у фараона Аменхотепа III (1388—1353/1351 до н. э.) .
H6a1a2a определили у образца I2051 (3260 лет до настоящего времени) из Краснодарского края (Marchenkova Gora, D13, дольмен, поздний бронзовый век) .
H23 определили у представителя лужицкой культуры , жившего 1113—1021 лет до нашей эры .
H101 определили у представителя тасмолинской культуры железного века KSH002.A0101 (Karashoky, 894—790 гг. до н. э.) из Казахстана, H6a1b определили у тасмолинца TAL005.A0101 (Taldy, 789—548 гг. до н. э.) .
H6a1a определили у представителя гальштатской культуры из Чехии (DA111, 2630 ± 48 лет до настоящего времени ) .
H13, H5 и H6b определены у мумий из Абусира .
H4a1 определили у мумии из музея Ольстера (Белфаст, Северная Ирландия). Женщина с 33 зубами жила в Луксоре более 2600 л. н. (около 660 года до н. э.), эпоха 25 династии. Современное распространение H4a1 является редким и спорадическим и было выявлено в таких районах как Канарские острова, Южная Иберия и Ливан. Гаплогруппа H4a1 также обнаружена в древних образцах из Германии — культуры колоколовидных кубков и унетицкой культуры (эпоха бронзы) .
H45 определили у этрусского образца CSN009 из (коммуна Чивителла-Паганико ) в провинции Гроссето региона Тоскана (427—265 лет до н. э.) .
H2 определили у мероитского образца MIS-TM и позднемероитского образца MIS-TMT из некрополя Миссиминия (Missiminia Necropolis) в районе Абри (Abri) в Верхней Нубии (350 год до н. э. — 350 год н. э.) .
H44a определили у этрусского образца TAQ020 Тарквинии в провинции Витербо региона Лацио (Центральная Италия, имперский период, 89—236 годы) .
H1c22 определили у раннеаварского образца CSBper9 (конец IV — начало V века) с высоким компонентом .
H56 определили у раннеаварского образца ANper286 (620—660 гг.) с высоким компонентом WHG .
H и H1af2 определили у славянских женских образцов RISE568 и RISE569 из Брандысека (район Кладно , Чехия), датированных VII—VIII веками нашей эры (660—770 годы) .
H1cf, H1e1a9, H2, H3 и H4a1e определены у древних коренных жителей Канарских островов ( гуанчей ), живших в VI—XIV веках. Наличие линий, производных от H1e1a и H4a1, как в европейском неолите, так и в древних образцах с Канарских островов, соответствует евразийским доисторическим вторжениям в Северную Африку .
H16 определили у четырёх братьев (VK483, VK485, VK490, VK497) из корабля (VIII век) в волости Сальме (Эстония). H1a определили у образцов VK482 и VK496, H1b — у образца VK495, H1b5 — у образца VK492, H1n+146 — у образца VK509, H1q — у образца VK498, H2a2a1 — у образца VK493, H2a2b1 — у образца VK512, H5c — у образца VK488, H6a1a — у образца VK484, H10e — у образцов VK510 и VK552, H17a — у образца VK487, H28a — у образца VK504 .
Гаплогруппа H обнаружена у 5 викингов с языческого захоронения Галгедил (Galgedil) на датском острове Фюн (700—1100 года) .
H6a1a4 определили у образца VK466 (X—XI века) из Гнёздова , H7a1 — у образца VK224, H13a1a1c — у образца VK223, H63 — у образца VK222 .
H5a2a определили у образца VK542 (XI век) из Чернигова (предполагаемые останки князя Глеба Святославича ) .
H1b1 определили у образца VK18 (X—XII века) из Старой Ладоги , H3h — у образца VK409, H5 — у образца VK218, H6c — у образца VK20 .
H1q определили у образца urm160 (church 1 (Urmakaren), покрытие ×1.3) из Сигтуны с Y-хромосомной гаплогруппой R1b1a2a1a1-L11*. H1ap1 определили у образца 84005 (cemetery 1 (Nunnan), покрытие ×1.03) из Сигтуны с Y-хромосомной гаплогруппой I1a1b3-Z74* .
H1e1b определили у кривича №5666 из могильника Болшево-1 (первая половина XII века) .
У сына Ефросиньи Мстиславны венгерского короля Белы III определена митохондриальная гаплогруппа H1b .
H2a2a1 (n=2) и H3b6 (n=2) определили у мужчин из Рубленого города в Ярославле (массовое захоронение № 76, 1238 год) .
H7 определили у образца VK541 (XIII век) из Луцка (предполагаемые останки князя Изяслава Ингваревича ) .
H6a1a определили образца с кладбища Чжэньзишань (Zhenzishan) в китайском Шанду (XIII век) .
Гаплогруппа H обнаружена у 73 % человек, похороненных в XIII—XVI веках в некрополе Сан-Мигель-де-Эреносар (San Miguel de Ereñozar) на северо-западе Испании .
Распространение
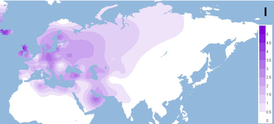
Гаплогруппа H — наиболее распространённая митохондриальная гаплогруппа в Европе — к ней относится более половины современного женского населения Северо-Западной Европы. Данная гаплогруппа также часто встречается в Северной Африке и на Ближнем Востоке. Частота распространения данной гаплогруппы в Европе уменьшается к юго-востоку, составляя всего 20 % на Ближнем Востоке и Кавказе, и менее 10 % в Персидском Заливе, Северной Индии и Центральной Азии.
Среди указанных кладов H1 и H3 подверглись наиболее детальному исследованию; их связывают с мадленской экспансией из Юго-Западной Европы около 13 тысяч лет назад :
Подгаплогруппа H1 составляет значительную долю западноевропейских митохондриальных ДНК, причём пик распространения приходится на басков (27,8 %). Также распространена среди других жителей Иберийского полуострова, Северной Африки и Сардинии . Составляет свыше 10 % во многих других регионах Европы (Франция, Британские острова, Альпы, многие регионы Восточной Европы) и не менее 5 % в прочих местах Европы. Субклад H1b имеет наибольшее распространение в Восточной Европы и на северо-западе Сибири.
Подгаплогруппа H3 составляет значительно меньшую долю «общеевропейского генома», чем H1, однако имеет примерно такое же распространение с максимумом среди басков (13,9 %), галисийцев (8,3 %) и сардинцев (8,5 %). Плотность её падает в направлении северо-востока Европы. Ряд исследований показал, что гаплогруппа H3 связана с весьма высокой сопротивляемостью риску заражения СПИДом.
Оставшиеся субклады встречаются намного реже:
Подгаплогруппа H5 , вероятно, возникла в Западной Азии, где чаще всего встречается в исходном виде. Её субклад H5a больше всего распространён на Центральноевропейских равнинах.
Подгаплогруппы H2, H6 и H8 довольно часто встречаются в Восточной Европе и на Кавказе. Вероятно, это наиболее распространённые субклады гаплогруппы H среди жителей Средней Азии, изредка встречаются и в Западной Азии.
Подгаплогруппы H4, H7 и H13 присутствуют как в Европе, так и в Западной Азии, а последняя — также на Кавказе. Все три указанных субклада довольно редки.
Подклассы
Филогенетическое дерево
Приведенное ниже филогенетическое дерево основано на публикации Ван Овена и последующих опубликованных исследованиях.
-
HV
14766C
-
H
2706A 7028C
-
H1
3010A
-
H1a
73G 16162G
- H1a1 6365C
- H1a2 8271T
- H1a3 16051
- H1b 3796G 16189C 16356C
-
H1c
477C
- H1c1 9150G
- H1c2 12858T
- H2c3 8473
- H1d 456T
-
H1e
5460A
-
H1e1
8512
- H1e1a 14902
- H1e2 15817
-
H1e1
8512
- H1f 4452C 7309C 9066G 16093C
- H1g 8602C
- H1o 267C 485 6446T 11002 14053G 15844
- H1p 13470G
-
H1a
73G 16162G
-
H2
1438A
-
H2a
4769A
-
H2a1
951A 16354T
- H2a1a 6173
-
H2a2
750A
- H2a2a 263A 8860A 15326A
-
H2a2b
16291T
- H2a2b1 16235G
- H2a3 10810C 16274A
- H2a4 11140T
- H2a5 1842G 4592C 13708A 16291T
-
H2a1
951A 16354T
- H2b 152 8598C 16311C
-
H2a
4769A
-
H3
6776C
- H3a (152) 13404C (16239G)
- H3b 2581G
- H3c 12957C
- H3d (152) 73G
- H3e 1618 15592
- H3f 93
-
H4
3992T 5004C 9123A
-
H4a
4024G 14365T 14582G
-
H4a1
8269A
-
H4a1a
10044G
- H4a1a1 73G
- H4a1b 195C 5773A 13889A
-
H4a1a
10044G
- H4a2 7581 15497 15930
-
H4a1
8269A
- H4b 10166C
-
H4a
4024G 14365T 14582G
-
H5
456T 16304C
-
H5a
4336C
- H5a1 15833T
- H5a2 (200) 5839 (16093)
- H5b 5471
-
H5a
4336C
-
H6
239C 16362C 16482G
-
H6a
3915A
-
H6a1
4727G 9380A
-
H6a1a
11253C
- H6a1a1 7325G 16311C
-
H6a1b
10589A
- H6a1b1 6218G 7859A 16284G 16519C
-
H6a1a
11253C
-
H6a1
4727G 9380A
- H6b 16300G
-
H6a
3915A
-
H7
4793G
-
H7a
1719A
- H7a1 16261
-
H7b
5348
- H7b1 12351
- H7c 6296A 16265
-
H7a
1719A
- H8 146C 195 709A 13101C 16288C 16362C
- H9 3591A 4310G 13030C
-
H10
14470A
-
H10a
4216
- H10a1 14548
-
H10a
4216
-
H11
8448C 13759A 16311
-
H11a
961G 16293G
- H11a1 14587 16092 16140
-
H11a
961G 16293G
- H12 3936T 14552G
-
H13
14872T
-
H13a
2259T
-
H13a1
4745G
-
H13a1a
13680T
-
H13a1a1
7337A
- H13a1a1a 13326C
-
H13a1a1
7337A
-
H13a1a
13680T
-
H13a2
709A
-
H13a2a
1008G
- H13a2a1 183G 11151T
- H13a2b 5899.1C 13762G 16311C
-
H13a2a
1008G
-
H13a1
4745G
-
H13a
2259T
-
H14
7645C 10217G
-
H14a
16256T 16352C
- H14a1 146C 7864T 12870T
-
H14a
16256T 16352C
-
H15
6253C
-
H15a
11410
- H15a1 (57G) 14953
- H15b 3847
-
H15a
11410
-
H16
10394T
- H16a 8592A
-
H17
3915A
- H17a 6296T
- H18 13708A 14364
- H19 6272 14869A
-
H20
16218T 16328A
- H20a 249del 292T 16362C
- H21 8994A
- H22 16145A 16227G
- H23 10211T
- H25 9620T
- H26 11152
- H27 16129 11719 16093 16316
- H28 186A 8715 11191
- H29 93 573.1CC 5582
- H30 8628G 14241G 16192T
- H31 (146) (195) 7930T 10771
- H32 73 152 8557
- H33 10211
- H34 15519 16291
- H35 3342
-
H1
3010A
-
H
2706A 7028C
Примечания
- ↑ Achilli et al.(2004), , "American Journal of Human Genetics", 2004 November; 75(5): 911.
- ↑ van Oven, Mannis; Manfred Kayser. (англ.) // : journal. — 2008. — 13 October ( vol. 30 , no. 2 ). — P. E386—E394 . 4 декабря 2012 года.
- ↑ Дата обращения: 28 мая 2009. 4 июля 2008 года.
- . Дата обращения: 22 августа 2011. Архивировано из 28 июня 2011 года.
- . Дата обращения: 22 августа 2011. 14 октября 2013 года.
- . Дата обращения: 12 октября 2013. 13 октября 2013 года.
- ↑ Дата обращения: 28 мая 2009. 4 июля 2008 года.
- Caramelli D. et al. от 3 марта 2016 на Wayback Machine
- Jones, E. R. et al. от 21 августа 2016 на Wayback Machine
- ↑ Szécsényi-Nagy (2015), от 21 июля 2015 на Wayback Machine
- Nikitin, A. G. (2012),
- . Дата обращения: 5 сентября 2015. 5 сентября 2015 года.
- ↑ . Дата обращения: 2 апреля 2015. 4 марта 2015 года.
- ↑ Iain Mathieson et al. от 3 марта 2016 на Wayback Machine
- Дата обращения: 2 января 2016. 19 июля 2017 года.
- ↑ Iain Mathieson et al. от 24 сентября 2017 на Wayback Machine , 2017
- Éadaoin Harney et al. от 20 августа 2018 на Wayback Machine , 2018
- ↑ Chuan-Chao Wang et al. от 9 мая 2020 на Wayback Machine , May 16, 2018
- Morten E. Allentoft , Martin Sikora , Andrey Epimakhov et al. от 30 апреля 2016 на Wayback Machine
- Helena Malmström et al. от 26 января 2021 на Wayback Machine , 2009
- Е. М. Чекунова , Н. В. Ярцева , М. К. Чекунов , А. Н. Мазуркевич . «Первые результаты генотипирования коренных жителей и человеческих костных останков из археологических памятников Верхнего Подвинья». С. 287—294. Таблица на с. 294. // Археология озёрных поселений IV—II тыс. до н. э.: хронология культур и природно-климатические ритмы. — СПб.: ООО «Периферия», 2014.
- Anna Juras et al. от 27 июля 2021 на Wayback Machine , 26 July 2021
- Iain Mathieson et al. от 6 июня 2020 на Wayback Machine , 2017
- Lehti Saag, Sergey V. Vasilyev, Svetlana V. Oshibkina et al. от 23 января 2021 на Wayback Machine (Table 1), July 03, 2020 ( от 30 января 2021 на Wayback Machine )
- Douka K. et al. от 30 августа 2017 на Wayback Machine , Journal of Human Evolution, Volume 107, June 2017, Pages 86–93
- Yehia Z Gad et al. 2020. от 2 февраля 2021 на Wayback Machine // Guardian of Ancient Egypt: Studies in Honor of Zahi Hawass, Volume I. Charles University, Prague, Faculty of Arts: 497—518
- Guido Alberto Gnecchi-Ruscone et al. от 18 августа 2021 на Wayback Machine , 26 Mar 2021
- Peter de Barros Damgaard et al. от 21 февраля 2020 на Wayback Machine , 2018
- Verena J. Schuenemann et al. от 30 сентября 2019 на Wayback Machine , 30 May 2017
- от 6 апреля 2021 на Wayback Machine , 27 January 2020 ( от 18 мая 2021 на Wayback Machine )
- ↑ Cosimo Posth et al. от 25 сентября 2021 на Wayback Machine // Science Advances • 24 Sep 2021 • Vol 7, Issue 39
- Yahia Mehdi Seddik Cherifi, Selma Amrani .
- ↑ Zoltan Maroti et al. от 22 января 2022 на Wayback Machine , 2021
- . Дата обращения: 13 февраля 2022. 5 февраля 2022 года.
- Rosa Fregel et al. Mitogenomes illuminate the origin and migration patterns of the indigenous people of the Canary Islands, 2019
- ↑ Ashot Margaryan et al. от 26 марта 2021 на Wayback Machine , 2020 ( от 12 февраля 2020 на Wayback Machine )
- Linea Melchior, Toomas Kivisild, Niels Lynnerup, Jørgen Dissing . от 10 апреля 2022 на Wayback Machine , May 28, 2008
- Maja Krzewińska et al. от 23 апреля 2020 на Wayback Machine , August 23, 2018
- Чернов С. З., Гончарова Н. Н., Семёнов А. С. от 17 октября 2021 на Wayback Machine // Русин, 2021
- Judit Olasz . от 29 сентября 2018 на Wayback Machine , 2018
- Мустафин Х. Х., Энговатова А. В., Альборова И. Э., Тарасова А. А. от 7 марта 2022 на Wayback Machine // Археология Подмосковья. Материалы научного семинара. Выпуск 18. М.: Институт археологии РАН, 2022. С. 111
- Jiawei Li et al. от 22 июня 2022 на Wayback Machine , 14 June 2022
- I. M. Laza, M. Hervella, M. Neira Zubieta, C. de-la-Rúa . от 1 января 2020 на Wayback Machine , 1 December 2019
- Ghezzi et al. (2005), от 28 октября 2016 на Wayback Machine , European Journal of Human Genetics (2005) 13, 748—752.
- от 17 ноября 2006 на Wayback Machine
- ↑ Дата обращения: 28 мая 2009. 19 августа 2009 года.
- Дата обращения: 3 октября 2017. 5 марта 2016 года.
- данный подкласс исключён из версии 3 дерева Ван Овена.
- ↑ Название данного подкласса — согласно Альваресу-Иглесиасу (Alvarez-Iglesias 2009)
Ссылки
Общие сведения
-
- Ian Logan’s
- Provides a phylogenetic tree of global human mtDNA variation.
- YFull MTree 1.02.00 (under construction)
Гаплогруппа H
-
- Rebekah A. Canada’s [www.familytreedna.com/public/h%20mtdna%20haplogroup/default.aspx mtDNA Haplogroup H Project] at
- National Geographic’s , from National Geographic
- Amelia’s
- Genebase’s
- Genebase’s
- Genebase’s
- (недоступная ссылка)
- (недоступная ссылка)
Гаплогруппа H1
-
- The H1 mtDNA Haplogroup Project
| Митохондриальная Ева | ||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||
| L0 | L3 | |||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||
| M | N | |||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| D | E | G | Q | R | A | S | X | Y | N1 | N2 | ||||||||||||||||
| | | | | | | | | |||||||||||||||||||||||
| C | Z | B | F | R0 | P | I | N1a | W | ||||||||||||||||||
| | | | | | | ||||||||||||||||||||||||
| HV | JT | U | K | |||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||
| V | J | T | Устаревшие кластеры | |||||||||||||||||||||||
| Ключевые понятия |

|
|
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
|
Формирование славянских народов — генофонд, история
|
|||||||
|---|---|---|---|---|---|---|---|
| Основное | |||||||
| Генофонд славян |
|
||||||
|
Древнеславянские
археологические культуры |
|
||||||
- 2020-08-29
- 1