Алтухов, Владимир Николаевич
- 1 year ago
- 0
- 0
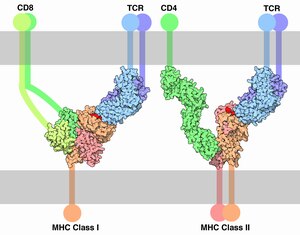
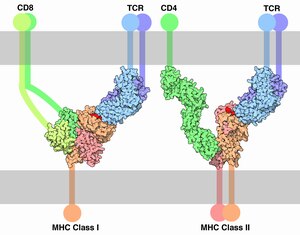
Кореце́птор ( англ. Co-receptor ) — находящийся на поверхности клетки дополнительный рецептор , который связывается с сигнальной молекулой ( лигандом ) в дополнение к первичному рецептору. Корецепторы Т-лимфоцитов усиливают взаимодействие T-клеточного рецептора с комплексом пептид (фрагмент антигена ) + молекула главного комплекса гистосовместимости (MHC). Корецепторы нередко используются вирусами для проникновения в клетку.
Корецептор — это поверхностный рецептор, который после связывания с лигандом взаимодействует с соответствующим первичным рецептором и запускает сигнальный путь в клетке. В иммунологии и биохимии корецепторы также называют дополнительными рецепторами. У корецепторов, как правило, имеется большой внеклеточный домен , на который приходится от 76 % до 100 % всей молекулы рецептора. Именно внеклеточный домен отвечает за связывание с лигандом и формирование комплекса с первичным рецептором. В состав мотивов , отвечающих за эти функции, могут входить гликозаминогликаны , повторы EGF, остатки цистеина или домены ZP-1. Благодаря разнообразным мотивам во внеклеточным доменам корецепторы не только могут связываться с 2—9 различными лигандами, но и взаимодействовать с другими корецепторами. У большинства корецепторов нет внутриклеточного домена, и они связаны с клеточной мембраной посредством GPI -якоря, хотя у некоторых корецепторов всё же есть короткий цитоплазматический домен, лишённый киназной активности. Так как внутриклеточного домена у корецепторов чаще всего не бывает, то сами они не могут запускать сигнальные пути в клетке — это делают первичные рецепторы .
Лиганды, с которыми связываются корецепторы, могут быть самыми разнообразными: интерлейкины , , фактор роста фибробластов (FGF), трансформирующий фактор роста (TGF), (VEGF) и эпидермальный фактор роста (EGF). Корецепторы клеток тканей эмбриона играют важнейшую роль в образовании градиентов морфогенов , которые необходимы для правильной дифференцировки тканей . Корецепторы клеток эндотелия усиливают способность клеток к пролиферации и . Кроме того, корецепторы играют важную роль в функционировании иммунной системы и часто используютя вирусами для проникновения в клетку .
В семейство CD входят разнообразные рецепторы иммунных клеток . Представители семейства, как правило, функционируют как корецепторы. Классическим примером может служить корецептор CD4 , обеспечивающий взаимодействие T-хелпера с комплексом -пептид посредством T-клеточного рецептора (TCR). Корецепторы необходимы для активации иммунных клеток и программируемой клеточной гибели активированных иммунных клеток . Показано, что блокада связывания корецептора CD4 с MHC-II приводит к тому, что активная T-клетка теряет возможность к программируемой гибели . Корецептор CD4 состоит из четырёх иммуноглобулиноподобных доменов и заякорен в клеточной мембране посредством одного . Чаще всего рецепторы CD являются мономерами или димерами и выполняют широкий спектр функций. Упомянутый CD4 не только необходим для функционирования T-клеток, но и служит главным рецептором, необходимым для проникновения ВИЧ в клетку (CD4 взаимодействует с поверхностным гликопротеином ВИЧ). CD28 функционирует как костимулирующий корецептор, влияющий на взаимодействие с MHC-II с TCR и CD4. Хотя CD28 усиливает выделение интерлейкина 2 T-клетками при их активация, блокада этого корецептора никак не отражается на способности T-клеток подвергаться программируемой клеточной гибели .
Представители семейства CCR являются рецепторами, связанными с G-белками ( GPCR ), и функционируют как рецепторы хемокинов . Эти рецепторы располагаются преимущественно на иммунных клетках, в особенности, T-клетках . Рецепторы CCR также экспрессируют некоторые клетки нервной системы , в частности, клетки микроглии . Один из наиболее изученных представителей семейства — CCR5 (и его близкий родственник CXCR4 ), который функционирует как корецептор для ВИЧ . Поверхностный гликопротеин GP120 ВИЧ взаимодействует со своим главным рецептором CD4, а CCR5 формирует комплекс с CD4 и ВИЧ, позволяя вирусу проникнуть внутрь клетки. Некоторые штаммы ВИЧ в качестве корецепторов могут также использовать близкие друг к другу CCR2b, CCR3 и CCR8 , однако все штаммы ВИЧ могут использовать CCR5 и близкий к нему белок CXCR4 для проникновения в клетку. CCR5, кроме того, имеет сродство к ( англ. macrophage inflammatory protein, MIP ) и играет роль в развитии воспалительной реакции. Роль CCR5 в возникновении воспаления хуже понятна, чем его роль в проникновении ВИЧ в T-клетку .
Так как корецепторы играют важную роль в передаче сигналов между клетками, они задействованы в развитии многих заболеваний. Мыши , нокаутные по корецепторам, чаще всего нежизнеспособны, и нокаут корецепторов, как правило, является эмбриональной или перинатальной леталью .
Многие заболевания, связанные с корецепторами, вызываются мутациями в кодирующей области генов , кодирующих корецепторы. Например, мутации , служащего корецепторами для гликопротеинов пути Wnt , которые регулируют , приводят к пониженной костной массе и развитию остеопороза . Некоторые случаи гигантизма могут быть связаны с корецептором, известным как .
Белок CEACAM1 (от англ. Carcinoembryonic antigen cell adhesion molecule-1 ) функционирует как корецептор, необходимый для адгезии эпителиальных , эндотелиальных и гемопоэтических клеток. Он связывается с VEGF и играет важную роль в формировании кровеносных сосудов . Мыши, лишённые гена Caecam1 , не имеют интенсивного роста сосудов, необходимого для питания опухолей , и образуют меньше оксида азота NO, поэтому Caecam1 может быть мишенью противораковой терапии . Важную роль в ангиогенезе в опухолях играют корецепторы семейства нейропилинов , связывающиеся с VEGF наряду с рецепторами / VEGFR2 и плексином .
Корецептор CD109 является отрицательным регулятором рецептора фактора роста опухолей TGF-β . При связывании с TGF-β рецептор интернализуется в составе везикул под действием CD109 , который снижает передачу сигнала через него. Таким образом, корецептор может функционировать как регуляторная молекула, снижающая способность клеток к росту и миграции, которые необходимы для развития опухоли .
Корецепторы синдекан 1 и 4 задействованы в развитии разнообразных видов рака ( рак шейки матки , молочной железы , лёгкого и толстой кишки ), причём ненормальный уровень экспрессии этих белков соответствует худшему прогнозу .
Помимо описанных выше корецепторов CD4, CCR5 и CXCR4, необходимых для проникновения ВИЧ в T-лимфоцит, другие корецепторы также могут быть задействованы в проникновении вирусов в клетку-мишень. Так, для вируса гепатита C необходим корецептор CD81, кроме того, проникновение этого вируса в клетку может регулировать белок плотных контактов .
Блокада корецептора CD4 с помощью антител может использоваться для того, чтобы снизить уровень активации T-клеток при аутоиммунных заболеваниях .