Знаете ли вы/Архив/12
- 1 year ago
- 0
- 0
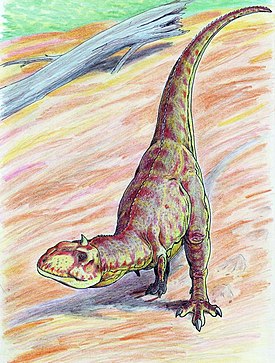
Майюнгаза́вр ( лат. Majungasaurus , «ящер из Махадзанги ») — род тероподовых динозавров из семейства абелизаврид , чьи ископаемые остатки найдены в слоях маастрихтского яруса верхнемеловой системы (72,1—66,0 млн лет назад) на территории Мадагаскара . В род включают типовой и единственный вид Majungasaurus crenatissimus .
Как и прочие абелизавриды, майюнгазавр был среднеразмерным бипедальным хищником с короткой мордой. Хотя его передние конечности полностью не известны, они были очень маленькими, а задние были короткими и коренастыми. Этого динозавра можно отличить от других абелизаврид по более широкому черепу, очень грубой текстуре утолщённой кости на вершине морды, а также по одному скруглённому рогу на крышке черепа , который изначально был принят за купол пахицефалозавра . У него также было больше зубов на верхней и нижней челюстях, чем у большинства представителей семейства.
Известный по нескольким хорошо сохранившимся черепам и множественному скелетному материалу, майюнгазавр стал одним из наиболее изученных теропод Южного полушария . Похоже, что он более тесно связан с абелизавридами из Индии , чем из Южной Америки или континентальной Африки , и этот факт имеет важные биогеографические последствия. Майюнгазавр был сверххищником в своей экосистеме , который охотился в основном на завропод , таких как рапетозавры , а также является одним из немногих динозавров, для которых есть прямые доказательства каннибализма .
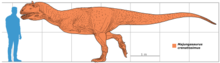
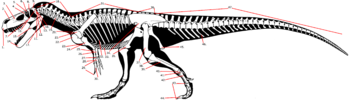
Майюнгазавр был тероподом среднего размера, длина которого составляла 6—7 метров . Фрагментарные остатки крупных особей указывают, что некоторые взрослые животные могли достигать длины 8—11 метров :39 . оценил массу динозавра в 750 килограммов , Сэмпсон и Уитмер дали более внушительную оценку — 1100 килограммов . Однако, образец FMNH PR 2100, на котором они провели измерения, был не самым большим из найденных. Более крупные экземпляры майюнгазавра сравнимы по размеру с родственным карнотавром :39 , который, по оценкам, весил 1500 килограммов .
Череп майюнгазавра известен исключительно хорошо по сравнению с большинством теропод и в целом похож на черепа других представителей семейства. Как и у других абелизаврид, его длина пропорционально короткая относительно высоты, хотя и не такая короткая, как у карнотавра. Черепа крупных особей имели длину 60—70 сантиметров. Типичной для семейства является высокая предчелюстная кость , которая визуально сильно притупляла кончик морды. Тем не менее, череп майюнгазавра был заметно шире черепов других абелизаврид. Черепа всех абелизаврид имеют грубую скульптурированную поверхность наружных граней костей, и майюнгазавр не является исключением. Эта анатомическая особенность доведена у него до крайности: его носовые кости были чрезвычайно толстыми, сросшимися с низким центральным гребнем, проходящим вдоль ноздрей. Отличительный куполообразный рог выступал из слитых лобных костей на верхней части черепа. В жизни эти структуры были покрыты неким покровом, возможно, состоящим из кератина . Компьютерная томография черепа показала, что как рог, так и носовая структура содержат пазухи, возможно, служившие для уменьшения веса головы :39 .
Посткраниальный скелет майюнгазавра очень схож со скелетами карнотавра и Aucasaurus — единственных абелизаврид с полным известным скелетным материалом. Майюнгазавр был бипедальным животным с длинным хвостом, уравновешивающим голову и туловище, с центром масс в районе бёдер . Хотя шейные позвонки содержали многочисленные полости (плевроцели) для уменьшения веса, они были крепкими, с увеличенными участками крепления мышц, и шейными рёбрами для силы. К шейным рёбрам крепились окостенелые сухожилия , придавая им раздвоенный вид — точно так же, как у карнотавра. Все эти особенности делали шею животного очень сильной и мускулистой. Уникальной особенностью шейных рёбер майюнгазавра были длинные углубления по сторонам для снижения веса :129—133 . Плечевая кость была короткой и изогнутой, очень похожей на таковую у карнотавра и Aucasaurus . Так же, как и у родственных видов, у майюнгазавра были очень короткие передние конечности с четырьмя сильно редуцированными пальцами, состоящими из одной (I и IV пальцы) или двух (II и III пальцы) фаланг :58A . На костях плюсны и пальцев у майюнгазавра, как и прочих представителей подсемейства , отсутствовали характерные ямки и бороздки, к которым крепились когти и сухожилия, а кости пальцев были слиты воедино, что указывает на то, что кисть животного была неподвижной . В 2012 году был описан образец лучшей сохранности, показывающий, что нижняя часть передней конечности была крепкой, хотя и короткой, и что кисть состояла из 4 плюсневых костей и 4, вероятно, несгибающихся и сильно редуцированных пальцев с небольшими когтями на II и III пальцах .
Как и у других абелизаврид, задние конечности майюнгазавра были коренастыми и пропорционально короткими по сравнению с длиной туловища. Его большеберцовая кость была ещё более короткой, чем у карнотавра, с заметным гребнем на колене. Таранная и пяточная кости были слиты воедино, а на ногах было 3 функциональных пальца и маленький I палец, который не касался земли .

Впервые ископаемые остатки неизвестного теропода, найденные на северо-западе Мадагаскара, описал французский палеонтолог Шарль Депре в 1896 году. Фоссилии включали в себя два зуба, коготь и несколько позвонков, обнаруженных на берегу реки Бецибука офицером французской армии и хранящихся ныне в коллекции . Депре отнёс эти образцы к роду мегалозавр ( Megalosaurus ), который в то время был мусорным таксоном , куда причисляли остатки всех крупных теропод, и ввёл в систематику новый вид Megalosaurus crenatissimus . Видовое название происходит от лат. crenatus — зубчатый, с добавлением латинского суффикса , означающего превосходную степень , -issimus и указывает на большое количество зубцов на переднем и заднем краях зубов . Позднее Депре отнёс этот вид к североамериканскому роду дриптозавр ( Dryptosaurus ) — ещё одному малоизученному таксону .

В течение следующих ста лет в провинции Махадзанга на северо-западе Мадагаскара французские исследователи обнаружили многочисленные фрагментарные остатки, многие из которых выставлены в Национальном музее естественной истории в Париже :8 . В 1955 году французский палеонтолог описал челюстную кость теропода (MNHN.MAJ 1) с зубами из геологической формации , расположенной в том же регионе, где был найден оригинальный материал. Зубы соответствовали тем, которые впервые описал Депре, но заметно изогнутая кость челюсти сильно отличалась от таковой у мегалозавра и дриптозавра. На основании этой кости Лавокат ввёл в систематику новый род Majungasaurus , названный по старому названию провинции Махадзанга, с добавлением греч. σαυρος/sauros — ящер . В 1979 году и описали фрагмент черепа в форме купола (MNHN.MAJ 4) как новый вид пахицефалозавра Majungatholus atopus . Это была первая находка пахицефалозавра в Южном полушарии .
В 1993 году учёные из Университета штата Нью-Йорк в Стоуни-Брук и Университета Антананариву начали Проект бассейна Махадзанги — серию экспедиций по изучению окаменелостей и геологии отложений позднего мела вблизи деревни Беривотра в провинции Махадзанга :9—11 . Среди этих учёных был палеонтолог из Стоуни Брук. Первая экспедиция обнаружила многочисленные ископаемые остатки меловых позвоночных, среди которых была изолированная предчелюстная кость малоизученного в то время майюнгазавра, которая была описана в 1996 году . В ходе последующих экспедиций были обнаружены десятки тысяч окаменелостей, многие из которых представляют новые для науки виды :9—11 .

Полевые работы 1996 года позволили обнаружить удивительно полный череп теропода, сохранивший мельчайшие детали ( FMNH PR 2100). На вершине этого черепа было куполообразное утолщение, почти идентичное тому, который описали Зюс и Таке под именем Majungatholus atopus . В 1998 году род Majungatholus переназначили в семейство абелизаврид. Хотя название Majungasaurus crenatissimus старше, чем Majungatholus atopus , авторы посчитали типовой образец майюнгазавра (зубную кость) слишком фрагментарным, чтобы уверенно отнести его к тому же виду, что и череп . Дальнейшие полевые работы, проведённые в течение следующего десятилетия, выявили серию менее полных черепов, а также десятки частичных скелетов, начиная от неполовозрелых особей и заканчивая взрослыми. Участники проекта также собрали сотни изолированных костей и тысячи выпавших зубов майюнгазавров. Вместе эти остатки представляют собой почти все части скелета, хотя большая часть передних конечностей, таза и кончик хвоста до сих пор неизвестны :9—11 . Эта полевая работа завершилась монографией 2007 года, состоящей из семи научных работ по всем аспектам биологии животных, опубликованных в «Мемуарах ». Документы написаны на английском языке , хотя у каждого есть введение, написанное на малагасийском . В этой работе зубная кость, описанная Лавокатом, была пересмотрена и определена в качестве диагностической для этого вида. Динозавра перестали рассматривать как пахицефалозаврида, а родовое название Majungatholus было заменено более старым названием Majungasaurus . Хотя монография является всеобъемлющей, редакторы отметили, что она описывает только материал, извлечённый с 1993 по 2001 годы. Значительное количество образцов — некоторые очень полные — были выкопаны в 2003 и 2005 годах и ожидают препарации и описания в будущих публикациях : .

Майюнгазавра классифицируют как представителя семейства (клады) теропод абелизаврид (Abelisauridae). Вместе с семейством ноазаврид (Noasauridae) абелизавриды входят в надсемейство абелизавроидов (Abelisauroidea), которое, в свою очередь, входят в состав инфраотряда цератозавров (Ceratosauria) :2 . Абелизавриды известны своими высокими черепами с укороченной мордой, обширной рельефной поверхностью лицевых костей ( конвергентно с кархародонтозавридами ), очень маленькими атрофированными передними конечностями (конвергентно с тираннозавридами ) и коренастыми задними конечностями .
Систематика семейства абелизаврид во многом запутана. Несколько исследований показали тесную связь майюнгазавра с южноамериканским карнотавром , в то время как другие исследователи не могли уверенно систематизировать животное . В 2007 году авторы описания восстановили майюнгазавра в политомии с Rajasaurus и Indosaurus из Индии , исключив при этом южноамериканские роды, такие как Carnotaurus , Ilokelesia , Ekrixinatosaurus , Aucasaurus и Abelisaurus , равно как и Rugops из материковой Африки :11—16 .
В 2013 году группа учёных под руководством Тьерри Тортозы поместила этого динозавра в новое подсемейство . В 2016 году палеонтологи во главе с Леонардо Филиппе включили майюнгазавра в это же подсемейство, при этом самыми родственными для него оказались Rajasaurus и Indosaurus . Приведённая кладограмма отражает результаты филогенетического анализа внутри семейства абелизаврид в соответствии с этой работой .
| Abelisauridae |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
.jpg)
Пожалуй, самой характерной чертой майюнгазавра была орнаментация черепа: выпуклые, слитые носовые кости и рог на лбу. Другие представители группы цератозавров , такие как карнотавр , цератозавр и Rajasaurus , несли на голове гребни. Эти костные структуры, вероятно, играли роль во внутривидовой конкуренции, хотя их точная функция в этом контексте неизвестна. Полость лобного рога майюнгазавра ослабляла эту структуру и, вероятно, исключала её применение в прямом физическом бою, хотя рог, возможно, служил в демонстрационных целях . В то время как есть различия в орнаментации разных особей майюнгазавра, нет никаких доказательств полового диморфизма у этих животных .

Учёные предположили, что уникальная форма черепа майюнгазавра и других абелизаврид указывает на несколько иные хищнические привычки, чем у прочих теропод. В то время как большинство теропод характеризовалось длинными, низкими и узким черепами, черепа абелизаврид были выше, шире и короче . Узкие черепа других теропод имели строение, позволяющее противостоять вертикальному напряжению, возникающему при укусе, но не так хорошо выдерживали скручивание . Если сравнивать с современными хищными млекопитающими, большинство теропод, вероятно, использовало стратегию, схожую в некоторой степени с таковой у длинно- и узкомордых псовых , которые многочисленными укусами ослабляют свою жертву .
Вместо этого абелизавриды, и особенно майюнгазавр, могли использовать стратегию охоты, сходной с таковой у современных кошачьих с короткими и широкими мордами, которые кусают один раз и удерживают добычу, пока она не ослабеет. У майюнгазавра ещё более широкая морда, чем у прочих абелизаврид, и в пользу гипотезы укуса и удержания могут говорить другие аспекты его анатомии. Его шея была усилена крепкими позвонками, переплетёнными шейными рёбрами и окостеневшими сухожилиями; помимо этого, места прикрепления мышц к шейным позвонкам и к задней стороне черепа также были очень сильными. Черепа абелизаврид также были усилены минерализованными костями, выступавшими за поверхность кожи и создававшими характерную грубую текстуру головы. Особенно хорошо это видно у майюнгазавра, чьи носовые кости слиты и утолщены. С другой стороны, нижняя челюсть этого динозавра имела большие окна с каждой стороны, как и у других цератозавров, а также между определёнными костями, которые обеспечивали высокую степень гибкости нижней челюсти, хотя и не такую высокую, как у змей . Такие особенности анатомии могли служить для предотвращения перелома челюсти при удержании сопротивляющейся жертвы. Передние зубы верхней челюсти были более крепкими, чем остальные, и обеспечивали опорную точку для укуса, в то время как низкие коронки майюнгазавра препятствовали ломке зубов во время борьбы. Наконец, в отличие от зубов аллозавра и большинства других теропод, которые были изогнуты как спереди, так и сзади, у абелизаврид были зубы, изогнутые на передней кромке, но более прямые на задней (режущей) кромке. Благодаря такому строению зубы хищника крепко вцеплялись в плоть жертвы, не разрывая мясо на куски . Исследование зубов майюнгазавра показывает, что его зубы менялись от 2 до 13 раз чаще, чем у других теропод, заменяя весь зубной ряд в течение 2 месяцев. Причиной для такой быстрой смены зубов, возможно, служила привычка животного разгрызать кости .

В своей экосистеме майюнгазавры были сверххищниками , а единственными крупными растительноядными животными были завроподы , такие как рапетозавры . Учёные предположили, что майюнгазавры, и, возможно, прочие абелизавриды специализировались на охоте на завропод. Анатомические адаптации головы и шеи для стратегии «укус и удержание» могли быть полезны против завропод, которые были очень сильными животными. В пользу этой гипотезы говорят короткие и коренастые задние конечности хищника, отличные от более длинных и тонких ног большинства других теропод. Хотя такие ноги не позволяли майюнгазавру двигаться так же быстро, как другие тероподы, это не создавало для него проблем в преследовании медлительных завропод. Прочные кости задних конечностей предполагают очень сильные ноги, а их более короткая длина могла бы понизить центр тяжести животного. Таким образом, майюнгазавр, возможно, жертвовал скоростью ради силы . Следы зубов майюнгазавра на костях рапетозавра подтверждают, что он, по крайней мере, питался этими завроподами независимо от того, действительно ли он их убивал .
Несмотря на то, что завроподы были основой рациона майюнгазавра, существуют находки, говорящие в пользу присутствия в их диете особей своего вида. Было обнаружено множество костей майюнгазавра с отметинами зубов, аналогичными найденным на костях завропод в тех же местах. Эти отметины имеют тот же размер, то же расстояние и несут более мелкие царапины, соответствующие пильчатой насечке на зубах майюнгазавра. Поскольку майюнгазавры — единственные крупные тероподы Мадагаскара, самое простое объяснение состоит в том, что они питались другими особями своего вида . Предположения о том, что триасовый теропод целофиз был каннибалом, опровергнуты, в результате чего майюнгазавр стал единственным динозавром с подтверждёнными склонностями к поеданию себе подобных , хотя имеются некоторые свидетельство существования каннибализма у других видов .
Неизвестно, активно ли охотился майюнгазавр на особей своего вида или только обгладывал из туши . Однако некоторые исследователи отмечают, что современные комодские вараны иногда убивают друг друга, когда соревнуются за доступ к мёртвым телам жертв. После этого вараны продолжают поедать остатки своих соперников, что может указывать на подобное поведение у майюнгазавра и других теропод .

Палеонтологи реконструировали дыхательную систему майюнгазавра на основе серии великолепно сохранившихся позвонков (образец UA 8678) из формации Мейварано. Большая часть этих позвонков, а также некоторые рёбра содержали полости, которые могли быть результатом инфильтрации лёгких и воздушных мешков , по принципу современных птиц. У птиц шейные позвонки и рёбра заполнены шейным воздушным мешком, верхние грудные позвонки заполнены лёгкими, а нижняя часть спины и крестцовые позвонки — брюшным воздушным мешком. Подобные особенности в позвонках майюнгазавра предполагают наличие таких же воздушных мешков. Эти воздушные мешки, возможно, позволили создать базовый вид «сквозной вентиляции» в птичьем стиле, где воздушный поток через лёгкие односторонний, так что обогащенный кислородом воздух, вдыхаемый извне, никогда не смешивается с выдыхаемым воздухом. Такой метод дыхания хотя и сложен, но очень эффективен .
Распознавание пневматических отверстий у майюнгазавра, помимо понимания строения его дыхательной системы, имеет более масштабные последствия для эволюционной биологии . Расхождение на эволюционном дереве между ветвью цератозавров, ведущей к майюнгазавру, и ветвью, ведущей к тетанурам , к которой принадлежат птицы, произошло в истории теропод очень рано. Поэтому дыхательная система птиц, присутствующая в обеих линиях, должна была развиться до расхождения и задолго до эволюции самих птиц. Это является дополнительным доказательством динозаврового происхождения птиц .
.jpg)
Компьютерная томография черепа майюнгазавра (образец FMNH PR 2100) позволила провести грубую реконструкцию структуры его мозга и внутреннего уха . В целом, мозг был очень маленьким по отношению к размеру тела, но в остальном сходен со многими другими не- целурозавровыми тероподами, причем его очень консервативная форма ближе к мозгу современных крокодилов, чем птиц. Одним из различий мозга майюнгазавра от мозга прочих теропод было наличие очень маленького ( лат. flocculus ) — области мозжечка, которая помогает координировать движения глаз с движениями головы. Это говорит о том, что майюнгазавр и другие абелизавриды, такие как Indosaurus , у которого также был маленький клочок, не полагались на быстрые движения головы, чтобы видеть и схватывать добычу .
Выводы о поведении также можно извлечь из обследования внутреннего уха. Полукружные каналы во внутреннем ухе помогают удерживать равновесие, а боковой полукруглый канал обычно параллелен земле, когда животное держит голову в позе тревоги. Когда череп майюнгазавра повёрнут так, что его боковой канал параллелен земле, весь череп почти горизонтален. Это контрастирует со многими другими тероподами, где голова была сильнее опущена, когда находилась в состоянии тревоги. Латеральный канал у майюнгазавра также значительно длиннее, чем у его более базального родственника цератозавра , что указывает на большую чувствительность к движениям головы из стороны в сторону .
В исследовании 2007 года описаны патологии костей майюнгазавра. Учёные исследовали окаменелости 21 особи и у 4 обнаружили заметные патологии . Ранее патологические образования были изучены у аллозаврид и тираннозаврид ; это первое проведённое исследование палеопатологий абелизаврида. На элементах черепа не было обнаружено никаких ран, в отличие от тираннозаврид, у которых иногда встречались ужасные укусы на морде. Одним из образцов была фаланга стопы, которая, по-видимому, была сломана и впоследствии зажила .
Больше всего патологических образований имеется на позвонках. Например, грудной позвонок неполовозрелой особи показывает патологический вырост на нижней стороне кости. Вероятно, рост был результатом преобразования хряща или связки в кость во время развития, но причину окостенения определить не удалось. Гипервитаминоз витамина А и костные шпоры были исключены, а остеому исследователи посчитали маловероятной. Другой аномальный рост тканей наши на другом образце — маленьком хвостовом позвонке. Этот позвонок имеет аномальное образование на невральной дуге . Подобные выросты на невральных дугах позвонков были обнаружены в образцах аллозавра и Masiakasaurus . Вероятно, они образовались в результате окостенения связки, проходящей либо между невральными дугами, либо вдоль их верхушек .
Наиболее серьёзная патология была обнаружена в серии из 5 крупных хвостовых позвонков. Первые 2 позвонка показывают только незначительные аномалии, за исключением большой борозды, которая тянется вдоль левой стороны обоих позвонков. Однако следующие 3 позвонка были полностью слиты во многих разных точках, образуя сплошную костную массу. Других позвонков после пятого нет, что указывает на отсутствие у этого животного почти всего хвоста. Исходя из размера последних позвонков учёные пришли к выводу, что были утрачены около десяти позвонков. Одним из объяснений этой патологии является тяжёлая физическая травма, которая привела к потере части хвоста, за которой последовал остеомиелит последних оставшихся позвонков. Возможно также, что инфекция возникла первой и привела к тому, что часть хвоста поразил некроз, и хвост отвалился. Это первый пример усечения хвоста, известный у нептичьих теропод .

Майюнгазавр, известный по многим хорошо сохранившимся экземплярам разного возраста, хорошо изучен в отношении роста и развития. На протяжении всего онтогенеза череп динозавра становился выше и крепче. Особенно это касалось скуловых , и костей. Кроме того, швы между костями черепа срастались, а глазницы становились пропорционально меньше. Такие изменения указывают на смену рациона животных при взрослении .
Исследование, которое провели Мишель Д'Эмик и его коллеги, показало, что майюнгазавр был одним из самых медленно растущих теропод. Изучив линии роста нескольких костей, учёные установили, что этому динозавру требовалось около 20 лет — большая часть жизни — для достижения максимального размера. Животное, чей образец изучили палеонтологи, к 27 годам достигло длины в 6 метров и массы 850 килограммов. Для примера, тираннозавр набирал 800 килограммов всего за несколько лет, достигая массы в 8—9 тонн к концу жизни. Такой медленный рост животного, вероятно, был обусловлен суровыми условиями жизни: климат мелового Мадагаскара изобиловал засухами и наводнениями. Однако было обнаружено, что и другие абелизавриды также имеют сравнительно медленные темпы роста, что могло быть общей особенностью группы .
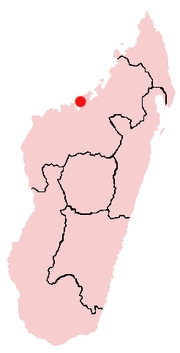
Все образцы майюнгазавра были извлечены из геологической формации Мейварано на северо-западе Мадагаскара. Большая их часть, включая наиболее сохранившиеся скелеты, были найдены в стратиграфическом горизонте Анембалемба, хотя зубы этого динозавра были также обнаружены в подстилающем горизонте Масоробе и перекрывающем горизонте Миадана. Несмотря на то, что эти отложения не подвергались радиометрической датировке, данные биостратиграфии и палеомагнетизма предполагают, что они образовались во время маастрихтской эпохи , которая длилась от 72 до 66 миллионов лет. Зубы майюнгазавра находят до самого конца маастрихта, вплоть до мел-палеогенового вымирания .

В позднемеловую эпоху, как и сейчас, Мадагаскар был островом, отколовшимся от Индийского субконтинента за 20 миллионов лет до этого. Он дрейфовал на север, находясь всё ещё на 10—15 ° южнее, чем сегодня. Преобладающий климат того времени был полузасушливым, с выраженной сезонностью по температуре и количеству осадков. Майюнгазавры населяли прибрежную пойму , прорезанную множеством песчаных речных каналов . Убедительные геологические данные говорят о том, что в начале сезона дождей эти каналы периодически заполнялись потоками мусора, закапывая туши организмов, погибших в предшествующий сухой сезон и обеспечивая их исключительную сохранность в виде окаменелостей . Уровень моря в этом районе повышался на протяжении всего маастрихта и будет продолжаться в эпоху палеоцена , поэтому майюнгазавр, возможно, также бродил по прибрежным районам, таким, как приливные равнины . Соседняя формация представляет существовавшую в то же время морскую среду .
Помимо майюнгазавра, ископаемые таксоны, извлечённые из отложений формации Мейварано, включают рыб , лягушек , ящериц , змей , 7 различных видов крокодиломорфов , 5 или 6 видов млекопитающих , птиц , таких, как Vorona , возможно летающего дромеозаврида Rahonavis , ноазаврида Masiakasaurus и двух титанозавровых завроподов, включая репетозавра . Майюнгазавры, безусловно, были крупнейшими сухопутными хищниками Мадагаскара, хотя некоторые крокодиломорфы, такие как Mahajangasuchus и , могли конкурировать с ним ближе к воде .
|
|
|
|---|