Ионные каналы
- 1 year ago
- 0
- 0

| potassium inwardly-rectifying channel, subfamily J, member 11 | |
|---|---|
| Обозначения | |
| Символы | ; K ir 6.2 |
| HGNC | |
| OMIM | |
| RefSeq | |
| UniProt | |
| Другие данные | |
| Локус | 11-я хр. , |
| ? | |
| ATP-binding cassette, sub-family C (CFTR/MRP), member 8 | |
|---|---|
| Обозначения | |
| Символы | ; SUR1 |
| HGNC | |
| OMIM | |
| RefSeq | |
| UniProt | |
| Другие данные | |
| Локус | 11-я хр. , |
| ? | |
| ATP-binding cassette, sub-family C (CFTR/MRP), member 9 | |
|---|---|
| Обозначения | |
| Символы | ; SUR2A, SUR2B |
| HGNC | |
| OMIM | |
| RefSeq | |
| UniProt | |
| Другие данные | |
| Локус | 12-я хр. , |
| ? | |
АТФ-чувствительные калиевые каналы (англ. ATP-sensitive potassium channels или K ATP channel ) — группа белковых калиевых каналов , активность которых регулируется внутриклеточными нуклеотидами , такими как АТФ и АДФ . Впервые были открыты в клетках сердца . АТФ-зависимые калиевые каналы относятся к классу калиевых каналов внутреннего выпрямления (K ir -каналы) и представляют собой семейство K ir 6. Они работают в основном в мышцах и нейронах , где в условиях недостатка энергии в виде АТФ переводят клетки в состояние покоя, снижая их возбудимость и . Также K ATP -каналы задействованы в регуляции выделения инсулина в бета-клетках поджелудочной железы . АТФ-зависимые калиевые каналы являются мишенями действия лекарственных средств, которые защищают сердце и сосуды во время перегрузки.
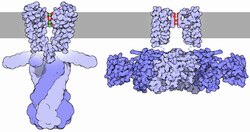
K ATP -каналы — это функциональные октамеры , которые состоят из четырёх Kir6.х субъединиц, формирующих пору канала, и четырёх дополнительных белков — рецепторов к сульфонилмочевине SURx (Sulphonylurea Receprors). Название последней субъединицы обусловлено тем, что при блокировании канала сульфонилмочевиной, она связывается именно с SUR.
K ir 6.x включает в себя два трансмембранных участка и один погружённый в мембрану и формирующий внутреннюю поверхность поры канала с селективным фильтром (P-домен или P-петля). K ir субъединица отвечает за ингибирование канала, то есть поддержание его в закрытом состоянии с помощью АТФ (за исключением каналов в гладких мышцах ).
SUR субъединица состоит из трёх трансмембранных доменов (TMD0, TMD1, TMD2), первый из которых содержит в себе пять, а остальные два — шесть трансмембранных сегментов. Также между TMD1 и TMD2 и после TMD2 на цитоплазматической стороне мембраны находятся нуклеотид связующие домены (NBD1, NBD2). Именно SURx субъединицы отвечают за активацию канала. Они относятся к классу (ATP-binding cassette transporters), основная функция которых — использовать энергию АТФ на нужды клетки, такие как транслокация различных субстратов вдоль мембраны .
Данные ионные каналы имеют высокую селективность в отношении к К + . Селективный фильтр, определяющий какой катион (например, Na + или K + ) может пройти через канал, расположен в самой узкой его части. Исследования показали, что сегмент H5 необходим для селективности к K + . H5 имеет в себе консенсусную последовательность ( Thr - Val - Gly -Tyr-Gly), которая была найдена во всех калиевых каналах с минимальными изменениями в ходе эволюции. Атомы кислорода создают отрицательный заряд в области фильтра, который имитирует водную среду, в которой находится калий в растворе. Считается, что селективность именно к ионам калия обеспечивается жёстко закреплённым размером поры. То есть, другие ионы, такие, как, например, литий не могут пройти (проходят с меньшей вероятностью) потому, что кислород находится от них слишком далеко, гораздо дальше, чем диполи воды, которые окружают ион в растворе, то есть для лития такая «имитация» некорректна.
Для каждой ткани характерна своя конфигурация различных субъединиц. Например, для гладкой мускулатуры сосудов характерен набор K ir 6.1-SUR2B и они образуют подтип K ATP — K NDP , который назван так, чтобы подчеркнуть важность активации с помощью и то, что АТФ сам не может их открыть . В то же время существуют и другие конфигурации, такие как K ir 6.2-SUR2B в мышцах сосудов. Есть сведения о том, что гладкие мышцы мочевого пузыря содержат K ir 6.2-SUR2A(B) . Для сердца характерна конфигурация K ir 6.2-SUR2A. Но если с порообразующей единицей всё чётко и понятно, то с другой возникают некоторые вопросы. Было показано, что у мышей SUR1 экспрессируется в предсердии , в то время как SUR2A — в желудочке . Глюкозо-чувствительные нейроны мозга содержат K ir 6.2-SUR1 субодинцы . Однако также есть исследования, которые показывают, что глюкозо-чувствительные нейроны в вентромедиальном ядре гипоталамуса , а такие же холинергические в полосатом теле мозга содержат в себе конфигурацию из субъединиц K ir 6.1-SUR1. В поджелудочной железе β-клетки содержат K ATP -каналы в конформации K ir 6.2-SUR1 .
Все вариации K ATP кодируются четырьмя генами: KCNJ8, KCNJ11, ABCC8, ABCC9. Они отвечают за субъединицы K ir 6.1, K ir 6.2, SUR1 и SUR2 соответственно. Последняя образует два сплайс варианта и так появляются субъединицы SUR2A и SUR2B. У крысы эти гены расположены в следующем порядке: в первой хромосоме ABCC8 — 39 экзон и KCNJ11 — 2 экзон, в четвёртой хромосоме ABCC9 — 40 экзон и KCNJ8 — 3 экзон.
Главная функция K ATP -каналов в эндокринной части поджелудочной железы — это секреция инсулина . Инсулиновая секреция активируется высоким уровнем глюкозы в крови. Во время допорогового уровня глюкозы K ATP -каналы открыты и поддерживают отрицательный электрохимический потенциал . В момент, когда уровень глюкозы в крови возрастает, запускается метаболизм в β-клетках островков Лангерганса. Как следствие в клетке возрастает уровень АТФ, а уровень АДФ падает. Вслед за этим K ATP -каналы закрываются, клетки деполяризуются и происходит активация потенциалзависимых кальциевых каналов (ПЗКК). Приток кальция ведёт к тому, что пузырьки, которые содержат инсулин, сливаются с мембраной и гормон таким образом высвобождается наружу.
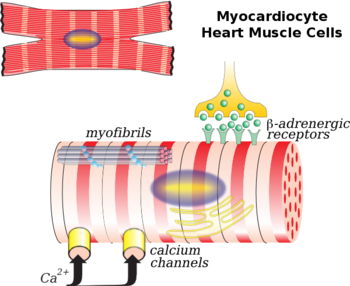
В сердце K ATP -каналы играют роль кардиопротекторов в условиях ишемии . За счёт того, что они гиперполяризуют мембрану, уменьшается амплитуда потенциала действия кардиомиоцитов и ограничивается приток кальция через ПЗКК, соответственно, уменьшается время неэффективного сокращения сердца. В отличие от K ATP -каналов в поджелудочной железе, в сердце они находятся в постоянно закрытом состоянии при физиологических условиях по причине того, что в этой ткани постоянно поддерживается высокая концентрация АТФ. Каналы открываются при таких условиях как перегрузки сердца, гипоксия или ишемия . Даже короткие эпизоды ишемии вызывают дальнейшую защиту миокарда против последующих инсультов . Такой феномен называют « » .
Основное место, где АТФ-зависимые калиевые каналы играют важную роль, — это гипоталамус . В гипоталамусе очень много «глюкозо-чувствительных» нейронов . Так, например, орексинные (гипокретинные) нейроны в мышином латеральном гипоталамусе регулируют бессонницу, локомоторную активность и аппетит и они тормозятся высоким уровнем глюкозы. А MCH-нейроны (Melanin-Concentrating Hormone neurons), которые регулируют поведение поиска пищи, настроение и энергетический баланс, активируются повышенным уровнем глюкозы. Большинство нейронов, которые активируются повышенным уровнем глюкозы работают по следующему механизму: глюкоза попадает внутрь клетки — повышается уровень АТФ — закрываются K ATP -каналы — деполяризация клетки — повышение уровня проводимости. Механизм ингибирования с помощью уровня глюкозы менее понятен на данный момент, однако есть предположение, что в этом принимает участие Na + -K + -АТФаза и активация гиперполяризирующего тока хлора, который, возможно, активирует хлорные каналы (CFTR-like Cl - channels). Кроме этой функции, K ATP -каналы играют защитную роль при патологических условиях . Возвращаясь к ишемии и гипоксии, от которых эти каналы защищали сердце, надо отметить, что у большинства млекопитающих при таких условиях нейроны деполяризуются и умирают. Следовательно, например, в чёрной субстанции K ATP -каналы подавляют нейрональную активность (гиперполяризуют клетку) во время гипоксии за счёт их открытия на постсинаптических мембранах.
В гладких мышцах K ATP -каналы наиболее распространены в стенках сосудов и располагаются в них по всему телу. Открытие этих каналов вызывает гиперполяризацию мембраны и как следствие — закрытие и расслабление мышечных волокон сосудов, особенно вен . То есть, обеспечивают функцию вазодилатации . Кроме обычной регуляции здесь ещё имеет место регуляция открытия-закрытия с помощью фосфорилирования , как уже было указано выше. Так на активность K ATP могут влиять такие вазодилататоры как простагландин , (Calcitonin Gene Related Peptide), аденозин и вазоконстрикторы , например, эндотелин , вазопрессин , гистамин . Возможно, они влияют на сам процесс фосфорилирования с помощью протеинкиназы А. Но, как уже было сказано выше, этот процесс ещё не изучен на достаточном уровне. K ATP присутствуют в мышцах мочевого пузыря. Пока не совсем понятно, зачем там нужны именно эти каналы, ведь кроме них там много других калиевых каналов. Тем не менее, они присутствуют, а их функции и свойства в настоящее время исследуются. Так, было показано, что глибенкламид (ингибитор) не влияет на возбудимость или сократимость мочевого пузыря при отсутствии стимуляции этого канала, тем не менее K ATP способствуют образованию и поддержанию мембранного потенциала покоя клеток мочевого пузыря . При этом за фармакологической активацией K ATP -каналов происходит гиперполяризация мембраны, как следствие — закрытие ПЗКК и снижение напряжения . Несмотря на всё это, маловероятно, что K ATP -каналы связаны с какими-то патологиями в мочевом пузыре, хотя логика и подсказывает другое.
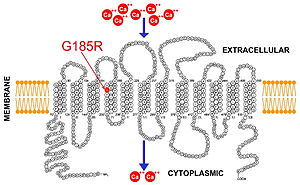
Как уже было отмечено, АТФ является главным регулятором активности канала и может выполнять две различные функции: 1) закрывать канал, и 2) поддерживать канал в активном состоянии в присутствии ионов магния. Сайт для связывания АТФ с целью ингибировать активность канала находится на субъединице K ir 6.2 , это такие остатки аминокислот как аргинин в 50-ом положении на N-конце и 182-й изолейцин , 185-й лизин , 201-й аргинин , 334-и глицин на C-конце . Считается, что эти остатки формируют так называемый АТФ-связывающий карман . То есть, с одним K ATP -каналом может связаться четыре молекулы АТФ. Также АТФ может активировать K ATP -канал, если связывается с SUR , но обычно за активацию отвечает НДФ. Сайт связывания находится на SUR. Последний содержит два NBD (Nucleotide Binding Domain) с такими консенсусными мотивами как Walker-A и Walker-B ( Gly -X-X-X-X- Gly - Lys - Thr / Ser ) . Мутации в Walker-A мотиве на NBD1 предотвращают связывание нуклеотидов с обоими NBD . Такой механизм активации и ингибирования действует при физиологических условиях , однако фармацевтические средства могут как активировать, так и ингибировать K ATP -каналы, связываясь с SUR. Ингибиторными агентами могут быть такие производные сульфонилмочевины как , и глибенкламид . Они используются в лечении сахарного диабета второго типа. Активационные агенты, которые также известны как KCOs (K + channel openers) могут быть представлены такими веществами как пинацидил , никорандил и диазоксид .
С помощью таких производных сульфонилмочевины как , толбутамид, глипизид , глибенкламид и лечат сахарный диабет второго типа.
Показано , что фармацевтические вещества KCOs (K + channel openers) (например, аналог пинацидила [3H]P1075) могут связываться с SUR при отсутствии K ir -субъединицы и не могут активировать канал, если имеется только порообразовывающая субъединица . Также известно, что различные субъединицы SUR по-разному реагируют на активаторы, так в присутствии Mg-АТФ пинацидил может активировать K ir 6.2-SUR2A, но не K ir 6.2-SUR1 . Таким образом показано, что KCOs связываются только с SUR, а также специфичность их связывания. В гладких мышцах KCOs вызывают релаксацию сосудов и снижение кровяного давления за счёт открытия K ATP -каналов. Некоторые из таких веществ используются в терапии как средство против гипертензии . Более того, KCOs используются в качестве лекарств против таких болезней, как острая и хроническая ишемия миокарда, застойная сердечная недостаточность , бронхиальная астма , недержание мочи и некоторых миопатий скелетных мышц . В β-клетках островков Лангерганса поджелудочной железы, вызывая открытие K ATP -каналов, лечат гиперсекрецию инсулина, что связано с инсулиномой и хронической гипогликемией у детей .
Для K ATP -каналов в различных тканях характерны различные уровни проводимости . Так, например, для мочевого пузыря это 11-12 pS (pS — пико сименс ), 15-50 pS — для гладкой мускулатуры воротной вены , культуры клеток коронарной артерии, 55-75 pS — для скелетных мышц, 50-90 pS — β-клетки островков Лангерганса поджелудочной железы, 70-90 pS — для сердца и высокая проводимость (130 pS) характерна для гладкой мускулатуры мезентеральных артерий, хвостовой артерии крысы и аорты собаки. Как уже упоминалось, K ATP -каналы являются семейством из класса K ir , то есть им свойственно входящее выпрямление тока. Выпрямлением тока в электрофизиологии называют явление, когда в условиях одинаковой концентрации иона изнутри и извне мембраны, входящий и выходящий токи не уравновешены. Выпрямление бывает выходным и входным. В нашем случае это входящее выпрямление, то есть калий проще входит в клетку через эти каналы. Также выпрямление бывает слабым и сильным. Для K ATP -каналов характерное «слабое» выпрямление . Также при изменении pH меняется и «сила» выпрямления: при нейтральном pH этим каналам свойственно «слабое» выпрямление, которое, собственно, является физиологическим, при защелачивании среды выпрямление становится более «сильным». Пока ещё не до конца выяснены детали, однако общий механизм выпрямления понятен. Оно происходит посредством алифатического полиамина спермина . Он положительно заряжен и имеет валентность ~5 . Когда калий начинает выходить из клетки он потоком «затягивает» спермин в пору. Пора блокируется и больше не пропускает ионы калия наружу, однако ионы, поступающие внутрь могут «выдавить» его и дальше заходить без помех.