Кошка
- 1 year ago
- 0
- 0


Обоня́ние млекопита́ющих — процесс восприятия млекопитающими присутствия в воздухе , проявляющийся в формировании специфического ощущения (ощущение запаха ), анализа запаха и формирования субъективных ощущений , на основании которых животное реагирует на изменения, происходящие во внешнем мире . За данный процесс у млекопитающих несёт ответственность обонятельный анализатор , возникший ещё на раннем этапе эволюции хордовых .
Обонятельная рецепция складывается из обнаружения пахучего вещества обонятельными хеморецепторами , передачи обонятельной информации в центральную нервную систему и обработку её структурами конечного мозга . Она обеспечивает ряд приспособительных поведенческих реакций : пищевых, половых, социальных, оборонительных, исследовательских .
У большинства млекопитающих обонятельный анализатор представлен двумя сенсорными системами; это — основная обонятельная система и дополнительная обонятельная система . Каждая из них включает три части: периферическую часть (органы обоняния), промежуточную часть (обеспечивает передачу нервных импульсов и состоит из цепи вставочных нейронов ) и центральную часть (обонятельные центры в коре головного мозга ). При этом основной орган обоняния представлен обонятельной областью — ограниченным участком эпителия носовой полости , а дополнительный орган обоняния — якобсоновым органом (иное название: вомероназальный орган ), который представляет собой замкнутую полость, сообщающуюся с полостью рта .
Органы обоняния млекопитающих развиты сильнее, чем у других наземных позвоночных , и играют в их жизни весьма важную роль. Способность к различению запахов млекопитающие используют для ориентирования в пространстве, при поисках пищи, в рамках межвидовых и внутривидовых контактов . О важности обоняния для млекопитающих говорит и то, что наиболее обширное в их геноме образуют как раз гены, кодирующие белки обонятельных рецепторов . Обоняние играет определённую роль и при питании млекопитающих: приятные запахи запускают секрецию слюны и желудочного сока , а неприятные предупреждают о потенциальном вреде (например, неприятный запах испорченной пищи) .
По степени развития обонятельной функции млекопитающих подразделяют на две группы: макросматов с исключительно острым обонянием (большинство млекопитающих) и микросматов с умеренно развитым обонянием ( приматы , китообразные , ластоногие ) .

Различие между данными группами хорошо видно при сопоставлении степени развития обоняния у человека и собаки . Если в носу человека имеется около 6 млн обонятельных клеток, то у собаки их — около 225 млн . Многие макросматы чувствуют запахи на расстоянии нескольких сот метров и способны находить пищевые объекты под землёй . Хорошо известна практика поиска растущих в лесу под землёй трюфелей при помощи специально обученных поисковых собак и свиней , которые способны учуять трюфели под землёй на расстоянии до 20 м .
Степень развития обоняния в целом коррелирует с числом генов , кодирующих различные типы функциональных белков обонятельных рецепторов . У макросматов их обычно более 1000, у многих приматов — около 500, у человека — всего 387 (около 1—2 % генома ), у утконоса — 262 . По-видимому, хуже всего развито обоняние у китообразных; у них же наиболее высок процент псевдогенов обонятельных рецепторов .

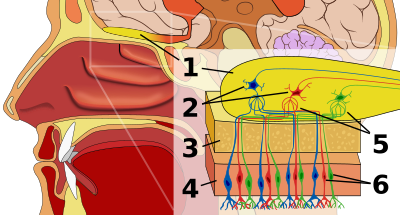
Органы обоняния млекопитающих располагаются в задней верхней части носовой полости , где возникает весьма сложная, особенно у макросматов, система — тонких костных лепестков, направленных внутрь полости и покрытых обонятельным эпителием . В носовых раковинах не только происходит обонятельный анализ вдыхаемого воздуха, но и его нагревание перед поступлением в лёгкие. Среди современных видов четвероногих обонятельные раковины имеются лишь у млекопитающих, а также у немногих видов птиц , у которых эти раковины возникли независимо . Обонятельный эпителий содержит обонятельные рецепторные клетки , опорные клетки, секретирующие слизь и по свойствам близкие к глиальным клеткам, а также базальные клетки, которые, подобно стволовым клеткам , способны делиться и давать начало новым функциональным нейронам в течение всей жизни животного. Размеры обонятельного эпителия у млекопитающих варьируют от 2—4 см² (человек) и 9,3 см² (кролик) до 18 см² (собака) и 21 см² (домашняя кошка). Впрочем, эти значения не дают представления об остроте обоняния, потому что в них не учитывается количество обонятельных рецепторов на единицу поверхности. Обонятельные рецепторы способны улавливать содержащиеся во вдыхаемом воздухе молекулы пахучих веществ. Как и рецепторы вкуса, их относят к группе хеморецепторов . Сигналы о наличии пахнущих веществ передаются через обонятельный нерв в соответствующий центр головного мозга — обонятельную луковицу , или первичные центры обоняния коры головного мозга . Из последней обонятельные сигналы передаются в гипоталамус , лимбическую систему , ретикулярную формацию и неокортекс .
Большинство млекопитающих сохраняют якобсонов орган как обособленный отдел обонятельной капсулы. Этот орган, имеющийся также у двоякодышащих и большинства четвероногих (важнейшие исключения — птицы и крокодилы ), служит главным образом для восприятия феромонов . У представителей ряда групп ( китообразные , сирены , большинство рукокрылых и узконосые приматы , включая человека ) якобсонов орган рудиментарен или полностью утрачен .
Вомероназальный орган выстлан обонятельным эпителием, похожим на тот, что покрывает носовые раковины носа. Обонятельные рецепторные клетки возобновляются в течение всей жизни и поддерживаются эпителиальными и базальными клетками, однако вместо ресничек имеют микроворсинки (микровилли). Рецепторные молекулы также представлены GPCR, однако их аминокислотная последовательность не имеет ничего общего с рецепторами носовых раковин. GPCR вомероназального органа представлены двумя различными семействами, каждое из которых содержит от 100 до 200 генов и развивалось независимо. Представители одного из этих семейств имеют длинный внеклеточный N-концевой домен, сходный с метаботропным глутаматным рецептором . Вторичным посредником здесь служит не цАМФ, как в раковинах носа, а инозитолтрифосфат. Афферентные волокна из вомероназального эпителия проецируются в дополнительную обонятельную луковицу, которая у большинства расположена кзади от основной обонятельной луковицы. Как и обонятельный эпителий носа, вомероназальный эпителий тоже подразделяется на зоны: в апикальной и базальной частях органа экспрессируются разные G-белки. Эти зоны сохраняются и в проекциях в дополнительную обонятельную луковицу: апикальная зона эпителия проецируется в переднюю зону луковицы, а базальная — в заднюю. Гломерулы в дополнительной луковице выражены хуже, чем в основной. Кроме того, вместо пространственной карты основной луковицы представительство дополнительной луковицы более сложное и мозаичное. Дополнительная луковица не имеет проекций в кору и связана только с лимбической системой: с миндалиной и гипоталамическими ядрами, которые играют важную роль в половом поведении. Возможно, дополнительная луковица реагирует только на особые видоспецифические сочетания веществ соответствующего феромона и просто игнорирует все остальные .
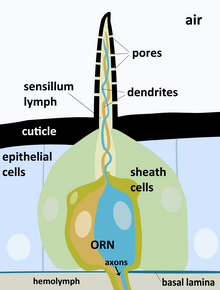
Обонятельные рецепторы ( обонятельные клетки ) представляют собой биполярные нейроны с одним неветвящимся дендритом. Он проходит между базальными клетками и оканчивается небольшим вздутием — обонятельной булавой . Из неё выходят до 20 длинных ресничек, которые представляют собой сенсорную поверхность обонятельной клетки. Они, как правило, погружены в слой слизи, покрывающей эпителий, и образуют с ним плотный матрикс. Обонятельная клетка имеет двойную функцию: восприятие стимула и передача нервного импульса в мозг , поэтому является нейросенсорной клеткой (сенсорным нейроном). Аксоны , выполняющие передачу сигналов в ЦНС , собраны в пучки — обонятельные нити . Обонятельные нейроны способны к замещению путём деления базальных клеток .
Слизь, в которой лежат обонятельные реснички содержит большое количество некрупных (20 кДа) белков , которые секретируются железами носа и встречаются в слизи, покрывающей не только обонятельный эпителий, но и чисто респираторный. Эти белки, возможно, очень неселективно связываются с молекулами пахучих веществ (одорантами) и обеспечивают их взаимодействие с рецепторными клетками .
Обонятельные реснички по ультраструктуре не отличаются от других ресничек и содержат обычную неподвижную аксонему . Обонятельные реснички очень длинные и тонкие: при длине от 5 до 250 мкм они достигают лишь 100—250 нм в диаметре . Они собраны в пучки по 5—40 и выходят из булавы обонятельной клетки, увеличивая её сенсорную поверхность. Рецепторные белки располагаются на поверхности ресничек. Каждый ген из семейства генов, кодирующих такие белки, кодирует определённую их разновидность, и на ресничках одной обонятельной клетки присутствуют обонятельные белки лишь одной разновидности; не все гены данного семейства, впрочем, могут экспрессироваться (так, у человека экспрессируется около 40 % этих генов). Долгое время оставалось неясным, отвечает ли ресничка на многие типы одорантов или только на один . Сейчас, однако, установлено, что обонятельные клетки одного типа специфичны к конкретному узкому классу химических соединений , поскольку распознают в них особые структурные мотивы .
Независимо от специфичности, чувствительность обонятельных клеток очень велика: они способны регистрировать вещества в концентрации от 10 -4 М до 10 -13 М. При простуде чувствительность снижается из-за того, что реснички оказываются погружёнными в слишком толстый слой слизи .
Кроме обонятельных клеток, связанных с обонятельным нервом, в слизистой оболочке носа имеются также свободные окончания тройничного нерва ; они способны реагировать на некоторые агрессивные запахи, например, кислотные или аммиачные испарения .
Проведение обонятельного стимула начинается следующим образом. Пахучее вещество связывается с рецептором в мембране обонятельной клетки. Обонятельный рецептор представляет собой рецептор, связанный с G-белком ( англ. G protein coupled receptor ) и, как и все GPCR, содержит 7 доменов . В отличие от других рецепторов надсемейства GPCR, для обонятельных рецепторов характерно большое аминокислотное разнообразие в 3, 4 и, особенно, 5. Кроме того, от других GPCR обонятельные рецепторы отличаются меньшей специфичностью: они в той или иной степени имеют сродство к целому ряду стереохимически сходных одорантов. Однако небольшим изменениям химической структуры одоранта может соответствовать изменение набора стимулируемых рецепторов и изменение субъективного восприятия. Так, замещение гидроксильной группы октанола на карбоксильную приводит к значительному изменению обонятельного восприятия: вместо запаха, напоминающего запах апельсина , ощущается запах прогорклости и пота. Кроме того, количество стимулируемых рецепторов и субъективное восприятие может зависеть от концентрации одоранта. Например, в низкой концентрации индол имеет приятный цветочный аромат, а в высокой — отвратительный гнилостный .
Связывание одоранта с рецептором активирует , который активирует фермент аденилатциклазу , в результате чего ГТФ распадается на фосфат и ГДФ . Аденилатциклаза переводит АТФ в цАМФ , который связывается с циклонуклеотид-зависимым катионным каналом в мембране и открывает ток ионов Na + и Са 2+ в обонятельную клетку, тем самым запуская потенциал действия в ней, который затем передается на афферентные нейроны . Иногда обонятельные рецепторы, правда, активируют не аденилатциклазу, а фосфолипазу , и вторичным посредником выступает не цАМФ, а инозитолтрифосфат и диацилглицерин . Кроме того, возможно, что в обонятельных клетках из-за активации кальцием NO-синтазы образуется NO , который приводит к образованию цГМФ .
Циклонуклеотид-зависимые каналы имеют шесть гидрофобных сегментов и по структуре напоминают потенциал-зависимые ионные каналы . Отличие заключается в наличии у циклонуклеотид-зависимых каналов большого С-концевого цитоплазматического домена, который связывается со вторичными посредниками. На ресничках расположено 2400 каналов/мкм² (на обонятельной булаве и дендрите лишь 6 каналов/мкм². В отсутствие кальция циклонуклеотид-зависимые каналы проницаемы для всех одновалентных катионов : Na + > K + > Li + > Rb + > Cs + . При воздействии одоранта ионные токи через циклонуклеотид-зависимые каналы изменяются, приводя к деполяризации мембраны клетки и запуску потенциала действия .
Обонятельные клетки одного типа передают свои сигналы в один и тот же обонятельной луковицы, и пространственная организация последней топографически повторяет расположение рецепторов на поверхности обонятельной раковины . Стоит отметить, что один обонятельный рецептор может быть возбуждён одной молекулой пахучего вещества .
В 2004 году Линда Бак и Ричард Эксел получили Нобелевскую премию в области физиологии и медицины за исследования обонятельных рецепторов млекопитающих ; именно они установили химическую природу белков обонятельных рецепторов, дали оценку числа генов в геноме млекопитающих, кодирующих эти белки, и обосновали правила, по которым одна обонятельная клетка экспрессирует одну разновидность белков обонятельных рецепторов, а за обработку сигналов всех обонятельных клеток одного типа отвечает один и тот же клубочек обонятельной луковицы .
Интересно, что циклонуклеотид-зависимые каналы обонятельных ресничек не , т. е. не утрачивают чувствительности при повторных предъявлениях одоранта. Однако в обонятельных клетках, тем не менее, происходит адаптация. Вероятно, это происходит из-за входа в клетку ионов Ca 2+ , которые или непосредственно, или через активацию кальмодулина приводят к закрытию ионных каналов и, кроме того, десенситизируют GPCR .
Кроме того, ответ на обонятельный стимул градуален, то есть большей концентрации одоранта соответствует больший ответ. Это связано с тем, что цАМФ увеличивает или уменьшает количество открытых циклонуклеотид-зависимые каналов. Для эффективного различения сигналов в реальном времени необходим быстрый ответ. Показано, что пик образования цАМФ наступает через 40—75 мс после воздействия пахучего вещества и через 100—500 мс падает до нуля. G-белковый каскад усиливает сигнал, благодаря чему один импульс одоранта активирует множество каналов. Впрочем, кинетика каналов достаточно медленна, и открытое состояние может отставать от импульса цАМФ на несколько миллисекунд. При продолжительной активации GPCR одорантов импульсы цАМФ обеспечивают поддержание циклонуклеотид-зависимые каналов в постоянно открытом состоянии .

В отличие от других сенсорных систем ( зрительной , слуховой , соматосенсорной и, в меньшей степени, вкусовой ), где сенсорный эпителий регистрирует пространственную информацию, «картирующая» функция обонятельного эпителия не так выражена. Тем не менее, в нём присутствуют зачатки пространственной организации. Картирование при помощи , которая даёт возможность выявить активные клетки, показало, что в обонятельном эпителии имеются группы клеток, связанные с определёнными запахами. Так, бутанол возбуждает клетки передней области, а лимонен активирует клетки задней части слизистой. Кроме того, недавно было показано, что рецепторные клетки организованы в передне-задние полосы (так называемые зоны экспрессии ), каждая из которых содержит полный набор клеток. Судя по всему, существует 3 неперекрывающиеся зоны экспрессии, которые перекрываются с меньшей, четвёртой зоной .
Аксоны обонятельных биполярных клеток объединяются в несколько десятков пучков, каждый из которых содержит несколько сотен или тысяч волокон. Они входят в полость черепа через отверстия решётчатой кости и объединяются в обонятельные нервы . Окончания первичных обонятельных клеток образуют синапсы с дендритами клеток обонятельных луковиц. Каждая такая клетка ( ), являющаяся сенсорным нейроном второго порядка, получает сигналы от около 1000 аксонов первичных сенсорных клеток, то есть около 1000 обонятельных аксонов конвергируют на разветвлениях апикального дендрита одной митральной клетки. Около 25 таких дендритов совместно с терминалями формируют сферические образования — гломерулы . На одной гломеруле конвергирует около 2500 обонятельных аксонов, а в обонятельной луковице кролика имеется около 2000 гломерул. Для митральных клеток характерна ритмическая активность, обусловленная вдыханием пахучих веществ. Локальные интернейроны обонятельных луковиц (перигломерулярные клетки, расположенные между гломерулами, и зернистые клетки, залегающие под митральными клетками) способны к контрастированию получаемых сигналов. На этих клетках оканчиваются пути противоположной обонятельной луковицы, лимбических структур и ретикулярной формации мозга. Система синаптических контактов в обонятельной луковицы чрезвычайно сложна, как и её химия: в ней идентифицировано около дюжины нейромедиаторов , среди которых ацетилхолин , дофамин , ГАМК и несколько нейропептидов .
Аксоны митральных клеток образуют обонятельный тракт, ведущий к обонятельным центрам высшего порядка, который, разделяясь на несколько частей, оканчивается на лимбических структурах переднего мозга: , перегородке, пириформной и парагиппокампальной извилинах. От этих структур информация поступает в гиппокамп , миндалины , орбитофронтальную кору (напрямую или через таламус ) и ретикулярную формацию среднего мозга .
Распознавание конкретного запаха является результатом совместной работы рецепторов и мозга, в результате чего он представляется как комбинация «первичных запахов». В соответствии со стереохимической теорией обоняния Монкриффа — Эймура , у человека имеется семикомпонентная система распознавания запахов, базирующаяся на различении семи первичных запахов: мускусного, камфарного, цветочного, эфирного, мятного, едкого и гнилостного (относящиеся к одной группе вещества сходны в стереомодели) .
У человека генетический анализ выявил несколько дюжин специфических аносмий — расстройств обонятельной системы, проявляющихся в неспособности различать определённые запахи. Например, неспособность определить запах цианида встречается с частотой 1:10, а (пахучее вещество скунса ) — 1:1000. Вероятно, аносмии обусловлены дефектами специфических обонятельных рецепторов. Многие аносмии демонстрируют менделевское наследование , однако генетика аносмий изучена плохо .
{{
cite journal
}}
: Википедия:Обслуживание CS1 (множественные имена: authors list) (
ссылка
)