Водоросли
- 1 year ago
- 0
- 0

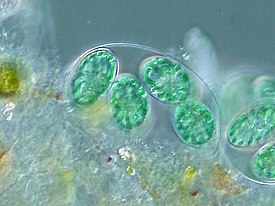
Криптофи́товые во́доросли , или криптомона́ды , или криптофи́ты ( лат. Cryptophyta ), — группа одноклеточных эукариотических фотосинтезирующих организмов , включающая около 165 видов , которой традиционно присваивают ранг типа . Почти все криптофитовые имеют монадную форму с дорсовентральным строением, несут два неравных жгутика . Покровы клетки представлены перипластом, имеются стрекательные структуры (эжектосомы). Хлоропласты окружены четырьмя мембранами и содержат редуцированное ядро — нуклеоморф . Основные фотосинтетические пигменты — хлорофиллы a , c 2 , а также каротиноиды и фикобилины . Митоз обычно открытый, без центриолей , размножение в основном — вегетативное ( делением клеток пополам).
Криптофитовые водоросли обитают как в морских, так и в пресных водах, могут вызывать цветение воды .

Начало научному изучению криптофитов положил немецкий естествоиспытатель Христиан Готфрид Эренберг , который в 1831 году в своей статье впервые ввёл имена родов и , не сопроводив их какими-либо словесными описаниями или рисунками. Упомянул эти два рода он и в другой статье (написанной в том же году, но опубликованной в 1832 году ), однако диагнозы обоих родов, обстоятельные описания и информативные рисунки появились только в основном труде Эренберга «Инфузории как совершенные организмы» ( 1838 ) .
В начале и середине XX века значительный вклад в изучение криптофитов внесли альгологи Адольф Пашер , Генрих Скуя и , а в 1980—1990-е серьёзные уточнения в систематике этой группы водорослей сделали Уве Санторе, Д. Хилл и Р. Уэзерби, Дж. Новарино и И. Лукас. В конце XX — начале XXI века особенное развитие получили исследования ультраструктуры криптофитов средствами электронной микроскопии , экология и молекулярная генетика криптофитов .

Большинство криптофитовых — одноклеточные подвижные организмы монадной формы с дорсовентральным строением и жгутиками. Своё название группа получила за очень маленькие размеры (от 3 до 50 мкм ), которые делают их малозаметными ( др.-греч. κρυπτός — скрытый и др.-греч. μόνος — одинокий) . У некоторых представителей в жизненном цикле имеется пальмеллоидная стадия (например, у и ). Лишь один вид — Bjornbergiella hawaiiensis — формирует простые нитчатые талломы . Среди криптофит известны неподвижные в вегетативном состоянии колониальные и одноклеточные коккоидные организмы (однако существование таких форм оспаривается) .
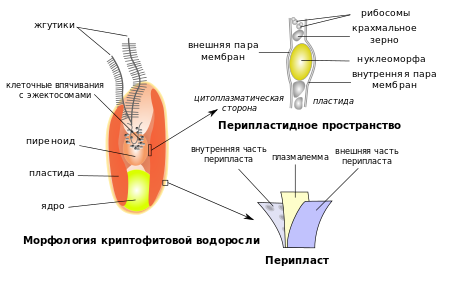
Клетка типичной криптофитовой водоросли уплощённая, с выпуклой спинной и вогнутой брюшной сторонами. Форма клеток разнообразна: яйцевидная, эллипсоидная, грушевидная, бобовидная, веретеновидная. Передний конец клетки скошен, от него отходит продольная борозда, не доходящая до заднего конца. Наличие борозды и её длина используются в качестве признаков для определения родов .
У некоторых представителей на переднем конце тела имеется мешковидная глотка , или вестибулум , или крипта . У разных форм глотка имеет различную длину, она может проходить вдоль или поперёк тела. Иногда она разветвляется; у некоторых видов криптофит глотка укреплена цитоскелетными образованиями — электронно-плотными пластинами. Функциональное значение глотки до сих пор неясно; во всяком случае, у большинства фототрофных форм никогда не наблюдался захват ею каких-либо частиц .

Покровы клеток криптофит представлены перипластом , который состоит из плазмалеммы и двух дополнительных слоёв белкового материала, расположенных над и под плазмалеммой. Внутриклеточная часть перипласта может иметь форму сплошного чехла, как у , или состоять из отдельных пластинок: шестиугольных, как у , или прямоугольных, как у . Прикрепление белковых пластинок к плазмалемме осуществляется за счёт округлых или многоугольных внутримембранных частиц. Наружная часть перипласта может состоять из пластинок, чешуек, слизи или их комбинации. В области глотки перипласта нет , однако закладка новых пластинок перипласта начинается в области рядом с глоткой. Перипласт обеспечивает постоянство формы клетки и предопределяет вид её поверхности: ровный или структурированный (так, у Chroomonas поверхность клетки состоит из прямоугольников, у — из шестигранников) . Клетка может секретировать наружу сульфатированные полисахариды , богатые фукозой .
На переднем конце клетки у монадных форм криптофит имеются два неравных жгутика , по длине сопоставимые с самой клеткой. Оба жгутика отходят от возвышения на краю глотки . Различия в длине между жгутиками невелики: отношения их длин колеблются от 3:5 до 9:10. Оба жгутика заканчиваются более узкой частью — акронемой , в которую заходят лишь две центральные трубочки аксонемы .
Когда клетка движется, то оба жгутика направлены вперёд или же один направлен вперёд, а второй — назад. На жгутиках имеются особые мастигонемы уникального строения, которые могут располагаться в два противолежащих ряда на длинном жгутике (длина мастигонем до 2 мкм), в один ряд на коротком жгутике или же на обоих жгутиках. Мастигонемы криптофит двухчастные и состоят из трубчатой части и тонкого терминального волоска. У некоторых представителей короткий жгутик не несёт мастигонем. У основания длинного жгутика может находиться дополнительный пучок, включающий около 45 волосков. Жгутики могут нести небольшие органические чешуйки в виде семиугольных розеток диаметром 140—170 нм . Известны формы с единственным перистым жгутиком .
Ультраструктура жгутика также имеет свои особенности. В переходной зоне жгутиков имеются две или более пластинчатые структуры (септы) ниже точки, где заканчиваются две центральные микротрубочки . Строение корешкового аппарата варьирует у представителей различных родов, но в общем случае корешковая система представлена ризостилем, который идёт в глубь клетки и состоит из 6—10 микротрубочек, соединённых сократительными фибриллами, трёх микротрубочковых корешков и фибриллярного корешка, прилегающего к одному из микротрубочковых .
Асимметричное расположение жгутиков вызывает несбалансированное вращение клетки вдоль продольной оси при плавании. Чем крупнее клетка и чем более изогнутую форму она имеет, тем более заметным становится такое «раскачивание» .

В клетке криптофитовой водоросли имеется один или два хлоропласта (реже больше), окрашенных в разнообразные цвета: от сине-зелёного и оливково-зелёного до жёлто-бурого, коричневого и тёмно-красного . Хлоропласты могут быть глубоко разделены на две доли или иметь Н-образную форму . Имеются бесцветные формы с деградировавшими пластидами , они ведут гетеротрофный образ жизни .
Хлоропласты криптофит содержат следующие пигменты: хлорофилл а , хлорофилл c 2 и фикобилины: фикоцианин и фикоэритрин (вместе эти пигменты никогда не встречаются, хлоропласт может содержать лишь один из них). За передачу энергии света от фикобилинов к хлорофиллу а отвечает хлорофилл c 2 , а аллофикоцианин , выполняющий эту функцию в клетках красных и синезелёных водорослей , отсутствует. Фикобилины криптофит расположены не в фикобилисомах , а во внутритилакоидных пространствах хлоропластов, поэтому тилакоиды криптофит толще, чем у других водорослей . Выявлено несколько типов фикоцианина и фикоэритрина, отличающихся различным положением максимумов поглощения света; так, в клетках содержится фикоэритрин 566, — фикоэритрин 555, — фикоэритрин 545; у клетки содержат фикоцианин 645, у — фикоцианин 630 (числа характеризуют длину волны в нанометрах, поглощение которой пигментом максимально) .
У криптофит имеется несколько уникальных ксантофиллов , в частности аллоксантин , который можно использовать для их обнаружения в смеси планктонных водорослей, однако главным ксантофиллом является диатоксантин . Имеются также и β-каротин , крококсантин, зеаксантин и монадоксантин .
Соотношение пигментов видоспецифично . Кроме того, оно зависит от интенсивности освещения: чем меньше интенсивность освещения, тем больше фикоэритрина содержится в хлоропластах и тем толще тилакоиды .
Криптофиты характеризуются уникальным типом расположения тилакоидов в хлоропласте. Они собраны в пары или в тройки, причём соседние тилакоиды не имеют соединений . Опоясывающей ламеллы нет. ДНК хлоропласта представлена небольшими нуклеоидами , которые рассеяны в строме хлоропласта .
В хлоропласте может присутствовать пиреноид , в который могут заходить ламеллы из 1—2 тилакоидов . У видов с двудольными хлоропластами пиреноид располагается в месте соединения двух долей . В отличие от хлоропластов красных и зелёных водорослей , хлоропласты криптофит не лежат свободно в цитоплазме , а связаны с другими органеллами (пиреноидом, вакуолями ) .
Хлоропласт криптофитовой водоросли окружён 4 , наружная несёт на своей поверхности рибосомы и продолжается в эндоплазматический ретикулум (ЭПР) и ядерную мембрану . Происхождение хлоропластов криптофит рассматривают как пример вторичного эндосимбиоза , когда бесцветный фаготрофный жгутиконосец, сходный с современной криптофитовой водорослью , «съел» красную водоросль. Поэтому две внутренние мембраны хлоропласта криптофит интерпретируют как мембраны хлоропласта красной водоросли, третью мембрану — как плазмалемму клетки красной водоросли, а четвёртую мембрану — как мембрану пищеварительной вакуоли клетки-хозяина. Между двумя парами мембран расположено перипластидное пространство , в котором располагаются 80S рибосомы, содержащие рРНК эукариотического типа, зёрна крахмала , а также нуклеоморф — сильно редуцированное ядро красной водоросли. Происхождение хлоропластов криптофит от красных водорослей подтверждается анализом 16S рРНК нуклеоморфа и ядер красных водорослей . Две наружные мембраны иногда называют пластидным эндоплазматическим ретикулумом .
Процесс доставки в хлоропласты белков, кодируемых ядерной ДНК, у криптофит организован гораздо более сложно, чем у растений , чьи пластиды представляют собой результат первичного, а не вторичного эндосимбиоза. У криптофит и других организмов, чьи хлоропласты появились в результате вторичного эндосимбиоза, белки, кодируемые ядром, но предназначенные для пластид, содержат на N-конце лидерную последовательность, состоящую из сигнального пептида и переходного пептида. Два этих сигнала совместно обеспечивают прохождение белком всех четырёх мембран для доставки к месту своего функционирования. Было высказано предположение, что в переносе белков в перипластидное пространство принимает участие кодируемая нуклеоморфом ЭПР-связанная система деградации. Поскольку эндосимбионт утратил свой ЭПР, то эта система могла приобрести новую функцию в транспорте белков .
Как уже отмечалось выше, нуклеоморф представляет собой рудиментарное ядро эукариотического фототрофного эндосимбионта , инкорпорированного предками криптофитов в ходе вторичного эндосимбиоза (кроме криптофит, нуклеоморф также имеется у водорослей группы Chlorarachniophyta ; однако в последнем случае происходила инкорпорация не красной, а зелёной водоросли ). У криптофит нуклеоморф обычно имеет округлую или грушевидную форму; он окружён двойной мембраной, содержащей поры, сходные с ядерными поровыми комплексами , и имеет ядрышко с типичными эукариотическими РНК. Нуклеоморф может быть ассоциирован с пиреноидом; иногда он располагается внутри самого пиреноида, иногда — в выросте хлоропласта .
Размер ДНК нуклеоморфа составляет только 660 тыс. пар нуклеотидов в трёх хромосомах (240, 225 и 195 тыс. пар нуклеотидов). Таким образом, геном нуклеоморфа — мельчайший из описанных ядерных геномов эукариот . Интересно, что нуклеоморфы Chlorarachniophyta также имеют 3 хромосомы. У различных видов криптофит ДНК нуклеоморфа содержит от 450 до 850 тыс. пар нуклеотидов. Нуклеоморф имеет 531 ген, которые кодируют 30 белков, функционирующих в хлоропластах. Плотность генов в ДНК нуклеоморфов криптофит довольно высока. Гены нуклеоморфа в большинстве своём являются генами домашнего хозяйства , их продукты участвуют в таких процессах, как фолдинг и разрушение белков, транскрипция и трансляция . В 2001 году был секвенирован геном нуклеоморфа Guillardia theta , а в 2007 году — Hemiselmis andersenii .
Деление нуклеоморфа происходит в препрофазе деления главного ядра вслед за репликацией базального тела , но до деления хлоропласта. Деление амитотическое , и, хотя геном нуклеоморфа содержит гены α-, β- и γ- тубулина , микротрубочки не принимают участия в образовании веретена деления ; вместо этого имеются фибриллярные структуры, появляющиеся только во время деления. Дочерние нуклеоморфы мигрируют в противоположные концы хлоропласта, поэтому каждый дочерний хлоропласт наследует один нуклеоморф. Единственная известная криптофитовая водоросль, не имеющая нуклеоморфа — Goniomonas , которая утратила пластиды. У другой бесцветной криптофитовой водоросли, , имеются лейкопласт и нуклеоморф .
У ряда видов криптофит имеется глазок ( стигма ). У криптофит он связан с хлоропластом, но не связан со жгутиковым аппаратом, а потому может располагаться и в центре клетки, как, например, у Chroomonas mesostigmatica . Стигма состоит из большого количества пигментных липидных глобул , расположенных параллельными рядами под оболочкой хлоропласта. Для некоторых криптофит показан положительный фототаксис . В частности, бесцветный Chilomonas наиболее чувствителен к синему свету при длине волны 366 нм .

Ядра криптофит, как правило, невелики и не превышают в диаметре 3 мкм. Клетка имеет единственное ядро, расположенное в задней части клетки. Для ядер криптофит характерен сложный хромоцентрический тип организации. Хроматиновый ободок (скопление хроматина по краю ядра) может быть сплошным, почти сплошным или же узким и едва различимым, сильно разорванным. Остальное пространство ядра заполнено многочисленными глыбками хроматина, только у Rhinomonas pauca они единичны . У различных криптофит имеется от 40 до 210 хромосом . В 2012 году было завершено секвенирование ядерного генома криптофитовой водоросли , её ядерный геном оказался гаплоидным .
Митоз у исследованных криптофит отличается поразительным сходством и относится к открытому центрическому типу. Процесс деления клеток начинается с удвоения базальных тел. Ядра перемещаются в переднюю часть клетки и иногда вплотную подходят к базальным телам. Базальные тела не меняют своего положения в течение всего митоза и во время метафазы располагаются по обе стороны метафазной пластинки . Они не становятся центриолями, но выполняют их функции, в частности около них формируются многочисленные микротрубочки, участвующие в образовании веретена .
В профазе митоза ядерная оболочка фрагментируется и постепенно растворяется, лишь малая часть её фрагментов сохраняется до конца митоза. Происходит формирование дополнительной перинуклеарной оболочки из хлоропластного ретикулума . Ядрышко на время митоза не исчезает .
Веретено деления криптофит также необычно. Оно состоит из параллельных неконвергирующих микротрубочек. Область полюсов веретена уплощена и ограничена цистернами ЭПР . На его концах, кроме ризостилей, которые отходят от каждой пары базальных тел и не контактируют с микротрубочками веретена, располагаются перемещающиеся туда нуклеоморфы. В метафазе вместо типичной экваториальной пластинки из морфологически дифференцированных хромосом образуется очень компактное скопление хроматина, в котором отдельные хромосомы не различимы, однако образуются особые проходы для микротрубочек веретена. Одно время считали, что истинных хромосом у криптофит нет, а компактную массу хроматина можно рассматривать как одну сборную хромосому. В настоящее время преобладают представления, согласно которым хромосомы криптофит очень мелки, а их кинетохоры морфологически слабо дифференцированы. Роль кинетохоров, по-видимому, выполняют выступы хроматина в области коридоров, которые контактируют с микротрубочками веретена . При делении клеток также происходит формирование новой глотки, которая достаётся одной из дочерних клеток .
Клетки криптофит нередко несут особые стрекательные структуры — эжектосомы, или трихоцисты . Их строение отличается от стрекающих структур динофлагеллят , однако они, возможно, родственны R-тельцам каппа-частиц инфузорий . Крупные эжектосомы располагаются под плазмалеммой в области глотки, а более мелкие разбросаны по всей поверхности клетки. Каждая эжектосома окружена одной мембраной и содержит внутри 2 ленты, скрученные в цилиндры. Более крупный цилиндр состоит из намотанного на спирали лентовидного мембранного материала. В его верхней части находится V-образное углубление, в котором залегает второй цилиндр. По-видимому, эти два цилиндра связаны друг с другом при помощи волокон. При химическом и физическом раздражении скрученные ленты распрямляются и выстреливают, разрывая мембрану эжектосомы и перипласт. Сначала выбрасывается маленький цилиндр, он вытягивает за собой ленту большого цилиндра. Выстрелившая эжектосома с периферии клетки имеет длину 4 мкм, а крупная эжектосома, расположенная около глотки, после срабатывания имеет длину 20 мкм. Выстреливание эжектосом заставляет клетку двигаться в противоположном направлении. Кроме того, при выстреливании из эжектосом высвобождаются вещества, убивающие бактерий, которыми питаются криптофиты . Эжектосомы формируются в пузырьках аппарата Гольджи , причём количество оборотов в цилиндре увеличивается по мере созревания эжектосомы .
С наличием эжектосом связаны трудности по переводу криптофит на лабораторные среды. Их клетки чрезвычайно чувствительны к изменению окружающих условий ( рН , осмотические условия, температура ) и так сильно отстреливают содержимое эжектосом, что клетки быстро разрушаются .
В цитоплазме клеток криптофит имеется одна сетчатая митохондрия с пластинчатыми кристами . Она образует две сети: периферическую (лежащую под плазмалеммой) и центральную (в центре клетки) .
На переднем конце тела у пресноводных криптофит располагаются сократительные вакуоли , изливающие своё содержимое в глотку или борозду . Они служат для регуляции осмотического давления внутри клетки .
В клетках криптофит имеются два тельца Мопа. Тельце Мопа представляет собой крупную везикулярную структуру, расположенную в передней части клетки. Точные функции телец Мопа неизвестны, но, так как они содержат много мембран и фибрилл, они могут принимать участие в разрушении клеточных органелл , особенно отработанных эжектосом, и других инородных частиц в цитоплазме .
Среди криптофит имеются автотрофы , гетеротрофы ( сапротрофы и фаготрофы) и миксотрофы . Большинство криптофит нуждаются в витамине B 12 и тиамине , а некоторые — в биотине . В качестве источников азота криптофиты могут использовать аммоний и органические азотсодержащие соединения, однако морские формы менее способны к превращениям нитрата в нитрит по сравнению с пресноводными представителями. Органические вещества стимулируют рост криптофит. Различные бесцветные формы, а также окрашенный Cryptomonas ovata способны к фагоцитозу , а сине-зелёный Chroomonas pochmannii является миксотрофом. Для захвата и удержания бактериальных клеток используются особые вакуоли. Бактерии втягиваются в них через маленькую пору в глотке, где нет пластинок перипласта, и в вакуолях же перевариваются . Некоторые криптофиты (например, Goniomonas truncata ) могут поедать вирусные частицы . При фаготрофном питании криптофиты демонстрируют хемотаксис в направлении наибольшей концентрации питательных веществ .
Основное запасное вещество у криптофит — крахмал, он откладывается между второй и третьей мембранами хлоропласта; из всех водорослей только криптофиты откладывают крахмал в этом месте. Как и у красных водорослей, он содержит больше амилопектина , чем амилозы , поэтому с йодом даёт красное окрашивание. В цитоплазме могут иметься липидные капли .
Криптофиты размножаются, главным образом, вегетативно — путём деления клетки надвое с помощью борозды деления, причём впячивание плазмалеммы начинается с заднего конца клетки. В большинстве случаев делящаяся клетка сохраняет подвижность. Наибольшая скорость роста для многих криптофит — одно деление в день при температуре около 20 °С .
Половой процесс описан у немногих видов, может протекать в форме изогамии . У описана хологамия , то есть вегетативные клетки функционируют как гаметы . Слияние клеток происходит на брюшной стороне, причём задний конец одной клетки прикрепляется к середине другой. После слияния клеток формируется четырёхжгутиковая зигота , которая делится, вероятно, мейозом .
У Proteomonas sulcata описан гетероморфный гапло-диплобионтный жизненный цикл, в котором имеются как гаплоидная , так и диплоидная стадии. Стадии различаются одна от другой по размеру, структуре перипласта и строению жгутикового аппарата , однако оплодотворение и мейоз у этого вида до сих пор не описаны .
При неблагоприятных условиях (дефицит азота, избыточное освещение) внутри клеток формируются толстостенные цисты .
Криптофиты — планктонные организмы, иногда они встречаются в иле солёных озёр и детрите в пресных водоёмах. Большая часть из них может жить в широком температурном диапазоне, однако некоторые виды Rhodomonas могут жить только при строго определённых температурах. Виды рода Cryptomonas криофильны , они способны жить и размножаться на поверхности льда и снега . Chroomonas africana не утрачивает жизнеспособности при перенесении из морской воды в пресную. Криптофиты устойчивы к загрязнению, они встречаются даже в сточных водах и навозной жиже .
По сравнению с другими водорослями, криптофиты чрезвычайно чувствительны к избыточному освещению, поэтому они часто формируют глубоководные популяции в чистых олиготрофных озёрах. В высокогорных водоёмах и северных озёрах криптофиты встречаются в воде в течение всей зимы. Поскольку лёд и снег пропускают очень мало света, они собираются в поверхностных слоях воды, чтобы получить достаточное для фотосинтеза количество света. Выживание в условиях такой низкой освещённости обеспечивается не только высокой эффективностью фотосинтетической системы, но также низкой скоростью клеточного дыхания при низких температурах и снижением поедания криптофит зоопланктоном в условиях зимы. Весной, когда снег тает и освещённость арктических и горных водоёмов повышается, криптофиты страдают от повышенной освещённости, и большая часть их биомассы уходит в более глубокие слои воды .
Криптофиты нередко совершают суточные миграции в вертикальном направлении с амплитудой менее 5 м. Cryptomonas , обитающие в небольших лесных озёрах, утром демонстрируют положительный фототаксис и перемещаются в верхний слой воды, бедный фосфором . Далее криптофиты уходят из самых верхних слоёв, избегая избыточного освещения, и перемещаются в богатый фосфором нижний слой холодной воды ( гиполимнион ). Этот цикл миграций также помогает криптофитам избежать поедания зоопланктоном .
Криптофиты — доминирующая группа водорослей в покрытых льдами пресноводных водоёмах Антарктики , где они могут составлять до 70 % биомассы фитопланктона . В таких водоёмах наблюдается значительное расслоение воды, поскольку факторы, способные вызывать движение воды (ветер, изменение температуры воды), отсутствуют. Криптофиты доминируют в самых нижних слоях, где в течение зимы они ведут гетеротрофный образ жизни и поглощают одну бактерию в час путём фагоцитоза. Летом криптофиты миксотрофны. Залогом выживания криптофитов в таких условиях является то, что они всё время находятся в вегетативном состоянии, а не уходят в покоящиеся формы. Находясь в вегетативной форме, популяция может быстро отреагировать на улучшение условий и приход короткого антарктического лета . Летом, когда начинается приток воды от таяния ледников, криптофиты вызывают локализованные цветения в антарктических водах . Цветение воды, вызванное криптофитовыми водорослями, было зафиксировано и в Белом море .
Очень редко криптофиты живут в океанических водах при температуре 22 °С и выше, они отсутствуют в горячих источниках и гиперсолёных водоёмах. Пресноводные криптофиты населяют искусственные и естественные водоёмы со стоячей водой, где также избегают сильно освещённых слоёв воды. Бесцветные формы распространены в водах, загрязнённых органикой, и могут служить показателями загрязнения воды органическими веществами .
Среди криптофит имеются виды, населяющие болота с низкими значениями рН, однако ряд видов не обладает избирательностью по отношению к рН: так, Chilomonas живёт при рН 4,1—8,4 .
Некоторые криптомонады обитают в пищеварительном тракте домашних животных .

Криптофиты живут в качестве эндосимбионтов в клетках инфузории Myrionecta rubra . Эта инфузория может вызывать нетоксичное красное цветение воды в зоне апвеллинга . Цвет обусловлен красновато-коричневыми хлоропластами эндосимбионта. Криптофитовая водоросль, находящаяся внутри инфузории, окружена двойной мембраной (плазмалемма водоросли и пищеварительная вакуоль клетки-хозяина) и функционирует как полноценный автотроф. Ей присущи все особенности строения криптофитовых водорослей, но она лишена перипласта, жгутикового аппарата и эжектосом. Инфузория, содержащая эндосимбионтов-криптофит, целиком полагается на них в своём питании и утрачивает клеточный рот .
Некоторые криптофиты являются эндосимбионтами некоторых динофлагеллят (например, ), причём в некоторых случаях у них может сохраняться ядро криптофитовой водоросли, а в других случаях сохраняются только пластиды .
Криптофиты имеют большое значение в круговороте кислорода , углерода , азота и фосфора в водоёмах, в синтезе органики из минеральных веществ. Криптофиты служат пищей для различных представителей зоопланктона, которым, в свою очередь, питаются рыбы . Ветвистоусые рачки интенсивно питаются криптофитами в эвтрофных прудах в зимнее время. Однако цветение воды в прудах, обусловленное Cyanomonas , приводило к массовой гибели сомов .
В настоящее время криптофиты находят применение в исследовательской сфере. Так, недавно открытые родопсины водоросли Guillardia theta , представляющие собой анионные каналы , могут использоваться в качестве очень эффективного оптогенетического метода для подавления нейронов .
На протяжении почти всего XX века классификация криптофитов разрабатывалась протозоологами и ботаниками- альгологами практически независимо. Протозоологи трактовали таксон как отряд Cryptomonadida Calkins, 1926 в составе типа Protozoa ; альгологи — либо как класс Cryptophyceae Pascher, 1914 в составе отдела Pyrrophyta (включавшего также класс Dinophyceae , а иногда — ещё и класс ) , либо как самостоятельный отдел Cryptophyta Silva, 1962 . При этом внутреннюю классификацию криптофитовых водорослей ботаники строили на основе типа организации таллома , выделяя порядки Cryptomonadales (монадные формы) и Cryptococcales (коккоидные формы) . В конце XX века эти ранние классификации полностью устарели.
Исследования в области филогении криптофитов и родственных им групп эукариот , развернувшиеся в конце XX — начале XXI века, нашли отражение в системе типа Cryptysta (тип Cryptophyta в широком понимании), которая была предложена в 2015 году Майклом Руджеро с соавторами и входит как составная часть в представленную ими макросистему живущих организмов. Здесь данный тип рассматривается в составе подцарства (включающего также гаптофитов , центрохелидных солнечников и несколько мелких групп с неясными родственными связями: отряды , , Picomonadida и ). Система доведена до уровня порядков и отрядов (по отношению к классу Cryptophyceae авторы системы следуют правилам ботанической номенклатуры , по отношению к другим классам криптистов — правилам зоологической номенклатуры ) и выглядит следующим образом :
Тип Cryptista Cavalier-Smith, 1989 — Криптисты
Внутри криптофит эволюционно наиболее древними считаются формы, близкие к роду Goniomonas . Представители этого рода лишены пластид и нуклеоморфа, и наличие у них продольной борозды в отсутствие глотки считается наиболее примитивной особенностью. Формирование глотки происходило за счёт смыкания краёв борозды, начиная с заднего конца; этот процесс завершился у Chroomonas , имеющего глотку, но лишённого борозды. В случае Chilomonas произошёл вторичный переход к гетеротрофности: у этого организма редуцирована борозда, развита глотка и имеются лейкопласты. Такая схема эволюционных взаимоотношений подтверждается анализом 18S рРНК .
Статус таксономической группы Hacrobia (а следовательно, и внешние связи криптофитов) до сих пор окончательно неясен. В одних исследованиях группа предстаёт как клада , сестринская по отношению к группе SAR ( страменопилы , альвеоляты и ризарии ) . Но есть и работы, в которых единство группы Hacrobia отвергается; в одной из них центрохелидные солнечники, гаптофиты и телонемиды предстают как последовательные ответвления у основания группы SAR, а криптисты не связаны с ними и оказываются наиболее ранним ответвлением группы Archaeplastida .