Дифференцировка клеток
- 1 year ago
- 0
- 0


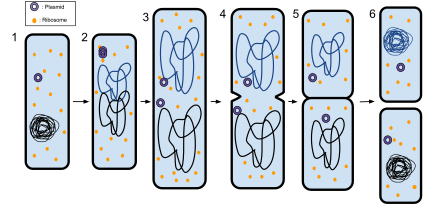
Деление прокариотических клеток — процесс образования дочерних прокариотических клеток из материнской. Ключевыми событиями клеточного цикла как прокариот, так и эукариот являются репликация ДНК и деление клетки . Отличительной чертой деления прокариотических клеток является непосредственное участие реплицированной ДНК в процессе деления . В подавляющем большинстве случаев прокариотические клетки делятся с образованием двух одинаковых по размеру дочерних клеток, поэтому этот процесс ещё иногда называют бинарным делением . Так как чаще всего прокариотические клетки имеют клеточную стенку , бинарное деление сопровождается образованием септы — перегородки между дочерними клетками, которая затем расслаивается посередине. Процесс деления прокариотической клетки подробно изучен на примере Escherichia coli .
Раскрытию механизма деления грамотрицательных бактерий способствовало изучение мутантных штаммов E. coli , у которых этот механизм нарушен. В результате мутаций , которые затрагивают гены , участвующие в делении клетки, могут формироваться следующие фенотипы :

Центральную роль в делении клеток грамотрицательных бактерий играет септальное кольцо — кольцевая органелла, расположенная примерно посередине клетки и способная сокращаться, образуя перетяжку между двумя новыми дочерними клетками. Зрелое септальное кольцо представляет собой сложный белковый комплекс, состоящий более чем из дюжины разных белков. Десять из них (FtsA, B, I, K, L, N, Q, W, Z и ZipA) абсолютно необходимы для формирования септы, и нарушение в их работе приводит к формированию филаментов типа Fts − . Остальные компоненты не являются строго необходимыми, их функции могут частично перекрываться. Формирование септального кольца происходит в несколько этапов, новые белки присоединяются по одному в таком порядке: FtsZ→FtsA/ZipA→FtsK→FtsQ→FtsL/FtsB→FtsW→FtsI→FtsN .
Белки, входящие в состав септального кольца, помимо FtsZ, можно разделить на несколько классов по функциям:
Однако для многих белков септального кольца точная функция до сих пор не известна .
Незрелую форму септального кольца называют Z-кольцом, по имени белка FtsZ, который играет ключевую роль в его формировании. Однако стоит отметить, что часто термины септальное кольцо и Z-кольцо используют как синонимы, поэтому в каждом отдельном случае это нужно оговаривать особо . Белок FtsZ имеет тенденцию формировать длинные фибриллярные структуры. После деления FtsZ формирует прилегающую ко внутренней мембране спираль, закрученную вдоль оси клетки. Эта спираль постоянно меняет своё положение и быстро осциллирует от одного полюса клетки к другому . Примерно ко времени завершения репликации ДНК спираль FtsZ схлопывается, в результате чего формируется Z-кольцо посередине клетки . Есть все основания предполагать, что Z-кольцо на самом деле также представляет собой короткую плотную спираль .
Белок FtsZ — прокариотический гомолог тубулина с похожей третичной структурой . Это позволяет предполагать, что ассоциация FtsZ в Z-кольцо может напоминать сборку микротрубочек эукариот . FtsZ, как и тубулин, обладает ГТФазной активностью , гидролиз ГТФ обеспечивает полимеризацию FtsZ с образованием линейных протофиламентов. Z-кольцо — динамичная структура: молекулы FtsZ в составе кольца постоянно заменяются молекулами из цитоплазматического пула .
FtsZ сам по себе не имеет сродства к мембране , формирование кольцевой структуры из протофиламентов, их закрепление во внутренней мембране и стабилизацию Z-кольца обеспечивают белки FtsA и ZipA, которые взаимодействуют с FtsZ прямо и независимо. ZipA — интегральный белок внутренней мембраны, FtsA — цитоплазматический белок, который тем не менее может связываться с мембраной за счёт особой аминокислотной последовательности на C-конце. ZipA, по-видимому, характерен только для γ-протеобактерий , в то время как FtsA более универсален . Z-кольцо у E. coli может формироваться при отсутствии одного из этих белков, но не двух сразу, что указывает на их перекрывающиеся функции .
Ещё два белка — ZapA и ZapB — включаются в состав Z-кольца на ранней стадии, однако их присутствие не строго обязательно для его формирования . ZapA — универсальный для многих прокариот белок, а вот ZapB, по всей вероятности, есть только у γ-протеобактерий . ZapA связывается с FtsZ непосредственно, а ZapB связывается с ZapA. Интересно, что ZapB формирует ещё одну кольцевую структуру, которая находится дальше от мембраны, чем Z-кольцо. Функции этих белков ещё до конца не установлены, однако предполагается, что они принимают участие в превращении спирали FtsZ в Z-кольцо, а также в последующей стабилизации Z-кольца .
Z-кольцо существует в описанном виде 14—21 минуту (в зависимости от скорости деления), и только после этого к нему присоединяются все остальные ключевые белки, начиная с FtsQ . В какое время присоединяется FtsK, пока точно не установлено. Оставшиеся белки включаются в состав септального кольца практически одновременно в течение 1—3 минут. До того момента, как начинает собираться септальное кольцо, Z-кольцо стимулирует синтез пептидогликана в центре клетки таким образом, что клетка начинает удлиняться. Молекулярный механизм этого процесса, однако, до сих пор не установлен .
Одними из последних в септальное кольцо включаются белки, ответственные за синтез полярного пептидогликана (FtsW, FtsI), и белки, обеспечивающие частичный гидролиз пептидогликана на границе раздела между двумя клетками (AmiA, B, C, EnvC, NlpD) .
Завершающим этапом деления прокариотической клетки является формирование перетяжки и конечное разделение двух новых клеток. Образование перетяжки затрагивает все компоненты клеточной оболочки (внутреннюю мембрану, слой пептидогликана и внешнюю мембрану). Есть основания полагать, что за инвагинацию внутренней мембраны отвечает Z-кольцо, однако как именно оно передаёт напряжение на мембрану, пока не известно. Параллельно с этим процессом ферменты септального кольца синтезируют (или модифицируют особым образом предсуществующий) пептидогликан септы . После формирования септы в работу вступают пептидогликангидролазы, которые отделяют будущие клетки друг от друга. Завершается процесс деления инвагинацией и обособлением внешних мембран клеток.
| Патогенные бактерии | |
|---|---|
|
Микрофлора
человека |
|
|
Субстратная
специфичность |
|
| Дыхание | |
|
Генетика и
размножение |
|
| Покоящиеся формы | |
| См. также | |