Огнёвка кукурузная
- 1 year ago
- 0
- 0

Южная амбарная огнёвка , или индийская моль , или южная огнёвка , или амбарная огнёвка ( лат. Plodia interpunctella ) — вид чешуекрылых из семейства настоящих огнёвок (Pyralidae). Единственный представитель рода Plodia . Бабочки имеют размах крыльев 16—20 мм и хорошо отличаются от других близких видов огнёвок прямыми губными щупиками и двуцветной окраской крыльев — беловато-жёлтой или серо-жёлтой в основании и ржаво-охристой или красно-коричневой в вершинных 2/3 передних крыльев. Опасный вредитель запасов пищевых продуктов. Изредка встречается в гнёздах птиц . Гусеницы могут паразитировать в полостях тела и под кожей позвоночных. Эта огнёвка может провоцировать развитие анафилактической аллергии . Симбиотические бактерии, живущие в кишечнике гусениц, способны разрушать полиэтиленовую и поливинилхлоридную плёнку, и обеспечивают иммунитет к патогенным микроорганизмам. На стадии имаго не питается. Общая плодовитость составляет до 400 яиц. Развитие в зависимости от экологических условий происходит с диапаузой или без неё. При оптимальных условиях в течение года может быть 18 поколений. В кариотипе Plodia interpunctella 30 пар хромосом . В W-хромосоме обнаружен большой специфичный для самок участок гетерохроматина (PiSAT1). Митохондриальный геном состоит 15287 пар нуклеотидов , в которых закодирована последовательность 36 генов. Космополитный и синантропный вид.
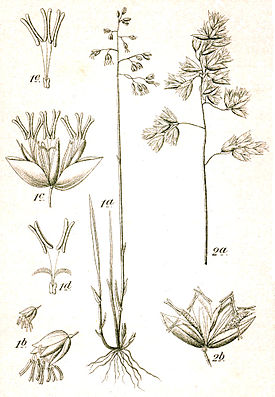
Размах крыльев 13—20 мм . Длина переднего крыла самцов 5,65—5,92 мм, самок — 6,40—6,82 мм . Длина тела 6—8,5 мм . Самки в среднем крупнее самцов . Губные щупики направлены вперёд и немного опущены, с прижатыми чешуйками, а конечный их членик короткий и заострён . Челюстные щупики укорочены, нитевидные. Губные щупальца 3-членистые, сильно выдаются вперёд, наподобие клюва. Голова довольно сильно сплющена сверху вниз, покрыта красновато-оранжевыми чешуйками, которые выступают над лбом в виде конуса. Лоб конусовидный. Глаза чёрные, обрамлены тонкими, довольно длинными щетинками. Простые глазки располагаются у границы между лобной поверхностью и теменем (за основанием усиков). Ротовые органы расположены в нижней части головы. У самцов усики покрыты короткими ресничками, у самок — нитевидные. Грудь и тело красновато-оранжевого цвета. Тегулы (небольшие пластинки, которые прикрывает сверху место прикрепления крыльев к груди) густо покрыты чешуйками. Крылья имеют по краям 3 ряда длинных волосовидных чешуек, которые образуют короткую бахромку. Передние крылья узкие, отношение длины и ширины 1:3,5, с изогнутым передним краем, округло-треугольные, с неразделённой срединной ячейкой. Окраска передних крыльев двухцветная — беловато-жёлтая или серо-жёлтая в области их основания и бурая, ржаво-охряная или красно-коричневая в остальной части. Рисунок на передних крыльях состоит из двух проходящих поперёк крыла тёмно-бурых перевязей со свинцово-бурым блеском и тёмного пятна (в середине радиально-кубитальной ячейки крыла) на конце скрученной ячейки. Задние крылья коротко-овальные, однотонные, грязно-белые или серовато-белые, с затемнённым наружным краем и коричневой каймой. Субкостальная жилка впадает в костальный край переднего крыла за его серединой. Передние крылья без жилок R 3 , R 5 и M 2 , на задних отсутствует только M 2 . Жилка A 2 на передних крыльях не ветвится. На задних крыльях субкостальная жилка на значительном протяжении слита со стволом радиальных жилок. У самцов нижняя сторона передних крыльев имеет заворот, снизу окружённый волосковидными чешуйками . Ноги хорошо развиты. Бедро и голень передних ног короткие. В строении генитального аппарата самца вальвы узкие с загнутым внутрь верхним краем. Перед их вершинами располагается небольшого размера вырост в форме зубца. Наружный край вальвы несёт на себе слабовыраженные зазубрины. Пенис имеет вид простой слабо склерозированной трубки с расширенным основанием, по длине равной вальве. Саккус (отросток, простирающийся внутрь брюшка) широкий, желобовидный. В строении генитального аппарата самки лопасти вагинальной пластинки не соединены между собой. Совокупительная сумка с коротким протоком, конец которого расширен и сильно пигментирован. Её стенки покрыты концентрически расположенными рядами из мелких шипиков .
Яйца беловато-желтоватого цвета и овальной формы. Их размеры составляют 0,4—0,62×0,23—0,28 мм. Оболочка слабо ячеистая. В конце развития яйца становятся мутно-белого цвета, сквозь оболочку просвечивается сформировавшаяся гусеница .
На момент вылупления из яйца гусеницы имеют длину 0,7—1 мм, на последней стадии развития достигают длины 10—16 мм. Тело мягкое, червеобразное, его форма приближена к цилиндрической, суживается к обоим концам. Кожа гладкая, поверхность кутикулы покрыта мелкими, густыми микроскопическими бугорками с плоской вершиной. Окраска тела взрослой гусеницы беловато-жёлтая или зеленоватая. Нижний край головы по середине тёмно-коричневый или чёрный. Лоб занимает 2/3 высоты головы. По бокам головы имеется по шесть, реже пять, простых глазков , при этом 3 и 4 глазки иногда соприкасаются. С каждой стороны у основания головы имеются по одному продолговатому пятну. Верхние челюсти на вершине с тремя зубцами. Самым крупным из них является средний зубец. Дыхальца располагаются на переднегруди и на восьми сегментах брюшка. Самыми крупными является дыхальца восьмого сегмента . Они отчётливо выступают над поверхностью тела и их край со слабо склеротизированной каймой . Тёмные пятна у основания щетинок, в отличие от гусениц огнёвок рода , отсутствуют . Тазики передних ног расставлены на 2/3 своего диаметра, а промежуток между средне- и заднегрудными тазиками примерно равен ему. Брюшко с четырьмя парами ног. Крючки на этих ногах расположены венцом в два круга. Большие крючки в три раза больше мелких .
Куколка длиной 6,2—9 мм . Форма куколки веретеновидная, голова округлая, покровы хитинизированные и плотные. Её окраска изменяется в процессе развития. Первоначально она жёлтая, затем — у неё темнеют глаза, а перед выходом бабочки темнеет вся куколка. Теменной шов на голове хорошо развит. Грудные дыхальца хорошо заметные, округлой формы. Поверхность куколки гладкая. Шипы и волоски на теле отсутствуют . Брюшко гладкое, без ямчатой скульптуры. Между IX и Х сегментами брюшка отсутствует борозда на дорсальной (спинной) стороне куколки .






Встречаются преимущественно в отапливаемых жилых помещениях, на складах, предприятиях пищевой промышленности, в хранилищах, подсобных помещениях и торговых залах продуктовых магазинов, на мукомольных предприятиях, хлебокомбинатах, пивных заводах, кондитерских фабриках, в пекарнях и т. п. Также обнаружены в гнёздах птиц , шмелей , пчёл-листорезов , пчёл антофор ( Anthophora ), бумажных ос ( Polistes ) и пчёл осмий ( Osmia ) .
Бабочки выходят из куколок обычно во второй половине дня или под вечер. Сразу после выхода бабочки обычно сидят около своего кокона или отползают в сторону, взбираются на возвышение, выступы стен и замирают в положении с висящим вниз брюшком до расправления крыльев. Расправление и высыхание крыльев длится обычно 30-40 минут. Расправленные крылья отвердевают и приобретают окончательную окраску. После этого бабочки прячутся или улетают . Лёт бабочек растянут. Они ведут ночной или сумеречный образ жизни, когда в спектре преобладает синий свет (400—475 нм) . В дневное время и во время отдыха они обычно мало подвижны, сидят на стенах, на нижней стороне различных предметов, реже на их боковых сторонах и крайне редко сверху . Летают вечером и в ночное время и обычно не летят на искусственные источники света . Самцы и спарившиеся самки иногда совершают дальние перелёты, это также происходит обычно в сумерках. Самцы более активные и подвижные, чем самки. Бабочки не питаются ( афагия ) и живут за счёт запасов питательных веществ, накопленных на стадии гусеницы. Продолжительность жизни бабочек обоих полов зависит от температуры окружающей среды, а также их участия в процессе спаривании. Она составляет 5-20 дней при спаривании и больше, если бабочки не спаривались. Самки живут меньше самцов .
Появившиеся весной из перезимовавших куколок бабочки обычно спариваются уже в день своего выхода из куколки, по другим источникам — через несколько дней после выхода из куколки . Оптимальными являются температуры от 20 до 30°С. В опытах в температурном интервале между 30 и 35°С активность спаривания увеличивалась при возрастании относительной влажности. При других температурах влияние влажности было менее заметно или не проявлялось. Спаривания при температурах ниже 20°С было более продолжительным, чем при температуре более. При высоких температурах увеличение относительной влажности воздуха приводило к увеличению продолжительности спаривания . Самки могут спариваться несколько раз с разными самцами , а самцы — с несколькими самками .
Как самки, так и самцы выделяют феромоны , которые влияют на половое поведение при поиске партнёра и спаривании . У Plodia interpunctella идентифицировано четыре первичных феромона самок: (Z,E)-9,12-тетрадекадиенилацетат, (Z,E)-9,12-тетрадекадиеналь, (Z,E)-9,12-тетрадекадиенол и (Z)-9-тетрадеценилацетат. Удаление любого компонента смеси снижает число привлечённых самцов . Было высказано предположение, что причина наличия такого большого количества компонентов в феромоне заключается в том, что он обеспечивает видовую специфичность . Самцы также выделяют феромоны. Приблизившись к самке, самец выделяет феромоны из андроконий , расположенных у основания каждого переднего крыла. Эти феромоны заставляют самку оставаться неподвижной в позе с приподнятым брюшком между крыльями, что облегчает спаривание .
Самцы прилетают к самкам. Спаривающиеся бабочки обычно сидят на стенах помещений, обращены головами в противоположные стороны. Самки спариваются с несколькими самцами. Последние имеют ограниченное количество сперматозоидов и в зависимости от различных факторов выделяют разное их количество во время спаривания. Самцы передают большее количество половых клеток самкам, которые ранее уже спаривались несколько раз, чтобы повысить свои шансы на успех в конкуренции сперматозоидов других самцов. Самцы также выделяют больше сперматозоидов при спаривании с более «молодой» самкой .
Созревание яиц в теле самки после спаривания с самцом происходит за два — четыре приёма. В первый раз созревает наибольшее количество яиц, которое и откладывается самкой через 1-2 суток после спаривания . По другой информации самка приступает к откладыванию яиц уже спустя несколько часов после спаривания . При откладке яиц самка стремится распределить их как можно шире по всей поверхности питательного субстрата для гусениц. Отложив первую порцию яиц, самка успокаивается до следующего дня, когда приступает к откладыванию новой кладки. Яйца откладываются одиночно или группами до 30 штук , иногда размер кладки достигает 275 яиц . Общая плодовитость одной самки составляет от 150 до 400 яиц . На неё влияют особенности потребляемой бабочками пищи, наличие воды, а также размеры и физиологическое состояние самки. Биоритмы , вероятно, оказывают слабое воздействие на откладку яиц . Продолжительность эмбрионального развития (стадия яйца) составляет от 2 до 16 дней : от 3—4 (при 30 °С) до 7—8 суток (при 20 °С). При температуре 15°С и ниже яйца не вылупляются . В среднем стадия яйца при благоприятных условиях составляет 4—5 дней .

За время своего развития гусеницы проходят пять-семь возрастов . Выйдя из яйца, гусеницы начинают активно и быстро ползать в поисках подходящего для себя места, например, трещин либо морщинок кожицы плода или зерна, и забираются внутрь них . На 2—3-й день жизни гусеницы сооружают из выделяемой ими шелковистой нити паутинную трубочку и прикрепляет к ней свои экскременты. Находясь в трубочке, гусеницы питаются, прогрызая вокруг себя широкий ход в пищевом субстрате . Они могут покидать свои паутинные трубочки, а при питании в таких пищевых субстратах, как сухофрукты, часто не сооружают их вовсе, а живут внутри сухих плодов. Потревоженные гусеницы сворачиваются кольцом .
Гусеницы могут питаться очень широким спектром сухих пищевых продуктов, главным образом, растительного происхождения: мукой, различными крупами, зерном, пшеницей, рожью, кукурузой, рисом и продуктами их переработки, зерновыми хлопьями, макаронными изделиями, сухими хлебобулочными изделиями, сушёными фруктами и овощами, изюмом, различными орехами, каштанами, желудями, семенами бобовых , включая фасоль и горох , различными семенами, арахисом, лекарственным сырьём, сушёными травами, пряностями , шоколадом и изделиями из него, сухими дрожжами , солодом и другими растительными субстратами . Известны случаи питания гусениц сухими кормами для домашних животных (на растительной основе), птичьим кормом, сухими суповыми смесями, измельчённым красным перцем , какао-бобами , заменителями кофе, печеньем, старыми сотами и пергой пчёл и даже ядовитыми семенами дурмана ( Datura stramonium ) . Предпочитают в первую очередь муку, особенно кукурузную , хотя питание ею приводит к увеличению времени развития бабочек . В зёрнах гусеницы в первую очередь выедают зародыш, и чем суше семя, тем сильнее оно повреждается таким образом . Качество и характер пищевого субстрата оказывают большое влияние на процесс развития гусениц. От них же зависит скорость развития гусениц . Так, из гусениц, питавшихся мукой, выходят бабочки меньших размеров, чем из гусениц, питавшихся сухофруктами .
Гусеницы покрывают пищевой субстрат шелковиной (паутиной), при значительном развитии в нём образуются комки. Продолжительность развития 23—56 суток . Самки развиваются быстрее самцов . Оптимальные температуры развития гусениц лежат в диапазоне от 18 до 35 °С. Порог устойчивости — 10°С .
В кишечнике Plodia interpunctella идентифицированы несколько видов симбиотических микроорганизмов ( YT1 и Bacillus spp. YP1), позволяющие им осуществлять биодеградацию полиэтилена и других пластмасс . Видовой состав и численность микроорганизмов кишечника зависит от состава корма. Помимо участия в пищеварении они могут защищать хозяина от патогенных микроорганизмов, формирую стойкий иммунитет к ним .
Характерной поведенческой особенностью гусениц южной мучной огнёвки является каннибализм . Он является одним из механизмов конкуренции, оказывает влияние на динамику популяции вида. Обычно гусеницы пятого и четвёртого возраста преимущественно поедают здоровых гусениц более ранних возрастов, а также заражённых вирусами гусениц равных с ними размеров. Заражённые гусеницы менее активны и не травмируют гусеницу-каннибала при нападении. Это характерно для гусениц третьего возраста, которые преимущественно нападают на инфицированных особей второго возраста .
Переход в состояние диапаузы происходит после пятого возраста и контролируется продолжительностью фотопериода и температурой, её интенсивность различается у разных линий . Непосредственным пусковым сигналом к диапаузе может быть резкое снижение температуры и продолжительность освещения меньше 13 часов . Диапауза, вызванная температурой около 20 °С, имеет продолжительность не более 4 месяцев, а при температуре выше 25 °С она может удлинятся до 9 месяцев . Максимальная зарегистрированная продолжительность развития 267 суток. Скорость развития гусениц из одной кладки в одинаковых условиях различается. Размер бабочек зависит от особенностей корма. Особи, питавшиеся сухофруктами на стадии гусеницы на стадии имаго, как правило, крупнее выросших на муке . Развитие может происходить и без диапаузы.
Последний возраст гусеницы отличается большой продолжительностью . В его начале гусеницы активно двигаются и питаются, а спустя 5-7 дней при благоприятных условиях перестают питаться , покидают кормовой субстрат и перемещаются от 12 до 48 часов в поисках подходящего места для образования кокона . Окукливаются в полупрозрачном коконе цилиндрической формы. Наиболее часто окукливаются в щелях и трещинах в стенах и полу, открыто на стенах, в складках мешков с кормовым субстратом, иногда — в кормовом субстрате, которым питалась гусеница .
Куколка развивается от 7 до 31 суток . Продолжительность её развития зависит от температуры окружающей среды. Количество поколений, способных развиться за год находится в зависимости от продолжительности времени, на протяжении которого сохраняются условия, необходимые для развития гусениц. При оптимальных условиях в течение года максимально может развиваться до 18 поколений . Наблюдается устойчивая зависимость количества поколений от климатических факторов места обитания, в первую очередь от температурных показателей. На севере ареала только одно поколение . Зимуют гусеницы последнего возраста в коконах, реже — куколки .

Гусеницы южной амбарной огнёвки являются хозяевами паразитических клещей перепончатокрылых и двукрылых . Среди таких паразитов отмечены ихневмониды ( , , , , = Diadegma kiehtani ), бракониды ( Bracon hebetor , , ), птеромалиды ( ), муравьи ( Iridomyrmex purpureus , ), тахины ( , ). Хищниками южной амбарной огнёвки являются клопы семейства ( ), клещи семейства ( ) .
Паразитами гусениц Plodia interpunctella являются нематоды родов и . Эти паразиты проникают в тело хозяина через рот, дыхальца или анальное отверстие и развиваются на протяжении двух-трёх поколений. Непосредственной причиной смерти являются выделения бактерий- мутуалистов spp., которые были обнаружены в кишечнике нематод. Смерть гусениц происходит в течение 48 часов . В лабораторных условиях смертность от нематод может составлять от 62,25 % до 71,25 % .
Южная амбарная огнёвка восприимчива к вирусу гранулёза, который относится к семейству бакуловирусов . Заражаются гусеницы на ранних стадиях развития, а личинки последнего возраста либо устойчивы к вирусу или могут содержать сублетальные концентрации патогена . Передача вируса гранулёза может происходить как горизонтально (от особи к особи), так и вертикально (от родителей к потомкам). Было показано, что и самки и самцы способны передавать сублетальную инфекцию своему потомству . Одним из способов передачи вируса в популяции является каннибализм . Отмечено также поражение микроспоридиями рода и бактериями Bacillus thuringiensis .
В кариотипе Plodia interpunctella 30 пар хромосом. Самцы имеют две одинаковые (ZZ) половые хромосомы, а самки гетерогаметны (ZW) . В W-хромосоме обнаружен крупный (более трети длины) участок гетерохроматина (PiSAT1), представляющий собой специфичную для самок последовательность нуклеотидов. У многих других видов чешуекрылых W-хромосома состоит из чередующихся специфических для самок и общих для обоих полов последовательностей. Выявлено, что этот уникальный для Plodia interpunctella участок является сателлитной ДНК и редко встречается в аутосомах и/или Z-хромосоме. Предполагают, что он может выполнять регуляторную роль через образование некодирующей РНК .
Митохондриальный геном состоит из 15287 пар нуклеотидов, в которых закодирована последовательность 36 генов и контрольной области. В 13 генах зашифрована информация белках, 2 гена производят рибосомальную РНК , 22 гена используются для синтеза транспортной РНК . В нуклеотидом составе 80,15 % составляют нуклеотиды A и Т . Белок-кодирующие гены начинаются обычно с последовательности ATA, ATC, ATG и ATT и только ген первой субъединицы цитохром-c-оксидазы ( cox1 ) может начинаться с кодона CGA. У четырёх белковых генов терминирующий кодоны неполные T или TA. Из генов транспортной РНК только trnS1 (AGN) не образует типичной структуры клеверного листа. Контрольная область состоит из 327 пар нуклеотидов .
Вид описан Якобом Хюбнером в 1813 году как Tinea interpunctella , является единственным представителем рода Plodia Guenée , 1845 . Близкородственными родами являются и .
Вид является космополитом . Современный ареал охватывает преимущественно тропические и субтропические регионы всех континентов . В мире широко распространён в странах Средиземноморья , Западной и Восточной Европы , на Кавказе, в Закавказье, странах Средней и Малой Азии , Северной Африке , Северной Америки , Южной Америки , Австралии . На севере ареала встречается только в помещениях . Распространена южная амбарная огнёвка в том числе в южной части России и стран СНГ , встречаясь также в помещениях среднеширотных и северных регионов . В летние месяцы на юге европейской части России и на Кавказе может летом давать и 1-2 поколения в природе. Южная амбарная огнёвка обычна в складах, подсобных помещениях продуктовых магазинов, на мукомольных предприятиях и т. п.
Существуют две основные гипотезы происхождения и расселения южной амбарной огнёвки. Одни учёные считают родиной вида Средиземноморье , другие предполагают, что он происходит из Северной Америки . В СССР завезена в 1920-х годах. Предполагают, что в Европу этот вид попал из Индии , поэтому его также называют индийской молью . По другой версии, название индийская моль было впервые предложено американским энтомологом в 1858 году, потому что он наблюдал, как личинки питаются кукурузной мукой , которую в то время называли индийской мукой .
Карантинный объект для Венгрии , Чехии , Словакии , Монголии , Китая .

Гусеницы южной амбарной огнёвки являются вредителями самых разнообразных продовольственных запасов в хранилищах, магазинах, пищевых предприятиях и жилых домах. Зарегистрированы в качестве вредителей как минимум 179 различных видов пищевых запасов , включая муку, особенно кукурузную, зерно, рожь, пшеницу, кукурузу, рис и продукты их переработки, сушёные фрукты и овощи, пряности, изюм , шоколад и изделия из него, различные орехи, арахис, лекарственное сырьё, сушёные травы, сушёный табак и табачные изделия , сухие дрожжи . Также могут повреждать гербарные образцы, энтомологические коллекции . Распространяются с заражёнными продуктами, а в пределах населённого пункта или предприятия также путём разлёта бабочек .
Гусеницами чаще всего повреждается верхний слой пищевого субстрата на глубину до 20 см. Повреждённые продукты оплетаются паутиной и засоряются экскрементами. Паутина , выделяемая гусеницами, смешивается с экскрементами и личиночными шкурками , что придаёт заражённым продуктам неприятный запах. При интенсивном поражении паутинный слой может быть очень толстый . В некоторых районах вредоносность южной амбарной огнёвки может быть выше, чем у мучной огнёвки ( Pyralis farinalis ) .
Южная амбарная огнёвка также является одним из опасных вредителей в шмелеводстве . Бабочки могут проникать внутрь гнёзд шмелей , например земляного шмеля ( Bombus terrestris ), и откладывают там яйца . Гусеницы поедают, а также загрязняют своими экскрементами кормовые запасы в гнёздах, а также повреждают расплод шмелей .
Известны случаи паразитирования гусениц в полостях тела некоторых домашних животных. Так живые гусеницы были извлечены из полости головного мозга индийского кольчатого попугая и из подкожной клетчатки домашних кошек .
Гусеницы четвёртого и пятого возраста могут проделывать входные отверстия в полиэтиленовых пакетах толщиной 0,03 мм. Для поиска входных отверстий молодые гусеницы могут преодолевать расстояние до 38 см . Они способны прогрызать также плёнки из поливинилхлорида в течение пяти дней , а через многослойную полипропиленовую плёнку они проникают на 21 сутки экспозиции .
Непосредственный контакт с гусеницами и бабочками, вдыхание фрагментов их тел или продуктов их жизнедеятельности может вызывать у человека различные бронхолёгочные заболевания . Попадание гусениц и их экскрементов вместе с заражёнными ими пищевыми субстратами в пищеварительную систему человека может привести к развитию желудочно-кишечных расстройств. Пищеварительные ферменты гусениц являются аллергенами, которые могут провоцировать развитие аллергических реакций — ринитов, конъюнктивитов, а при попадании на кожные покровы вызывать различные дерматиты . Около половины (51 %) пациентов, страдающих аллергией на домашнюю пыль, имели иммуноположительную реакцию на экстракт этого вида чешуекрылых . Идентифицированными аллергенами являются и тиоредоксин . Белок аргининкиназа является перекрёстно-реагирующим аллергеном. При этом известно, что перекрестные свойства аллергенов могут влиять на клинические проявления возникающих аллергических реакций и на результаты проводимых диагностических проб . Развитие данных аллергические реакций может быть как самостоятельным, так и выступить в качестве этиологического фактора при возникновении бронхиальной астмы , аллергодерматозов, ангионевротических отёков и других заболеваний . Доказано существование чётких связей между плотностью южной амбарной огнёвки и развитием сенсибилизации у больных бронхиальной астмой . Развитие вышеуказанных заболеваний отмечается главным образом у людей, работающих в заражённых огнёвкой складских помещениях и приобретающих товары, поражённые её гусеницами .
Южная амбарная огнёвка используется наряду с другими видами насекомых в качестве белкового корма при разведении беспозвоночных энтомофагов, например в биологических лабораториях при крупных тепличных комбинатах, а также при разведении муравьёв в муравьиных фермах ( формикариях ) .
Профилактические меры борьбы с южной амбарной огнёвкой включают подготовку хранилищ и помещений перед приемкой и размещением на хранение сухого растительного сырья, проведение комплексного обследования всех объектов на зараженность, очистка и дезинсекция при помощи влажной или аэрозольной обработки; подготовка хранимого растительного сырья — сушка до сухого состояния или средней сухости, очистка от сорных примесей, снижение температуры хранения, опрыскивание контактными инсектицидами; проведение постоянного контроля за зараженностью насекомыми-вредителями. Также одним из важнейших условий предотвращения заражения сырья и продукции гусеницами южной амбарной огнёвки является содержание в чистоте складских и производственных помещений .
В качестве традиционного способа борьбы с огнёвкой использовалась фумигация бромистым метилом и фосфином . Производство бромистого метила, который является одним из озоноразрушающих веществ, было ограничено Монреальским протоколом с 1995 года . Были запрещены некоторые инсектициды на основе фосфорорганических соединений и уретанов . Появились устойчивые линии южной амбарной огнёвки к ряду веществ, например малатиону и хлорпирифосу . Устойчивость к фосфину была выявлена в 2016 году . Экологически безопасными фумигантами против южной амбарной огнёвки считаются метилбензоат и сульфурилфторид .
Это всё вынуждает находить альтернативные комплексные подходы к борьбе : прежде всего общесанитарные меры, такие как очистка трещин и щелей в стенах и самих стен от коконов, сортировка заражённого зерна, термическая обработка, хранение продуктов при пониженной концентрации кислорода в воздухе . Рекомендуется хранить продовольственные запасы при температуре ниже +15 °С . Понижение температуры до 10°С приводит к развитию стрессовых реакций, возрастает смертность, снижается плодовитость и жизнеспособность яиц .

В лабораторных экспериментах показана возможность развития устойчивости к энтомопатогенной бактерии Bacillus thuringiensis , хотя нет прямых доказательств что такая устойчивость имеется у природных популяций .
Для контроля популяций Plodia interpunctella проводились лабораторные испытания с использованием аттрактантов , репеллентов , веществ стимулирующих и подавляющих спаривание. Широкого коммерческого применения в качестве мер борьбы эти методы не получили. Для оценки численности используют пищевые аттрактанты и феромонные ловушки . Применяются также клеевые ловушки, содержащие половые феромоны самок, которые способствуют удалению самцов из популяций, что помогает поддерживать численность вредителя на экономически безопасном уровне .
Осуществлялись лабораторные и полевые опыты применения естественных врагов, например яйцевых паразитов рода Trichogramma и нематод рода Heterorhabditis . Показана высокая эффективность Bracon hebetor .
Применяются также вирусы, вызывающие гранулёз . Было обнаружено, что в популяциях, подвергшихся воздействию вируса гранулёза, почти в два раза возрастает устойчивость к инфекции . Новой группой токсикантов , апробируемой для борьбы с южной амбарной огнёвкой, являются пропептиды , которые подавляют активность пищеварительных ферментов гусениц . Для контроля численности предложено также использовать стерилизованных самцов .
Альтернативой методов фумигации является обработка диатомитной пылью и спиносадом . При дозах 500 и 1000 ppm погибали до 86—97 % гусениц первого возраста. Природный инсектицид спиносад уже при дозе 1 ppm приводит к гибели до 97,6—99,6 % .
Для борьбы с гусеницами используется также обработка хранящихся продуктов гамма-излучением в дозах 250 Гр . При этом происходит разрушение клеток эпителия кишечника и снижение численности симбиотических бактерий .