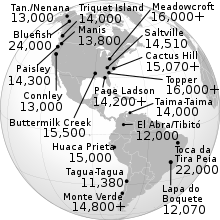
Древние контакты между Китаем и Средиземноморьем
- 1 year ago
- 0
- 0

Межкле́точные конта́кты — молекулярные комплексы, обеспечивающие соединения между смежными клетками или между клеткой и внеклеточным матриксом (ВКМ). Межклеточные контакты критически важны для жизнеспособности многоклеточных организмов . Среди контактов, опосредующих соединение двух клеток, выделяют плотные контакты , которые регулируют межклеточный транспорт и предотвращают диффузию мембранных белков ; адгезивные контакты , которые связывают актиновый цитоскелет примыкающих друг к другу клеток; десмосомы , которые связывают промежуточные филаменты соседних клеток; щелевые контакты , обеспечивающие прямой перенос ионов и небольших молекул между соседними клетками. У беспозвоночных животных имеются септированные контакты , которые обладают такими же функциями, что и плотные контакты. У растений многие клетки соединены цитоплазматическими мостиками — плазмодесмами . К числу контактов, связывающих клетки и ВКМ, относят десмосомы и фокальные контакты .

Плотные контакты связывают клетки эпителия или эндотелия . Они регулируют транспорт частиц между клетками, а также выполняют роль «загородки», которая предотвращает диффузию мембранных белков между верхней (апикальной) и нижней (базальной) областями мембраны. В зоне плотных контактов обнаружено более 24 различных белков, которые подразделяют на 4 группы: трансмембранные , полярные, белки цитоскелета и сигнальные белки. Трансмембранные белки представлены тремя типами белков: клаудины , окклюдины и контактные адгезивные молекулы ( англ. junctional adhesion molecule ), которые выполняют «запирающую» функцию. Некоторые белки плотных контактов, например, , взаимодействуют со многими сигнальными белками, а также актиновым цитоскелетом .

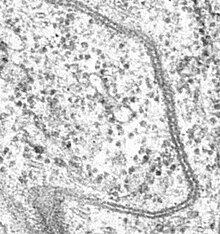
Септированные контакты обнаружены только у беспозвоночных. В электронный микроскоп септированные контакты выглядят как серия (стопка) прямых или изогнутых стенок (или септ, за что они и получили своё название), располагающихся параллельно друг другу. Септы соединяют промежуток шириной 15—20 нм между плазматическими мембранами соседних клеток. Иногда контакт имеет складчатый вид. Со стороны цитоплазмы с септированным контактом могут быть связаны актиновые филаменты .
Септированные и плотные контакты отличаются по белковому составу. Кроме того, плотные контакты находятся на латеральной мембране выше опоясывающих адгезивных контактов, а септированные контакты — ниже, вблизи основания клетки. Иногда одна клетка связана со своими соседями и плотными, и септированными контактами .
Как и плотные контакты, септированные контакты регулируют транспорт молекул через клеточный слой, ограничивая возможность диффузии, а также ограничивают поток фосфолипидов и мембранных белков между верхней ( апикальной ) и нижней ( базальной ) половинами клеточной мембраны. Однако у септированных контактов есть и функции, не свойственные плотным контактам. Например, у дрозофилы и нематоды Caenorhabditis elegans в отсутствие функциональных септированных контактов начинают развиваться опухоли, что указывает на роль этих структур в сдерживании опухолевого роста. Кроме того, септированные контакты играют важную роль в контроле формы клеток .

Чаще всего адгезивные контакты встречаются в эпителиальных тканях и в эндотелии, здесь они образуют вокруг каждой клетки адгезивный поясок, который также называют зоной прилипания ( лат. zonula adherens ). Такие зоны в эпителии позвоночных животных преимущественно размещаются базальнее участка плотных контактов ( лат. zonula occludens ) и апикальнее десмосом ( лат. macula adherens ) . В электронный микроскоп адгезивные контакты выглядят как тёмные плотные полосы, располагающиеся в примыкающих областях мембран смежных клеток. Адгезивные контакты содержат трансмембранные белковые рецепторы — кадгерины . Кадгерины, расположенные на латеральной мембране одной клетки, взаимодействуют с такими же белками на латеральной мембране соседней клетки. Адгезивные контакты достаточно сильны и прочны, чтобы изменять форму ткани и противостоять силам сдвига . Например, в адгезивном пояске кадгерины взаимодействуют с актиновыми филаментами при участии белков группы катенинов . Актиновые филаменты присоединены к миозинам , которые обеспечивают возможность скольжения нитей актина. За счёт скольжения нити происходит изменение формы апикального полюса эпителиальных клеток. Это особенно важно для правильного развития нервной трубки .

Щелевые контакты могут содержать от нескольких десятков до многих тысяч каналов, проходящих сквозь плазматические мембраны смежных клеток. Каждый канал и состоит из двух половин, которые известны как коннексоны , или полуканалы. Эти половины стыкуются в узкой щели шириной 2—3 нм , разделяющей соседние клетки. Каждый коннексон состоит из шести белковых субъединиц — коннексинов . Известны ещё два семейства белков щелевых контактов. обнаружены только у беспозвоночных животных, однако они не являются гомологами коннексинов. Тем не менее, они формируют межклеточные контакты, которые по строению и по функциям похожи на щелевые контакты позвоночных . Другое семейство представлено паннексинами , которые имеются как у позвоночных, так и у беспозвоночных. По структуре они отличаются и от коннексинов, и от иннексинов. Паннексины обнаруживаются почти исключительно в нейронах и, вероятно, играют важную роль в их функционировании и развитии даже у животных с примитивной нервной системой .
Щелевые контакты служат для перемещения ионов и малых молекул между соседними клетками. Через щелевой контакт могут проходить молекулы массой до 1,2 кДа , а молекулы с массой 2 кДа задерживаются. Клетки могут обмениваться такими молекулами, как сахара , нуклеотиды , вторичные посредники (цАМФ или цГМФ ), небольшие пептиды и РНК . Щелевые контакты особенно важны, когда большому количеству клеток необходимо выдать быстрый, хорошо скоординированный ответ. Так, щелевые контакты составляют основу очень быстрых электрических синапсов , которые можно найти, например, в нейронах головного мозга и в клетках миокарда ( кардиомиоцитах ) . Щелевые контакты также используются опухолевыми клетками для взаимной поддержки.

Десмосомы связывают клетки эпителиев, миокарда , печени , селезёнки и некоторые клетки нервной системы. В электронный микроскоп десмосома выглядит следующим образом. В области контакта на цитоплазматической стороне мембраны каждой из двух соседних клеток находится электронно-плотная бляшка, от цитоплазматической стороны которой отходит пучок промежуточных филаментов. Ширина щели между двумя соседними клетками составляет около 30 нм .
Десмосомы обеспечивают структурную целостность слоёв клеток за счёт связывания воедино их сетей промежуточных филаментов. Помимо белков промежуточных филаментов, в состав десмосом входит множество других белков с разнообразными функциями. Десмосомы особенно характерны для клеток, подвергающихся физическим нагрузкам, таким как клетки кожи и миокарда, где они играют важную структурную роль, обеспечивая «точечную сварку» клеток. Десмосомы также задействованы в передаче сигнала. Например, и , входящие в состав десмосом, при активации сигнальных рецепторов на поверхности клетки перемещаются в ядро , где регулируют экспрессию многих генов , а плакоглобин, кроме того, непосредственно связывается с . Таким образом, десмосомы могут контролировать экспрессию многих генов .

Плазмодесмы образуются в ходе последнего этапа деления растительных клеток — цитокинеза , когда дочерние клетки окончательно отделяются друг от друга. В новообразованных клеточных стенках остаются поры, соединяющие цитоплазмы соседних клеток. Плазмодесмы формируются, если при формировании клеточной пластинки в неё попадают трубочки эндоплазматического ретикулума (ЭПР). При этом мембраны соседних клеток и трубочки ЭПР не сливаются друг с другом. Вместо этого трубочка ЭПР оказывается разделённой на две части, которые находятся в соседних клетках и соединены узким перешейком. Перешеек же залегает в канале в клеточных стенках соседних клеток и окружён цитоплазмой; его также называют десмотубулой . Плазмодесмы, сформированные при цитокинезе, называются первичными . Однако иногда плазмодесмы соединяют клетки, не являющиеся продуктом одного деления; такие плазмодесмы называются вторичными . Вторичные плазмодесмы образуются при локальном утоньшении стенки между двумя клетками, и в образующееся отверстие каким-то образом вставляется трубочка ЭПР .
Плазмодесмы объединяют растительные клетки в одну большую структуру, которая называется симпласт . В пределах симпласта через плазмодесмы проходят электрические сигналы, ионы и небольшие водорастворимые молекулы, включая аминокислоты , сахара, мРНК , малые интерферирующие РНК и белки (в том числе транскрипционные факторы ). Ионы и молекулы проходят через плазмодесмы с помощью простой диффузии . Диаметр пор плазмодесм может изменяться, обеспечивая возможность настройки избирательной проницаемости плазмодесм .
Многие патогены растений — вироиды и вирусы — переносятся между клетками посредством плазмодесм, несмотря на свои большие размеры. Дело в том, что вирусы экспрессируют белки, которые расширяют поры плазмодесм, делая возможным транспорт даже довольно крупных вирусных частиц. Механизм действия этих белков пока не известен .
Полудесмосомы располагаются на базальной стороне мембраны эпителиальной клетки и связывают её с внеклеточным матриксом. Точнее, полудесмосомы связывают сеть промежуточных филаментов эпителиальных клеток с внеклеточным матриксом при помощи . Электронная микроскопия показала, что структуры десмосом и полудесмосом очень похожи (полудесмосома выглядит как половина десмосомы, за что эта структура и получила своё название), однако десмосомы связывают не клетку и внеклеточный матрикс, а две соседние клетки. Основная функция полудесмосом заключается в прикреплении слоёв эпителия к базальной мембране .
Полудесмосомы обеспечивают прикрепление многослойного эпителия к базальной мембране. Полудесмосомы и десмосомы в клетке ориентированы под прямым углом друг к другу, благодаря чему они обеспечивают защиту от разных видов механического стресса. Связанные с промежуточными филаментами, полудесмосомы и десмосомы образуют плотную сеть, обеспечивающую структурную поддержку слоёв эпителия .
Несмотря на важную архитектурную роль, десмосомы и полудесмосомы не являются статическими структурами. Так, при ранении кожи клетки эпителия отсоединяются от базальной мембраны и мигрируют в область раны. Там они делятся, восстанавливая популяцию клеток в зоне ранения, и затем снова прикрепляются к базальной мембране (с помощью полудесмосом) и друг к другу (с помощью десмосом). Таким образом, полудесмосомы и десмосомы способны к обратимой разборке .

Фокальные контакты представляют собой скопление интегриновых рецепторов на клеточной мембране, которые связывают клетку с внеклеточным матриксом; со стороны цитоплазмы они взаимодействуют с актиновым цитоскелетом. Помимо интегринов, в состав фокальных контактов входят белки , , киназа фокальных контактов, , , , α-актинин и другие. Фокальные контакты выявляются только в тех участках клеточной мембраны, которые приближаются к внеклеточному матриксу на расстояние менее 15 нм .
Фокальные контакты обеспечивают сильное прикрепление клеток к внеклеточному матриксу и участвуют в передаче механического напряжения на мембране клетки. Они задействованы во многих сигнальных путях клетки, в частности, активирующихся в ответ на механический стресс .
Нарушения в работе межклеточных контактов разных типов приводят к разнообразным заболеваниям, чем подчёркивается их функциональная важность для многоклеточного организма. Например, мутации в генах клаудина-16 и клаудина-19, нарушающие работу плотных контактов, приводят к вследствие чрезмерной потери магния с мочой. Мутации в генах клаудина-13 и трицеллюлина вызывают наследственную глухоту . Дисрегуляция некоторых белков плотных контактов связана с онкологическими заболеваниями (например, экспрессия ZO-1 и ZO-2 снижается во многих типах рака). Компоненты плотных контактов также могут быть мишенями для онкогенных вирусов .
Точечные мутации в генах, кодирующих белки щелевых контактов — коннексины, у человека приводят к очень специфическим дефектам, из чего можно заключить, что большая часть коннексинов экспрессируется лишь в нескольких тканях . Рецессивные мутации в гене являются наиболее частой причиной наследственной глухоты. Коннексин-26 участвует в транспорте ионов калия в клетках эпителия, поддерживающего чувствительные волосковые клетки в ухе . Люди с мутациями в гене, кодирующем , могут страдать от разрушения миелиновой оболочки аксонов ( X -связанный вариант болезни Шарко — Мари — Тута ). Возможно, стабильность миелина может зависеть от щелевых контактов между клетками разных слоёв миелиновой оболочки, и нарушения в функционировании контактов приводят к её разрушению .
Нарушение функционирования десмосом также приводит к различным заболеваниям. Так, развивается при мутациях в белках десмосом. Аутоиммунные буллёзные дерматозы — такие, как , наблюдаются, когда у пациентов образуются антитела к собственным белкам десмосом. Нарушения в функционировании межклеточных контактов, в частности, десмосом, могут приводит к летальному исходу . Заболевания, связанные с дисфункцией полудесмосом, проявляются в образовании пузырей на коже . Такие заболевания известны под общим названием буллёзный эпидермолиз (БЭ). При БЭ кожа становится хрупкой, отслаивается и покрывается волдырями в ответ на малейший механический стресс. Иногда также происходит отслоение эпителия роговицы , трахеи , пищеварительного тракта , пищевода , а также имеет место мышечная дистрофия .
|
|
|||||
|---|---|---|---|---|---|
|
|||||
| Запирающие контакты |
|
||||
| Контакты, формирующие каналы |
|
||||
|
Структуры
клеточной мембраны
|
|
|---|---|
| Мембранные белки | |
| Другое | |