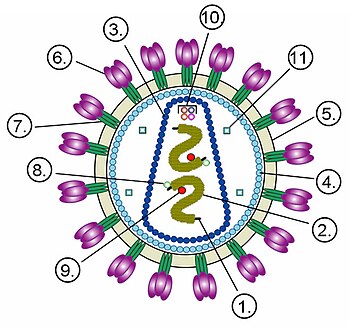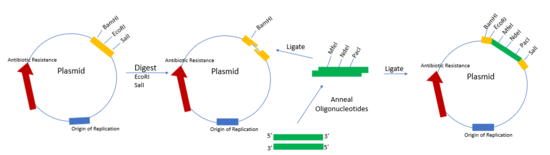
Плазмиды
- 1 year ago
- 0
- 0

Транспозоны ( англ. transposable element, transposon) — участки ДНК организмов, способные к передвижению (транспозиции) и размножению в пределах генома . Транспозоны также известны под названием «прыгающие гены» и являются примерами мобильных генетических элементов .
Транспозоны формально относятся к так называемой некодирующей части генома — той, которая в последовательности пар оснований ДНК не несёт информацию об аминокислотных последовательностях белков, хотя некоторые классы мобильных элементов содержат в своей последовательности информацию о ферментах , транскрибируются и катализируют передвижения; например, ДНК-транспозоны и кодируют белки транспозаза , и . У разных видов транспозоны распространены в разной степени: так, у человека транспозоны составляют до 45 % всей последовательности ДНК, у плодовой мухи Drosophila melanogaster часть мобильных элементов составляет лишь 15—20 % всего генома . У растений транспозоны могут занимать основную часть генома — так, у кукурузы ( Zea mays ) с размером генома в 2,3 миллиарда пар оснований по крайней мере 85 % составляют различные мобильные элементы .
Барбара Макклинток исследовала вариации окраски зерна и листьев кукурузы, и в 1948 году путём цитологических и генетических исследований пришла к выводу, что мобильные участки ДНК, -элементы, приводят к соматическому мозаицизму растений . Она была первой, кто доказал, что геном эукариот не статичен, а содержит участки, которые могут передвигаться. В 1983 году за эту работу Барбара Макклинток получила Нобелевскую премию .
Хотя транспозоны были открыты в 1940-х годах, только через полвека стало понятно, насколько велика их доля в геноме организмов. Так, получение первой нуклеотидной последовательности ( секвенирование ) генома человека показало, что мобильных элементов в последовательности ДНК не менее 50 %. Точную оценку получить трудно, поскольку некоторые транспозонные участки со временем настолько изменились, что их нельзя уверенно идентифицировать .
Поскольку транспозоны потенциально способны вызывать вредные мутации и поломки хроматина , с начала открытия мобильных элементов считалось, что их действие сводится к геномному паразитизму. Но в начале XXI столетия появляется всё больше данных о возможных благоприятных эффектах транспозонов для организмов , об эволюционном влиянии ретротранспозонов на геном плацентарных млекопитающих . Идентифицируют случаи использования транспозонов организмами. Например, РНК ретротранспозона участвует в образовании гетерохроматина во время инактивации X-хромосомы . Плодовая муха не имеет теломеразы , а вместо этого использует обратную транскриптазу ретротранспозонов для продления теломерных участков, которые у Drosophila melanogaster представлены повторами транспозонов .

Мобильные генетические элементы относятся к повторяющимся элементам генома — тем, которые имеют несколько копий в последовательности ДНК клетки . Повторяющиеся элементы генома могут располагаться в тандеме ( микросателлиты , теломеры и т. д.) и могут быть рассеяны по геному (мобильные элементы, псевдогены и т. д.) .
Мобильные генетические элементы по типу транспозиции можно разделить на два класса: ДНК-транспозоны , которые применяют метод «вырезать и вставить», и ретротранспозоны , передвижение которых имеет в своем алгоритме синтез РНК из ДНК с последующим обратным синтезом ДНК из молекулы РНК, то есть метод «копировать и вставить».
Транспозоны также можно разделить по степени автономности. Как ДНК-транспозоны, так и ретротранспозоны имеют автономные и неавтономные элементы. Неавтономные элементы для транспозиции нуждаются в ферментах, которые кодируются автономными элементами, которые часто содержат значительно изменённые участки транспозонов и дополнительные последовательности. Количество неавтономных транспозонов в геноме может значительно превышать количество автономных .
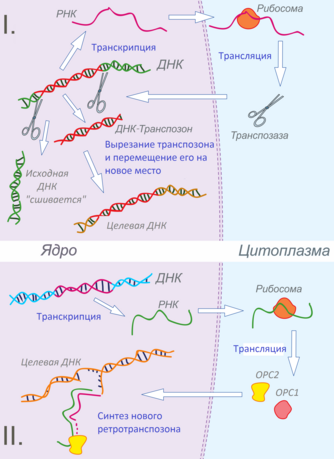
ДНК-транспозоны передвигаются по геному способом «вырезать и вставить» благодаря комплексу ферментов под названием транспозаза . Информация об аминокислотной последовательности белка транспозазы закодирована в последовательности транспозона. Кроме того, этот участок ДНК может содержать другие, связанные с транспозоном последовательности, например гены или их части. Большинство ДНК-транспозонов имеют неполную последовательность. Такие транспозоны не являются автономными и передвигаются по геному благодаря транспозазе, которая закодирована другим, полным, ДНК-транспозоном .
На концах участков ДНК-транспозона расположены инвертированные повторы, которые являются особыми участками узнавания транспозазы, таким образом отличая эту часть генома от остальных. Транспозаза способна делать двухцепочные разрезы ДНК, вырезать и вставлять в ДНК-мишень транспозон .
К ДНК-транспозонам принадлежат -элементы растений, которые были впервые открыты Барбарой Макклинток в кукурузе. Ac -элемент ( англ. Activator) является автономным и кодирует транспозазу. Есть несколько типов Ds -элементов, которые способны к формированию разрывов хромосом и которые перемещаются по геному благодаря Ac -элементам .
Гелитроны ( англ. Helitron) — тип транспозонов, который есть у растений , животных и грибов , но который широко представлен в геноме кукурузы, где он, в отличие от других организмов, находится в частях ДНК, богатых генами . Гелитроны транспозируются по механизму «катящегося кольца» ( англ. rolling circle). Процесс начинается с разрыва одной цепи ДНК-транспозоны. Высвобожденный участок ДНК вторгается в последовательность-мишень, где формируется гетеродуплекс . С помощью ДНК-репликации завершается внедрение транспозона в новый участок .
Гелитроны могут захватывать соседние последовательности при транспозиции.
Ретротранспозоны — это мобильные генетические элементы, которые применяют метод «копировать и вставить» для распространения в геноме животных . По крайней мере 45 % генома человека составляют ретротранспозоны и их производные. Процесс передвижения включает промежуточную стадию молекулы РНК , которая считывается с участка ретротранспозона и которая затем, в свою очередь, используется как матрица для обратной транскрипции в последовательность ДНК. Новосинтезированный ретротранспозон встраивается в другой участок генома.
Активные ретротранспозоны млекопитающих делятся на три основные семьи: Alu-повторы, ДДП-1, SVA.



Мобильные элементы генома достаточно широко представлены в растительных и животных геномах. Их высокая активность является риском для стабильности генома , поэтому их экспрессия жестко регулируется, особенно в тех тканях , которые принимают участие в формировании гамет и передаче наследственной информации потомкам. У растений и животных регуляция активности мобильных элементов генома происходит путём de novo - метилирования последовательности ДНК и активности некодирующих РНК вместе с белковыми комплексами Аргонавт .
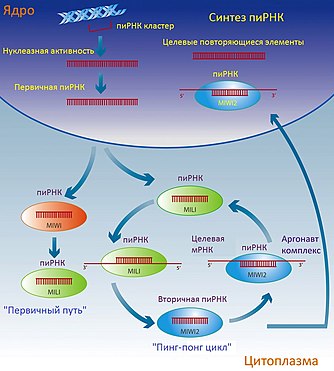
Основная роль малых некодирующих РНК, которые взаимодействуют с пиви-комплексом, или пиРНК , заключается в подавлении мобильных элементов генома в зародышевых тканях. Эта роль пиРНК достаточно высоко консервативна у животных .
У мышей мобильные элементы генома на протяжении онтогенеза находятся преимущественно в неактивном состоянии, которое достигается путём эпигенетических взаимодействий и активности некодирующих РНК . В период эмбрионального развития эпигенетическая метка метилирования ДНК подвергается репрограммированию: родительские метки стираются, а новые устанавливаются . В этот период часть белков-аргонавтов — пиви-белки (Mili и Miwi2) — и некодирующие РНК, которые с ними взаимодействуют — пиРНК — играют ключевую роль в de novo подавлении ретротранспозонов мышей путём метилирования ДНК, и пинг-понг-цикла амплификации пиРНК, и подавления мишени . Если у мышей возникает недостаток белков Mili и Miwi2, это приводит к активации и и остановке гаметогенеза и стерильности у самцов . Недавние работы показали, что у мухи Drosophila melanogaster активным кофактором в подавлении является белок .
Механизм пиРНК -индуцированного подавления транспозонов окончательно не выяснен, но схематически его можно представить такой моделью :
В отличие от вирусов , которые используют организм хозяина для размножения и способны его покинуть, мобильные генетические элементы существуют исключительно в организме хозяина. До некоторой степени поэтому транспозоны способны регулировать свою активность. Примером этого является Ac - ДНК-транспозоны — автономные мобильные элементы растений, кодирующие собственную транспозазу. Ac -элементы проявляют способность снижать активность транспозазы при увеличении её копий .
Также подавление растительных автономных ДНК-транспозонов MuDR может происходить с помощью Muk. Muk является вариантом MuDR и имеет в своей последовательности несколько палиндромных участков ДНК. Когда Muk транскрибируется, такая РНК формирует шпильку, затем режется комплексом ферментов на малые интерферирующие РНК (миРНК), которые заглушают активность MuDR с помощью процесса РНК-интерференции .
По состоянию на 2012 год задокументировано 96 различных заболеваний человека, причиной которых является de novo внедрение мобильных генетических элементов . Alu-повторы часто вызывают хромосомные аберрации и являются причиной 50 разновидностей заболеваний . Так, у нейрофиброматоза I типа было найдено 18 случаев встроенных ретротранспозонов , 6 из которых происходят в 3 специфических местах. Активность мобильных элементов в соматических тканях зафиксирована у пациентов с раком легких .
Если транспозиция, которая вызывает заболевания, происходит в гаметах , то следующие поколения наследуют болезни. Так, гемофилия может возникать из-за встраивания ретротранспозона ДДП-1 в участок ДНК, кодирующий ген VIII фактора свертывания крови . У мышей были зафиксированы случаи онкогенеза, остановки развития и стерильность в связи со встраиванием мобильных элементов генома .
Некоторые этапы эволюционирования организмов были вызваны активностью мобильных элементов генома. Уже первая нуклеотидная последовательность генома человека доказала, что многие гены были производными транспозонов . Мобильные элементы генома могут влиять на организацию генома путём рекомбинации генетических последовательностей и входя в состав таких фундаментальных структурных элементов хроматина , как центромеры и теломеры . Мобильные элементы могут влиять на соседние гены, меняя узоры ( паттерны ) сплайсинга и полиаденилирования или выполняя функции энхансеров или промоторов . Транспозоны могут влиять на структуру и функции генов путём выключения и изменения функций, изменения структуры генов, мобилизации и реорганизации фрагментов генов и изменения эпигенетического контроля генов .
Репликация транспозонов может вызвать некоторые заболевания, но, несмотря на это, в процессе эволюции транспозоны не были удалены и остались в ДНК -последовательностях почти всех организмов, или в виде целых копий, которые имели возможность передвигаться по ДНК, или в укороченном виде, потеряв способность к передвижению. Но укороченные копии также могут принимать участие в таких процессах, как пост-транскрипционная регуляция генов, рекомбинация и т. п. Также важным моментом в потенциальной способности транспозонов влиять на темпы эволюции является то, что их регуляция зависит от эпигенетических факторов. Это приводит к возможности транспозонов реагировать на изменения окружающей среды и вызывать генетическую нестабильность . На стресс транспозоны активируются или прямо, или путём снижения их подавления белками-аргонавтами и пиРНК . У растений мобильные генетические элементы очень чувствительны к различным типам стресса, на их активность могут влиять многочисленные абиотические и биотические факторы , среди которых солёность , ранения, холод, тепло, бактериальные и вирусные инфекции .
Ещё одним возможным механизмом эволюции геномов организмов является горизонтальный перенос генов — процесс передачи генов между организмами, которые не находятся в отношениях «предки-потомки». Есть сведения о том, что взаимодействия паразитических организмов и животных-хозяев могут повлечь горизонтальный перенос генов с помощью транспозонов, который состоялся между позвоночными и беспозвоночными организмами .
Считается, что приобретённый иммунитет млекопитающих возник у челюстных рыб примерно 500 миллионов лет назад . Приобретённый иммунитет позволяет формировать антитела для многих видов патогенов , попадающих в организм млекопитающих, включая человека. Для формирования различных антител клетки иммунной системы изменяют последовательность ДНК путём соматической рекомбинации с помощью системы, которая возникла и эволюционировала благодаря мобильным элементам генома .
Нейроны , клетки нервной системы, могут иметь , то есть последовательность ДНК у них отличается от последовательности ДНК других клеток, хотя все они сформировались из одной клетки-предшественника — зиготы . Доказано, что у крыс специально вставленные -ретротранспозоны человека активны даже в зрелом возрасте. Также зафиксировано увеличение копий ДДП-1-ретротранспозонов в нейронах некоторых участков мозга , в частности гипоталамуса , по сравнению с другими тканями у взрослых людей . Также установлено, что мобильные элементы приводят к разнородности в нейронах мухи Drosophila melanogaster . Активность мобильных элементов в нейронах может повлечь синаптическую пластичность и большую вариабельность поведенческих реакций .
Последовательности ДНК генов теломеразы и ДДП-1-ретротранспозонов имеют высокую гомологию, что свидетельствует о возможности происхождения теломераз от ретротранспозонов .
У растений очень большая скорость эволюции геномов, поэтому лучше всего известны те влияния мобильных элементов, которые возникли вследствие одомашнивания , поскольку оно произошло недавно, и эти изменения легко идентифицировать, поскольку известны черты, по которым велась селекция культурных растений . Примером может быть приобретение овальной формы римским помидором Solanum lycopersicum . Ген, который находится в локусе SUN, был перемещён путём ретротранспозиции в другой участок ДНК, где он регулируется другими промоторными последовательностями у овальных томатов .
Поскольку мобильные элементы генома способны к встраиванию в хроматин , они используются в генной инженерии для специального и контролируемого встраивания генов или участков ДНК, которые изучают учёные. Транспозоны используются для мутагенеза и для определения регуляторных элементов генома в лабораториях.
Наиболее известная система для введённого мутагенеза in vivo — P-мобильный элемент мухи D. melanogaster , с помощью которого можно изучать функции генов, налаживание хромосомных аберраций и т. п.
У позвоночных животных долгое время не было эффективной методики транспозонной модификации генома. Сейчас есть система мобильного элемента Tol2, полученная из японской рыбы Oryzias latipes , которая используется как у мышей, так и на клеточных линиях человека . Также успешной является система транспозонов Minos .
Система транспозонов «Спящая Красавица» ( англ. Sleeping Beauty) была создана на основе последовательности ДНК транспозазы из рыбы. Удачное использовании этой системы на мышах позволило определить кандидатов в онкогены рака кишечника человека .
Кроме использования транспозонов в генной инженерии, изучение активности транспозонов является методом филогенетики . Путём анализа и сопоставления нуклеотидных последовательностей геномов различных видов можно найти транспозоны, которые имеются у одних видов, но отсутствуют у других. Виды, у которых есть одинаковый ретротранспозон, скорее всего получили его от общего предка. Таким образом, можно получить информацию об эволюционном развитии видов и строить филогенетические деревья .