Новые муравьи в штанах
- 1 year ago
- 0
- 0


Муравьи́ ( лат. Formicidae ) — семейство насекомых из надсемейства муравьиных , отряда перепончатокрылых . Являются общественными насекомыми , образующими 3 касты : самки, самцы и рабочие особи. Самки и самцы крылатые , рабочие особи — бескрылые. Усики коленчатые, у самок и рабочих особей 11—12-члениковые, у самцов 12—13-члениковые , у ряда видов 4-, 6- или 10-члениковые. Основной членик усика (скапус) обычно намного длиннее всех остальных. Задний отдел груди (эпинотум) представляет собой первый сегмент брюшка, слившийся с заднегрудью. Собственно брюшко присоединяется к эпинотуму стебельком, образованным первым или вторым сегментами. У муравьёв некоторых подсемейств ( мирмицины , понерины и других) имеется развитое жало. Крылья с редуцированным жилкованием .
Муравьи живут семьями в гнёздах , называемых муравейниками , которые устраивают в почве, древесине, под камнями; некоторые сооружают муравейники из мелких растительных частиц и т. п. Существуют паразитические виды , которые обитают в гнёздах других муравьёв, муравьи-«рабовладельцы», содержащие в своих гнёздах «рабов» — муравьёв других видов. Ряд видов приспособился к обитанию в жилищах человека. Некоторые виды ценятся за регулирование численности насекомых-вредителей , другие могут считаться вредителями.
Питаются преимущественно соком растений, падью тлей и других сосущих насекомых, в период кормления личинок — преимущественно насекомыми. Есть также виды, питающиеся семенами ( муравьи-жнецы ) и культивируемыми грибами ( муравьи-листорезы ).
Распространены по всему миру, за исключением Антарктиды и некоторых удалённых островов, образуя 10—25 % земной биомассы наземных животных . Общая численность всех муравьев на Земле составляет примерно 20 квадриллионов особей . Успех муравьёв во многих средах обитания обусловлен их социальной организацией и способностью изменять место обитания и использовать разнообразные ресурсы.
Одним из первых исследователей, который в своих научных работах описал общественную жизнь муравьёв, был энтомолог Эрих Васманн , который также является одним из основоположников мирмекологии — науки, изучающей муравьёв.
В мире известно более 14 000 современных видов и 345 родов, а также ископаемых 166 родов и более 760 видов муравьёв, распространённых преимущественно в тропиках . В Палеарктике около 1350 видов из 94 родов, в России отмечено более 260 видов из 44 родов .
Русское слово муравей восходит к праслав. *morvь(jь) , которое продолжает пра-и.е. *morwis/*mormis (к тому же источнику восходят лат. formica и др.-греч. μύρμηξ ). При этом фонетически закономерной формой в русском было бы моровей . Современная форма получилась из контаминации со словом мурава .

Муравьи — наиболее эволюционно продвинутое семейство насекомых с точки зрения этологии , экологии и физиологии . Их семьи представляют собой сложные социальные группы с разделением труда и развитыми системами коммуникации и самоорганизации , позволяющими особям координировать свои действия при выполнении задач, которые не по силам одному индивиду. Некоторые виды муравьёв обладают развитым «языком» и способны передавать сложную информацию. Кроме того, многие виды муравьёв поддерживают высокоразвитые симбиотические отношения с другими насекомыми, грибами, бактериями и растениями.
Преимущества, предоставляемые муравьям кооперацией, привели к тому, что на сегодняшний день они являются доминирующей по численности группой членистоногих. Так, на 1 км² саванны в Кот-д’Ивуаре ( Африка ) обитает почти 2 млрд особей муравьёв, образующих примерно 740 тысяч колоний . Вместе с термитами (ещё одной крупной группой общественных насекомых ) муравьи составляют треть общей биомассы наземных животных в тропическом лесу бассейна Амазонки . Таким образом, при средней популяционной плотности 800 млн муравьёв и 100 млн термитов на 1 км² они в сумме весят лишь вдвое меньше, чем все прочие сухопутные животные этого дождевого леса . В областях с умеренным климатом их меньше. На площади 8 км² во Флориде обнаружено 76 видов муравьёв из 30 родов, а на площади 5,5 км² в Мичигане — 87 видов из 23 родов .
Размеры представителей семейства разнообразны — от 1 до 30—50 мм и более. К одним из самых маленьких муравьёв относятся представители родов мономориум (рабочие особи 1—2 мм , самки и самцы 3—4 мм ) , Mycetophylax (1—3 мм) и Cyphomyrmex (1,5—3 мм) и вид Leptothorax minutissimus — эндемик США , паразитирующий в колониях муравьёв и достигающий длины до 3 мм .
Одним из крупнейших представителей семейства является вид Camponotus gigas , размер рабочих особей которого составляет около 20 мм, самцов — 18,3 мм, солдат — 28,1 мм, матки — до 31,3 мм . Также самыми крупными муравьями являются динопонера гигантская ( Dinoponera gigantea ) и Paraponera clavata , достигающие длины 25—30 мм . Самцы из африканского рода Dorylus могут достигать длины до 3 см, а матки (королевы) в оседлую фазу в момент созревания яиц имеют сильно увеличенное физогастрическое брюшко и общую длину до 5 см . Однако крупнейшими в истории являются ископаемые муравьи рода Formicium . Их самки достигали 7 см в длину, а крылья имели размах до 15 см .
|
|
Семейство муравьёв относится к отряду перепончатокрылых , который также включает пилильщиков , пчёл и ос , и ранее считалось, что принадлежит к надсемейству Vespoidea . По другим взглядам выделяется в отдельный таксон Formicoidea . По данным прежнего филогенетического анализа считалось, что муравьи произошли от каких-то веспоидных ос в середине мелового периода , примерно 110—130 млн лет назад, вероятно, на территории бывшего суперконтинента Лавразии и в качестве ближайших к муравьям семействам рассматривались настоящие осы и сколии ( Scoliidae и Bradynobaenidae ) . По другим более современным филогеномным данным, основанным на исследовании нескольких сотен генов, веспоидные осы не являются родственниками муравьёв, а последние скорее сближаются со сфекоидными роющими осами ( Sphecoidea , Apoidea ) . Подтверждением происхождения муравьёв от ос , помимо анатомического и поведенческого сходства, является обнаружение в 1967 году в мезозойских отложениях переходной формы между ними — ископаемого вида Sphecomyrma freyi . Данный вид сочетает в себе как признаки муравьёв, так и признаки ос, и датируется концом мелового периода (80 млн лет назад). Впоследствии были обнаружены и другие виды, которые также были отнесены к подсемейству Sphecomyrminae .

Вероятно, Sphecomyrma freyi были наземными фуражирами , но, основываясь на строении и поведении современных представителей подсемейств Leptanillinae и Martialinae , некоторые учёные полагают, что примитивные муравьи были подземными хищниками . После становления доминирующей роли цветковых растений , около 100 млн лет назад, муравьи начали эволюционировать быстрее, адаптируясь к различным экологическим нишам .
В меловом периоде численность муравьёв была невысокой и составляла около 1 % от общей численности насекомых . Муравьи стали доминировать после адаптивной радиации в начале третичного периода . Из всех насекомых, найденных в датированных олигоценом и миоценом ископаемых, от 20 до 40 % составляли муравьи. Из всех родов, живших в эпоху эоцена , примерно 10 % дожили до наших дней. Роды, существующие сегодня, составляют 56 % родов, найденных в балтийском янтаре (датируется началом олигоцена) и 92 % родов, найденных в доминиканском янтаре (начало миоцена) .
| Докембрий | Фанерозой | Эон | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Палеозой | Мезозой | Кайнозой | Эра | ||||||||||
| Кембрий |
Ордо
вик |
Сил
ур |
Девон | Карбон | Пермь | Триас | Юра | Мел |
Палео
ген |
Нео
ген |
П-д | ||
| 4570 | 541 | 485,4 | 443,4 | 419,2 | 358,9 | 298,9 | 252,2 | 201,3 | 145,0 | 66,0 | 23,03 |
млн
лет ← |
|
| 2,588 | |||||||||||||
Термиты , которых иногда и называют «белыми муравьями», муравьями не являются. Они принадлежат к инфраотряду Термиты ( Isoptera ) и находятся в более тесном родстве с тараканами и богомолами . Так же, как и муравьи, термиты — это общественные насекомые , но они имеют неполное превращение . Сходство социальных структур термитов и муравьёв объясняется конвергентной эволюцией . Так называемые бархатные муравьи (Mutillidae), хоть и похожи на муравьёв, являются бескрылыми осами .
Семейство муравьёв включает по разным оценкам от 5 до 6 вымерших подсемейств ( Armaniinae , Formiciinae , Haidomyrmecinae , Sphecomyrminae и другие), 5 вымерших триб , 121 вымерший род , около 600 вымерших видов . Первый мезозойский муравей был обнаружен в 1967 году , когда Эдвард Уилсон с коллегами описал ископаемые останки мелового муравья ( Sphecomyrma freyi ) в куске янтаря с пляжа в Нью-Джерси ( США ). Возраст находки составляет около 90 млн лет. Палеонтологическая история муравьёв в последнее время интенсивно исследовалась , однако многие ископаемые остатки плохо сохранились, и описание их весьма затруднительно.
|
Обнаруженный в 2008 году в Бразилии примитивный слепой вид муравьёв Martialis heureka был признан обладающим уникальными признаками строения и выделен в отдельное новое подсемейство муравьёв Martialinae . Другим примером живого ископаемого является «динозавровый муравей» Nothomyrmecia macrops из Австралии. Найденный в 1931 году , он был описан в 1934 году, а повторно , несмотря на многочисленные попытки и экспедиции, его нашли только в 1977 году . В своё время его даже выделяли в самостоятельное подсемейство Nothomyrmeciinae .
Семейство муравьёв включает 16 современных и 6 ископаемых подсемейств (22 с учётом Armaniidae в статусе Armaniinae ), 44 трибы , более 14 000 современных валидных видов и 345 родов (а также ископаемых 166 родов и более 760 видов), распространённых преимущественно в тропиках .
Трудность классификации муравьёв связана с двумя феноменами — наличием видов-двойников и гибридов . Существует довольно много видов, практически не различимых по внешнему виду. Таким образом, вид, описанный по анатомическим признакам небольшого числа особей, нередко разделяют на два или более самостоятельных, изолированных друг от друга репродуктивно. Отличить их друг от друга можно по генетическим или ферментным особенностям. И напротив, два близкородственных вида муравьёв, которые легко различимы по внешним признакам, в местах совместного обитания нередко скрещиваются и дают гибридные формы. При плодовитости таких гибридов делается вывод о том, что виды не самостоятельные, а являются лишь различными расами одного вида, так как потомство от скрещивания разных видов не плодовито .
Филогенетические взаимоотношения подсемейств муравьёв показаны ниже по данным Brady et al. (2006), Moreau et al. (2006) , Ward (2007) , Rabeling et al. (2008) . Недавно, в работе Kück et al. (2011) было показано, что позиция подсемейств Martialinae и Leptanillinae должна быть пересмотрена, так как они близки к кладе, включающей Formicinae .
В 2014 году на основе молекулярно-филогенетического исследования кочевых и полукочевых муравьёв было предложено (Brady et al., 2014) снова объединить все дориломорфные подсемейства ( Aenictinae , Aenictogitoninae , Cerapachyinae , Ecitoninae и Leptanilloidinae ) в составе расширенного Dorylinae S.l. , принимаемого в широком объёме (вместо одного рода Dorylus там теперь 18 родов и 797 видов) .


Муравьи отличаются по своей морфологии от других насекомых наличием коленчатых усиков , метаплевральной железой , а также сильным сужением второго абдоминального сегмента в узел-петиоль . Голова, мезосома и брюшко являются тремя различными сегментами тела. Петиоль — это узкая талия между мезосомой (три грудных сегмента, а также первый абдоминальный сегмент, который сливается с ними) и брюшком (абдоминальные сегменты после петиоля). Он может состоять из одного или двух члеников (только второй или второй и третий абдоминальные сегменты) . Брюшко и петиоль вместе объединяют в метасому .
Как и другие насекомые, муравьи имеют экзоскелет — наружную хитиновую оболочку, которая обеспечивает поддержку и защиту тела. Нервная система состоит из брюшной нервной цепочки, которая расположена по всей длине тела, и нескольких нервных узлов , связанных между собой . Наиболее важной частью нервной системы является надглоточный ганглий , в котором образуются временные связи. Его объём относительно наибольший у рабочих, меньше у цариц и самый маленький у самцов.
Как и большинство насекомых, муравьи имеют сложные фасеточные глаза , состоящие из многочисленных крошечных линз. Муравьиные глаза хорошо различают движение, но не обладают высокой разрешающей способностью. Кроме сложного глаза, имеются три простых глазка на верхней части головы, которые определяют уровень освещённости и плоскость поляризации света . По сравнению с позвоночными животными , большинство муравьёв обладают посредственным зрением , а некоторые подземные виды являются полностью слепыми.
Усики на голове являются органами чувств, служащими для обнаружения химических веществ, воздушных потоков и вибраций, а также используются для приёма и передачи сигналов через прикосновения.
Голова муравьёв имеет сильные мандибулы , которые используются для переноса пищи, манипулирования различными объектами, строительства гнезда и обороны . У некоторых муравьёв эти жвалы открываются на 270° и защёлкиваются словно капканы, например, у таких родов, как Anochetus , Odontomachus , Myrmoteras , Strumigenys . У некоторых видов имеется небольшой отросток пищевода, называемый «общественным желудком», или зобом . В нём может храниться пища, которая впоследствии распределяется между другими муравьями и личинками . (См. Трофаллаксис ).

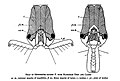


Крючковатый коготь на конце каждой лапки помогает муравью подниматься по вертикальным поверхностям. У большинства цариц и самцов муравьёв есть крылья. Царицы отгрызают крылья после брачного полёта .
В брюшке муравья расположены внутренние органы, включая репродуктивные, а также выделительные железы. У рабочих муравьёв многих видов на конце брюшка присутствует видоизменённый яйцеклад в форме жала , который используется для добывания пищи, а также защиты гнезда или нападения (например, у мирмицин , Myrmeciinae , Paraponerinae , понерин и др.) . У примитивных форм подсемейства мирмицины жало редуцировано, а у эволюционно продвинутых форм — видоизменено, при этом даже у примитивных родов данной группы оно неспособно эффективно парализовать жертву.
Сигнальные вещества выделяются специальными железами. У различных видов муравьёв их может быть десятки. Данные железы различаются по количеству, форме, функциям и никогда не встречаются одновременно у одного вида . Экзокринная система у муравьёв хорошо развита и включает более 75 желёз, которые располагаются во всех частях тела: в голове и усиках (более 15), в груди и ногах (более 30), в брюшке (более 30) . Они выделяют феромоны, участвуют в выработке широкого спектра других веществ, таких как ферменты, компоненты ядов, антибиотики, смазочные материалы и т. д. У муравья Pachycondyla tridentata только в одном брюшке содержится не менее чем 28 желёз .
Метаплевральные железы выделяют антибиотические вещества (например, фенилуксусную кислоту ), а иногда также феромоны тревоги и репелленты для защиты от врагов. Однако они отсутствуют у ряда видов, ведущих древесный образ жизни. Для защиты и нападения также используются половые железы, изменившие своё назначение у рабочих особей. Среди них выделяют ядовитую железу, вырабатывающую кислотный экскрет, и железы Дюфура , вырабатывающие щелочной экскрет, разнообразные углеводороды, кетоны , спирты , сложные эфиры и лактоны . Основу яда многих нежалящих видов составляет муравьиная кислота . Такие муравьи выбрызгивают секрет на расстояние до нескольких сантиметров, одновременно выделяя «феромоны тревоги». Наиболее токсичен яд некоторых жалящих видов, имеющий, как и большинство ядов животных, сложный состав . Основное действующее начало яда огненных муравьёв — алкалоид из группы пиперидинов , соленопсин ; в состав яда входят также несколько аллергенных белков. Наиболее токсичен яд некоторых видов муравьёв-жнецов рода Pogonomyrmex ; LD50 яда P. badius для мышей при внутрибрюшинном введении — 0,42 мкг/г .
Пигидиальные железы открываются сзади протоками на верхней стороне брюшка. У ряда видов они вырабатывают феромоны тревоги и репелленты, других — следовые феромоны. Стернальные железы открываются протоками на нижней стороне брюшка и выделяют следовые и призывные феромоны. Мандибулярные железы открываются протоками на внутренней стороне жвал .
Ранее энтомологами считалось, что все муравьи способны выделять ядовитую муравьиную кислоту (откуда и произошло её название) , однако сейчас известно, что на это способны только представители подсемейства формицины .
Выделение шёлкоподобного вещества личинками муравьёв для создания защитного кокона зафиксировано в подсемействах Aneuretinae , Apomyrminae , Cerapachyinae , Ecitoninae , Formicinae , Myrmeciinae , Nothomyrmeciinae и Ponerinae .

Муравьиная семья — многолетнее, высокоорганизованное сообщество, состоящее из расплода ( яйца , личинки , куколки ), взрослых половых особей ( самок и самцов ) и, как правило, многочисленных рабочих особей (бесплодных самок) .
Муравьи образуют семьи, размеры которых варьируют от нескольких десятков особей до высокоорганизованных колоний , состоящих из миллионов особей и занимающих большие территории. Крупные семьи состоят в основном из бесплодных бескрылых самок, формирующих касты рабочих, солдат или другие специализированные группы. Почти во всех семьях есть самцы и одна или несколько репродуктивных самок, называемых царицами или королевами. Иногда семьи называют суперорганизмом , потому что муравьи работают как единое целое .
В муравьиных семьях существуют разделение труда , связи между особями и самоорганизация при решении сложных проблем. Такие параллели с человеческим обществом давно являются предметом исследований учёных .
Муравьиная семья имеет одну ( моногиния ) или несколько ( полигиния ) репродуктивных самок, в зависимости от вида и размера семьи — называемых королевами или матками . Самки похожи на рабочих, но отличаются от них строением груди и, как правило, более крупными размерами. Имеют крылья, которые откусывают себе после оплодотворения. У некоторых видов есть эргатоидные самки , бескрылые и с упрощённым строением груди .
У большинства видов царицы и рабочие развиваются из оплодотворённых яиц — у них имеется два набора хромосом , полученных от сперматозоида и яйцеклетки .
Самка спаривается только один раз во время «брачного полёта» , получая при этом запас спермы самца, расходуемый постепенно на протяжении всей её жизни. Продолжительность жизни муравьиной самки максимальна для мира насекомых и может составлять до 12—20 лет, в зависимости от вида (рекорд для самок в 28 лет зафиксирован в лабораторном гнезде Lasius niger ) . Оплодотворённые самки сбрасывают крылья и либо основывают новую семью, либо остаются в своём муравейнике . Иногда молодых самок принимают в другие, уже существующие семьи своего вида. В первом случае самка должна выбрать место для гнезда, подготовить первую камеру нового муравейника и приступить через некоторое время к откладыванию яиц.

У некоторых видов самки занимаются сбором пищи, и им приходится покидать гнездо. У других — они остаются в гнезде, поддерживая собственное существование и выращивая первых рабочих за счёт жировых запасов и подвергающихся гистолизу мышц крыльев . Царица кормит личинок специальным слюнным секретом и/или особыми «кормовыми» яйцами. Количество доступного корма сначала весьма ограничено, поэтому между числом и размером первых рабочих ищется компромисс — все они являются мелкими или даже карликовыми .

Самцы (за некоторыми исключениями) рождаются из неоплодотворённых яиц и являются носителями только одного набора хромосом (гаплоидного), достающегося им от материнской яйцеклетки .
Обычно самцы имеют крылья, которые используют в период роения. У представителей рода Cardiocondyla , кроме крылатых, известны также и бескрылые самцы, ведущие конкурентную борьбу друг с другом (с летальным исходом) за молодых самок в материнском гнезде .
Роль самцов сводится к оплодотворению молодых крылатых самок. Самцы обычно появляются в муравейнике незадолго до брачного лёта и вскоре после спаривания погибают .
Подавляющее большинство особей в муравьиной семье составляют рабочие, являющиеся самками с недоразвитой половой системой (для сравнения, у термитов рабочими особями могут быть также и неполовозрелые самцы), основной ролью которых является забота о семье. Крылья у них отсутствуют, имеется упрощённое строение груди, глаза меньше, чем у самок, или редуцированы, а у части видов отсутствуют.

Для некоторых видов характерен полиморфизм — отличие в размерах, восприимчивости органов чувств и активности среди муравьёв одного вида в зависимости от задач, которые они выполняют в семье .
Часто муравьи-рабочие больших размеров имеют непропорционально большую голову и, соответственно, сильные мандибулы . Таких рабочих называют муравьями- солдатами , так как сильные мандибулы делают их очень эффективными в бою, но в то же время они по-прежнему являются муравьями-рабочими, и их «обязанности», как правило, незначительно отличаются от «обязанностей» других рабочих. У некоторых видов муравьёв нет рабочих средних размеров, что создаёт резкий разрыв между маленькими и большими формами . Например, муравьи-ткачи имеют чёткое бимодальное распределение размеров . У некоторых других видов муравьи-рабочие претерпевают изменения на протяжении всей жизни.
Помимо полиморфизма , рабочие муравьи могут делиться по специализации в семье ( полиэтизм ). Одни муравьи ухаживают за молодью (няньки), другие — участвуют в строительстве гнезда, становятся фуражирами , третьи — чистят помещения, четвёртые хранят запасы жидкой пищи ( медовые бочки ) и т. д.
У некоторых видов муравьёв рабочие особи могут откладывать яйца .
Мощность рабочего муравья возрастает при работе в группе. Например, рабочий волосистого лесного муравья в одиночестве способен развивать мощность 24,2 эрг /сек, а в паре каждый рабочий развивает мощность до 31,6 эрг /сек . У муравьёв рода Formica мозг занимает 0,57 % объёма тела .
Муравьи, как представители насекомых с полным превращением , в своём развитии проходят несколько стадий: яйцо, личинка, куколка и имаго . Цикл развития муравьёв, как и всех других перепончатокрылых , включает полное превращение ( голометаболия ). Из яйца вылупляется личинка — единственная растущая стадия насекомого.

Жизнь муравья начинается с яйца . Если яйцеклетка оплодотворена , то из неё вырастет самка , а если нет, то самец . Обычно яйца хранятся не по отдельности, а небольшими «пакетами». После инкубационного периода из яйца выходит малоподвижная червеобразная личинка , кормлением и уходом за которой занимаются рабочие особи. Наружные покровы личинки могут растягиваться только до определённых пределов, и в ходе роста происходят линьки. Соответственно принято различать несколько возрастных стадий личинки. Для муравьёв типичны четыре личиночные стадии, которые завершаются окукливанием, хотя у некоторых видов может быть три или пять личиночных стадий. Кормление личинки осуществляется при помощи трофаллаксиса , когда муравей отрыгивает жидкую пищу из своего зоба — как и при обмене пищей между имаго , хранящими пищу в «общественных желудках». Личинки могут потреблять и твёрдую пищу, такую как кормовые яйца, куски добычи или семена, принесённые рабочими. У некоторых видов личинок переносят непосредственно к месту, где была захвачена добыча. Перед окукливанием личинка перестаёт питаться и выделяет содержимое своего кишечника. У некоторых видов муравьёв (например, у представителей подсемейств формицины , понерины , Amblyoponinae ) личинки перед окукливанием плетут кокон . Коконы муравьёв в народе неверно называют «муравьиными яйцами». Куколка свободная — придатки имаго внутри неё свободны и не слиты с телом . В зависимости от питания , которое получает личинка, она может развиться в царицу или рабочую особь. Если у вида существует деление рабочих на касты, то питание также определяет, к какой касте будет относиться будущее имаго муравья. Личинки и куколки должны находиться при определённой постоянной температуре, поэтому рабочие часто перемещают их из одной камеры муравейника в другую, с более подходящими условиями . После завершения стадии куколки рабочие муравьи помогают новой особи выйти из неё, так как муравей самостоятельно не в состоянии вскрыть кокон.

Первые несколько дней жизни новые рабочие особи проводят, ухаживая за королевой и расплодом. Затем они обычно занимаются рытьём туннелей и другой работой внутри гнезда. Впоследствии муравей становится защитником гнезда и фуражиром . Эти изменения довольно внезапные и являются примерами временных каст. Объяснение такой последовательности связано с высокой смертностью среди фуражиров .

У большинства видов муравьёв способность к спариванию имеется только у самок (будущих цариц) и самцов. Вопреки распространённому мнению, в некоторых семьях муравьёв могут быть несколько королев (полигинность), а в других может не быть королевы вовсе. Рабочие, способные к размножению, называются гамэргатами ( англ. gamergates ), а семьи, в которых нет королевы, — гамэргатными . Крылатые самцы муравьёв выходят из куколок вместе с будущими царицами и на протяжении своей жизни только питаются и спариваются . Большинство муравьёв моновольтинны, то есть за год развивается только одно поколение . В определённый момент времени, который зависит от вида, крылатые самки и самцы покидают гнездо и отправляются в брачный полёт . Как правило, самцы взлетают раньше самок и выделяют феромоны , заставляющие их следовать за ними. Самки большинства видов спариваются только с одним самцом, но встречаются также виды, самки которых спариваются с десятью и более самцами . После спаривания самка ищет подходящее место для создания нового муравейника. Найдя таковое, она отгрызает себе крылья и роет первую камеру будущего гнезда, а затем начинает откладывать яйца и ухаживать за ними . Царица хранит сперму , полученную во время брачного полёта, и оплодотворяет ею свои яйца. Первые рабочие в новой семье являются слабыми и мелкими по размеру в сравнении с более поздними рабочими особями. Они увеличивают гнездо, добывают корм и ухаживают за расплодом. Так создаются новые семьи у большинства видов, однако существуют виды, у которых колонию образуют сразу несколько королев. В определённое время одна из королев со «свитой» покидает семью и переселяется на новое место . Этот процесс сродни роению у медоносных пчёл .
У муравьёв отмечен широкий спектр стратегий размножения. Например, самки некоторых видов способны размножаться путём однополого размножения ( партеногенеза ), при котором самки появляются из неоплодотворённых яиц (без участия самцов) , а у вида Mycocepurus smithii все особи женского рода . Факультативная или облигатная телитокия экспериментально подтверждена у нескольких видов : Pristomyrmex pungens , Messor capitatus ( Myrmicinae ) , Cerapachys biroi ( Cerapachyinae ) , Cataglyphis cursor ( Formicinae ) , Platythyrea punctata ( Ponerinae ) .
В тропиках муравьи активны круглогодично, а в более холодных регионах переживают зиму в состоянии покоя и бездействия. Формы бездействия разнообразны, и у некоторых видов даже личинки переходят в состояние диапаузы . Однако у большинства видов взрослые особи перезимовывают в состоянии пониженной активности .
Уникальный вариант естественного клонирования существует у малого огненного муравья , самцы и самки которого размножаются клонированием независимо, благодаря чему генофонды обоих полов не смешиваются. У данного вида рабочие особи развиваются из оплодотворённых яиц, матки — из неоплодотворённых диплоидных яиц. В некоторых яйцах, оплодотворённых самцами, все хромосомы матери разрушаются, и из таких гаплоидных яиц развиваются самцы.
Диплоидный набор хромосом у муравьёв имеет самый большой диапазон среди всех насекомых. Он варьирует от двух хромосом у самок Myrmecia pilosula (2n = 2) до 120 хромосом у Dinoponera lucida (2n = 120). У самцов, имеющих гаплоидный набор , он в два раза меньше, то есть у представителей мужского пола Myrmecia pilosula всего одна хромосома .
Размер генома муравьёв варьирует от 180 млн пар оснований ДНК у ( Dolichoderinae ) и Paratrechina longicornis ( Formicinae ) до 470 млн пар у огненного муравья Solenopsis invicta и 500 млн пар у Aphaenogaster treatae и (все три Myrmicinae ), и 550 млн пар у ( Ponerinae ) .
Размножение и расселение муравьёв происходит один раз в году (у некоторых видов — дважды за год).

Во время брачного лёта самки и самцы выходят из гнёзд и скапливаются у их выходов, а затем начинают подниматься на любые возвышенности — травянистые растения, деревья, стены построек и т. п., откуда и взлетают. Более подвижные самцы часто взлетают непосредственно с земли. Самки и самцы из разных гнёзд спариваются в воздухе или на земле. Вскоре после этого самцы погибают, а оплодотворённые самки отправляются на поиски места для будущего гнезда. Найдя подходящее место, самка строит небольшую замкнутую камеру в земле, а потом начинает откладывать яйца . Иногда несколько самок делают такую камеру совместно. Яйца у муравьёв очень мелкие, длиной около 0,5 мм , и всегда склеены в общий комок. Спустя неделю из яиц начинают появляться первые личинки. Молодые личинки остаются в общем «комке», более крупные размещаются группами или отдельно на полу камеры, а иногда (у видов мелких муравьёв) подвешиваются на стенках камеры. Через 2 недели личинки заканчивают рост, и начинается окукливание . К этому времени они становятся крупнее рабочих муравьёв. До выхода из куколок первых рабочих самки ничем не питаются. У некоторых примитивных видов муравьёв самки выходят из гнезда и охотятся на насекомых. Но у преобладающего большинства видов самка так и не покидает гнезда до конца жизни, а личинок выкармливает выделениями слюнных специальных желёз . При этом у самки полностью исчезают летательные мышцы и расходуются запасы жира, накопленные ещё в родительском гнезде. После выхода из куколок первых рабочих они выходят наружу из камеры и начинают добывать пищу. С этого момента самка только откладывает яйца. Все работы в гнезде берут на себя рабочие особи .
Среди муравьёв широко распространены явления временного социального паразитизма. Самки данных видов обычно намного мельче, чем видов, которые самостоятельно основывают гнёзда . Самки малых лесных муравьёв , например, не могут самостоятельно основать новую семью. Для этого им нужна помощь рабочих другого вида — бурого лесного , краснощёкого или серого песчаного муравьёв . Молодая самка рыжего лесного муравья находит муравейник одного из этих видов, потерявший собственную самку, и поселяется в нём. У других временных социальных паразитов, например, у жёлтого пахучего муравья , самка проникает в гнездо вида-хозяина, где имеется своя самка, и оказывается настолько привлекательной для рабочих-хозяев, что они позволяют ей убить собственную самку и занять её место . Некоторое время в гнезде мирно сосуществуют муравьи двух видов, но «хозяева», лишившиеся матки, постепенно вымирают, и новые обитатели полностью сменяют естественным путём рабочих вида-хозяина, целиком заселяют гнездо и перестраивают его .
По-видимому, из-за большой смертности при предыдущих двух типах создания новой семьи у муравьёв возник новый путь — деление материнской общины и обособления её частей . Существует несколько вариантов деления: деление семьи пополам — гесмозис или выделение небольшого отводка — почкование, при котором на некотором расстоянии от родительского гнезда строится дочернее гнездо, или отводок, куда переселяется часть рабочих семьи с расплодом и молодой самкой. Почкованием размножаются семьи бродячих муравьёв , например эцитон Бурчелли . Самка малайских муравьёв ( Carebara ), достигая длины до 2 см, вылетая из гнезда, захватывает с собой нескольких рабочих особей длиной 1—2 мм , которые цепляются своими челюстями за конечности самки .
Муравьиные семьи могут существовать на протяжении долгого периода времени. Королева может жить до 20 лет, рабочие особи живут от нескольких месяцев до 3 лет. Самцы, однако, живут всего несколько недель . Муравьиные королевы живут в 100 раз дольше, чем большинство одиночных насекомых аналогичного размера . Рекордный показатель для муравьёв и социальных насекомых равен 28 годам для матки Lasius niger , которую содержали в лабораторном гнезде . В естественных условиях в течение года население муравейника почти полностью обновляется.
У типичных муравьёв колония состоит из одного гнезда с одной или несколькими царицами, в котором выращивается расплод. Однако известно более 150 видов муравьёв из 49 родов, у которых колонии состоят из нескольких пространственно разделённых гнёзд. В таких полидомных (в отличие от монодомных) колониях пища и рабочие перемещаются между гнёздами . Принадлежность к колонии определяется по реакции рабочих муравьёв, которые определяют, принадлежит ли другая особь к их колонии или нет. Характерный коктейль химических веществ на поверхности тела (также известных как кутикулярные углеводороды или CHCs) образует так называемый запах колонии, который могут распознать другие члены колонии . Некоторые виды муравьёв, по-видимому, менее разборчивы, а у аргентинского муравья ( Linepithema humile ) рабочие, перенесённые из колонии, расположенной на юге США и Мексики, приемлемы в других колониях того же региона. Аналогично рабочие из колоний, обосновавшихся в Европе, принимаются любыми другими колониями в Европе, но не колониями в Америке. Интерпретация этих наблюдений вызывает дискуссии, и одни учёные называют эти крупные популяции суперколониями , а другие — униколониями .
Сохранение всей сложной структуры муравьиной колонии, связей всех особей и их способностей распознавать прочих членов колонии обусловлено трофаллаксисом (обменом проглоченной жидкой пищей) и химической коммуникацией.

Муравьи общаются с помощью феромонов . Эти химические сигналы у муравьёв развиты более, чем у других перепончатокрылых . Как и другие насекомые , муравьи воспринимают запахи своими длинными и тонкими усиками . Парные усики предоставляют информацию о направлении и интенсивности запаха. Так как муравьи проводят жизнь в контакте с землёй, поверхность почвы является хорошим местом, чтобы оставлять след феромона, который может ощущаться другими муравьями. У видов, которые добывают еду в группе, фуражир, нашедший пищу, маркирует свой обратный путь в муравейник , и по этому пути идут другие муравьи, которые также маркируют с помощью феромонов свой обратный путь в гнездо в случае нахождения еды по отмеченному маршруту. Когда источник пищи исчерпан, муравьи больше не маркируют этот маршрут, и запах постепенно рассеивается. Такое поведение муравьёв помогает справиться с изменениями окружающей среды. Например, если установленный маршрут до пищи блокируется препятствием, то фуражиры приступают к поиску нового пути до пищи. Если поиск был успешный, то на обратном пути муравей маркирует кратчайший маршрут своего возвращения в гнездо. По таким успешным маршрутам идут другие муравьи, усиливая оптимальный маршрут и постепенно находя наилучший путь до пищи .
Муравьи используют феромоны не только для прокладывания маршрутов. Раненый муравей выделяет феромон тревоги, призывающий муравьёв издалека и заставляющий всех находящихся рядом особей атаковать врага. Некоторые муравьи используют даже «феромон пропаганды», чтобы запугать врагов и заставить их драться между собой . Феромоны производятся широким спектром желёз : железой Дюфура , ядовитой железой, пигидиальными железами, открывающимися сзади на верхней стороне брюшка , стернальными железами, открывающимися на нижней стороне брюшка около ануса , мандибулярными железами, открывающимися на внутренней стороне жвал . Феромоны также смешиваются с пищей и передаются при трофаллаксисе , распространяя информацию о семье . Это позволяет другим муравьям узнать, в чём нуждается семья (например, в питании или в обслуживании гнезда) . В семьях видов, у которых есть королевы, рабочие начинают растить новую королеву для семьи, если правящая царица не производит нужные феромоны .
Физические сигналы коммуникации могут использоваться муравьями в сочетании с феромонами. Так, муравьи могут общаться при помощи тактильных стимулов (например, при выпрашивании еды ) и звуков. В частности, некоторые муравьи издают стрекочущие звуки , используя для этого сегменты брюшка или жвалы. Звуки применяются для связи между членами семьи или с другими видами . По мнению некоторых учёных, муравьи практически глухи к звуковым волнам в воздухе ; другие учёные оспаривают эту точку зрения . Муравьи весьма чувствительны к вибрациям твёрдых тел. Например, постукивание брюшком или мандибулами свойственно муравьям-древоточцам . Муравьи некоторых подсемейств ( Myrmicinae , Nothomyrmecinae , Ponerinae , Pseudomyrmecinae ) издают звуки с помощью стридулитрума (органа, расположенного на стебельке между постпетиолем и брюшком). Некоторые виды способны к акустической коммуникации даже на стадии куколки. Например, у M. scabrinodis это позволяет куколкам сообщать о своём социальном статусе рабочим-нянькам .


Муравьи атакуют и защищаются, кусаясь, а у многих видов и используя жало, применяемое для инъекций или распыления химических веществ, таких как муравьиная кислота. Вид Paraponera clavata , живущий в Центральной и Южной Америках, имеет, как считается, самый болезненный укус среди насекомых, который обычно не смертелен для человека. По жалящему индексу Шмидта это ужаление имеет самый высокий рейтинг. Ужаление муравьёв вида Myrmecia pilosula может быть смертельным , поэтому для него было создано противоядие . Огненные муравьи имеют уникальный яд, в состав которого входит алкалоид пиперидин . Их укусы болезненны и могут быть опасны для сверхчувствительных людей .

Муравьи рода Odontomachus имеют мандибулы, прозванные челюстями-капканами, которые мгновенно закрываются и являются самыми быстрозащёлкивающимися челюстями у животных . Проведённое исследование показало, что их челюсти защёлкиваются со скоростью от 126 до 230 км/ч , за 130 микросекунд в среднем. Это исследование также установило использование муравьями своих жвал, как катапульт для отбрасывания врагов . До того как нанести удар, муравей широко раскрывает мандибулы и фиксирует их в таком положении с помощью внутреннего механизма. Энергия хранится в толстых группах мышц и резко высвобождается при стимулировании чувствительных волосков на внутренней стороне жвал. Помимо этого, эти жвалы позволяют выполнять медленную и тонкую работу. Челюсти-капканы есть и у других родов: Anochetus , Orectognathus и Strumigenys , а также у некоторых видов из трибы Dacetini , что объясняется конвергентной эволюцией. Малайзийский вид муравьёв имеет настолько гипертрофированные мандибулярные железы, что они занимают почти всё тело муравья. В случае опасности рабочий сокращением мышц разрывает брюшко и разбрызгивает секрет железы, содержащий ацетофенон и другие химические вещества, которые склеивают врага . Самоубийственная защита рабочих также характерна и для бразильского муравья Forelius pusillus , у которых небольшая группа муравьёв каждый вечер остаётся снаружи после закрытия входов и маскирует их . Например, у муравьёв-кочевников крупные рабочие выстраиваются вдоль троп, выставляя жвалы в стороны от тропы, защищая её .

Помимо защиты от хищников, муравьи защищают своё гнездо от патогенов. Некоторые рабочие муравьи занимаются поддержанием чистоты в гнезде, в их задачи также входит вынос за пределы гнезда умерших особей (некрофорез) . Олеиновая кислота, выделяемая мёртвыми муравьями, запускает некрофорезное поведение рабочих . У некоторых видов, например у аргентинского муравья , некрофорезное поведение запускается отсутствием характерных химических веществ (долиходиаль и иридомирмецин), которые присутствуют в кутикуле живых особей .
Гнездо может быть защищено от природных угроз, таких как наводнение или перегрев, с помощью особенностей архитектуры . Рабочие муравьи вида Cataulacus muticus , живущие в полостях деревьев, при наводнении начинают пить воду, попавшую в гнездо, и выделять её наружу . , живущие в полостях древесины мангровых лесов, при погружении под воду могут переключаться на анаэробное дыхание .
Многие животные могут обучаться с помощью подражания, но муравьи, возможно, единственная группа помимо млекопитающих , у которых наблюдается интерактивное обучение . Осведомлённый фуражир вида Temnothorax albipennis приводит товарища к недавно открытому источнику питания с помощью . «Ученик» получает информацию от «лидера». При этом «лидер» и «ученик» всегда находятся в контакте и следят за продвижением друг друга: «учитель» замедляется, когда «ученик» отстаёт, и ускоряется, когда ученик слишком близко . При этом сам по себе учитель достиг бы корма в четыре раза быстрее. Получив урок, «ученики» зачастую сами становятся «учителями», таким образом информация о положении корма распространяется по всему гнезду .
Эксперименты с семьями Cerapachys biroi показали, что муравьи могут выбирать себе роль в муравейнике согласно их опыту. Целое поколение идентичных рабочих было разделено на две группы, чьи результаты поиска еды полностью контролировались. Фуражиры из первой группы всегда вознаграждались добычей, в то время как поиски пищи рабочих из второй группы всегда были неудачны. В итоге рабочие первой группы активизировали свои попытки поиска пищи, а фуражиры второй группы отправлялись искать пищу всё реже и реже. Через месяц муравьи из первой группы продолжали работать фуражирами, в то время как муравьи из второй группы сменили свою специальность на заботу о потомстве .

По мере роста муравьиной общины и укрепления её целостности разделение функций становится всё более глубоким: число «профессий» рабочих муравьёв возрастает, а специализация каждой особи сужается. В пределах касты рабочих муравьёв выделяют полиэтические группы — группы особей, занятых выполнением определённого круга обязанностей. В пределах одной полиэтической группы может быть несколько функциональных групп особей, выполняющих задачи, разные по конкретному содержанию, но сходные по типу поведения исполнителей . Разделение функций, или полиэтизм , может быть возрастным или постоянным .


Обычно муравьи живут в гнёздах. Сложные гнёзда строятся большинством видов муравьёв, но есть виды, ведущие кочевой образ жизни и не строящие постоянных гнёзд. Муравьи могут создавать подземные гнёзда или строить их на деревьях. Гнёзда можно найти на земле, под камнями или в брёвнах, под корягами, в полых стеблях или даже в желудях . Материалы, используемые во время строительства, включают в себя почву и растительные вещества , муравьи тщательно выбирают их для своего гнезда; Temnothorax albipennis избегают участков с мёртвыми муравьями, так как это может указывать на наличие болезней или вредителей. Они немедленно отказываются от строительства гнезда при первых признаках угрозы . Муравьи-листорезы строят сложные гнёзда больших размеров под землёй, характеризующиеся особой конструкцией, обеспечивающей поддержание постоянных и оптимальных показателей вентиляции, влажности воздуха и т. п., обеспечивающих выращивание грибов . У пустынных муравьёв гнёзда могут уходить под землю на глубину до 4 м, а по косвенным данным — даже до 10 м .
В тропических лесах многие виды муравьёв делают открытые гнёзда в кронах деревьев. Одни из них строят шаровидные висячие гнёзда из картона, другие — из листьев, третьи — из паутинных нитей . Так, ряд древесных видов, например обитающий в африканских саваннах , строит картонные гнёзда вокруг веток. Рабочие муравьёв-ткачей строят гнёзда на деревьях, соединяя живые листья вместе и закрепляя их шёлком, выделяемым их личинками — прикасаясь головой личинок то к одному, то к другому, они «сшивают» края листьев . Аналогичные методы строительства гнезда есть у видов из рода полирахис .
Некоторые виды, например с примитивной социальной организацией , не занимаются постройками видоспецифичного гнезда и могут поселяться в пустотах под камнями, дренажных канавах, в кронах деревьев, просто под повалившимися деревьями и опавшими листьями . Многие виды сооружают гнёзда в древесине, например пахучий и красногрудый муравьи-древоточцы .
Кочевые муравьи Южной Америки ( Ecitoninae ) и африканские Dorylus не строят постоянных гнёзд, а вместо этого выбирают кочевой образ жизни и временные гнёзда-бивуаки , созданные из сцепленных друг с другом тел рабочих .


Большинство муравьёв являются универсальными хищниками, падальщиками и косвенными потребителями листвы . Основу питания почти всех муравьёв составляют два компонента: белковый и углеводный. При этом углеводный в основном потребляется взрослыми особями, а белковый — личинками.
Вся пища, которую собирают муравьи, приносится в гнездо и там распределяется между всеми членами семьи. По данным многолетних наблюдений западногерманского исследователя Г. Велленштайна, питание рыжих лесных муравьёв (по массе) :
У ряда видов, преимущественно примитивных муравьёв, имеется пищевая специализация . Например, представители подсемейства понерины специализированы на питании определённой группой беспозвоночных. Также род Centromyrmex и представители, по крайней мере, девяти родов муравьёв — термитами . Триба целиком специализируется на поедании других муравьёв. Австралийские муравьи-бульдоги охотятся даже на пчёл , убивая их с помощью жала .
Обычным источником сахаров для муравьёв служит падь, выделяемая различными насекомыми, сосущими соки растений, — преимущественно представителями семейств Настоящие тли ( Aphididae ), Кокциды ( Coccidae ) и Белокрылки ( Aleyrodidae ). Три вида из рода муравьёв-древоточцев собирают камеди , выделяемые растениями . Мирмицины часто питаются семенами. Многие виды, являющиеся обитателями саванн и пустынь, в период засухи занимаются сбором и запасанием семян . Высокоспециализированными (поедателями семян) являются муравьи-жнецы — так, рацион Messor pergandei на 97 % составляют семена .

Немногие виды развили специальные пути получения пищи. Грибы разводят и ими питаются около 190 видов муравьёв . Более примитивные виды используют в качестве субстрата для грибниц кусочки насекомых и фекалии, а наиболее эволюционно продвинутые роды — муравьи-листорезы Atta и Acromyrmex — срезанные части листьев живых растений. Если вытащить данную массу из муравейника, то вскоре на ней появятся плодовые тела гриба, но в гнёздах муравьи не допускают этого . Муравьи-листорезы питаются исключительно гифами грибов , которые растут только в пределах их гнезда. Они постоянно собирают листья, которые приносят в гнездо, измельчают и располагают в грибных садах. Специализация рабочих зависит от их размера — большие муравьи с сильными челюстями разрезают листья; более мелкие — разжёвывают их, а совсем маленькие ухаживают за грибами. Муравьи и их личинки питаются гифами грибов . Симбиотические бактерии из рода Streptomyces , живущие на поверхности тела муравья, производят особые вещества, которые убивают «грибы-сорняки» рода . Разведение муравьями и перенос грибков (виды аскомицетов из отряда ) молодыми самками -основательницами также обнаружено у древесных видов рода Azteca из подсемейства долиходерины .
Муравьи-фуражиры в поисках пищи уходят от гнезда на расстояние до 200 м и обычно находят путь назад, используя «запах» тропы. Некоторые муравьи ищут пищу ночью. Днём муравьи-фуражиры из жарких и засушливых районов рискуют умереть от обезвоживания, поэтому умение найти кратчайший путь до гнезда очень важно для них. использует визуальные ориентиры в сочетании с другими сигналами для навигации . При отсутствии визуальных ориентиров близкие к ним красные фаэтончики начинают отслеживать направление и отсчитывать расстояние внутренним шагомером , подсчитывая, сколько шагов они делают в каждом направлении. Интегрируя эту информацию, муравьи находят кратчайший путь до гнезда . Некоторые виды муравьёв способны использовать для навигации магнитное поле Земли . Муравьиные сложные глаза имеют специальные клетки, обнаруживающие поляризованный свет Солнца, используемый для определения направления . Эти поляризационные детекторы чувствительны к ультрафиолетовой части спектра .
Рабочие особи бескрылые, а самки и самцы — крылатые, однако самки утрачивают крылья после брачного полёта. Таким образом, в отличие от своих предков ос, муравьи преимущественно передвигаются, ползая. Только представители 4 родов муравьёв способны прыгать с помощью ног: Gigantiops (Южная Америка, Formicinae ), Harpegnathos (юго-восточная Азия, Ponerinae , Myrmecia (Австралия, Myrmeciinae ) и Odontomachus (Ponerinae). Причём представители Odontomachus могут прыгать и вперёд (с помощью ног), и назад (с помощью быстро закрывающихся челюстей) . Например, прыжок Harpegnathos saltator происходит с помощью синхронизации средних и задних лап . Есть несколько видов планирующих муравьёв , включая Cephalotes atratus . Это, в некотором роде, общая черта большинства древесных муравьёв. Такие муравьи в состоянии контролировать направление падения во время полёта .
Другие виды муравьёв могут образовывать мосты через водные преграды, под землёй или между растениями. Некоторые виды также создают плавающие плоты, помогающие им пережить наводнение. Такие плоты могут играть определённую роль при колонизации муравьями островов . Вид полирахис Соколова , найденный в австралийских мангровых лесах , может плавать и жить в подводных гнёздах. Поскольку у них нет жабр, они дышат в воздушных карманах подводного гнезда .
В пустынях Азии и Северной Африки обитают муравьи-бегунки из рода Cataglyphis . Передвигаются они, исключительно бегая, держа брюшко поднятым вертикально вверх. Активны они в самое жаркое время дня, когда почва нагревается до +50…+70 °C . Большая скорость передвижения объясняет тем, что им требуется за как можно меньшее время обежать большую территорию и при этом «не свариться» от жары почвы .
Большинство высокоорганизованных муравьёв прокладывают так называемые «дороги» на поверхности земли или подземные тоннели, ведущие из гнёзд в окружающую территорию. Эти дороги образуются на месте часто посещаемых пахучих троп. Муравьи охраняют их, постоянно обновляют и убирают с них любые препятствия .

Миграции свойственны только для бродячих муравьёв подсемейств Ecitoninae и Dorylinae , обитающих преимущественно в тропиках Африки и Южной Америки. Наиболее крупные колонии, насчитывающие до 22 миллионов особей, известны у африканского вида Dorylus wilverthi . Кочёвка длится несколько дней, стоянка — от недели до трёх месяцев. У бродячих муравьёв рода Eciton оседлая и кочевая фазы чередуются — каждая из них длится по 2—3 недели . Смена этих фаз определяется репродуктивным циклом . Вместо постоянных гнёзд они образуют временные гнёзда-биваки , созданные из сцепленных друг с другом тел рабочих особей .
Долгая коэволюция муравьёв с другими видами привела к появлению разнообразных отношений между ними — мимикрии , комменсализма , паразитизма и мутуализма .
Не все муравьи имеют одинаковый вид общества. Австралийские муравьи-бульдоги являются одними из самых крупных и примитивных муравьёв. Как и почти все остальные муравьи, они являются общественными насекомыми, но их поведение в обществе слабо развито по сравнению с другими видами. Каждый муравей охотится в одиночку, используя лишь свои большие глаза, вместо химических ощущений для поиска добычи .
Некоторые виды (например, дерновый муравей ) атакуют и захватывают соседние семьи муравьёв. Другие, менее экспансионистские, но такие же агрессивные, вторгаются в гнездо, чтобы украсть яйца и личинки, которые они либо едят, либо выращивают из них рабов. Крайность такой специализации — это муравьи-амазонки , которые не в состоянии самостоятельно питаться и должны захватывать рабочих, чтобы выжить . Захваченные рабочие Temnothorax развили стратегию борьбы с рабовладельцами, они разрушают все женские куколки своих хозяев вида Protomognathus americanus , но оставляют в живых самцов (которые не принимают участия в угоне рабов) .

Муравьи узнают членов семьи по запаху, который исходит от углеводородных выделений, находящихся на их экзоскелете. Если муравей отделён от материнской семьи, он со временем теряет запах семьи. Любой муравей, который входит в гнездо и не имеет соответствующего запаха, будет атакован .
Паразитарные виды муравьёв проникают в гнёзда муравьёв-хозяев и становятся социальными паразитами. Такие виды, как Strumigenys xenos , являются полностью паразитарными и не имеют рабочих. Они полностью зависят от собранной пищи вида-хозяина . Такая форма паразитизма встречается у многих родов муравьёв, и виды-паразиты тесно связаны со своими видами-хозяевами. Для проникновения в гнездо используются различные методы. Паразитарная королева может проникнуть в гнездо ещё до того, как вылупится первое поколение рабочих, и участвовать в формировании запаха семьи. Другие виды применяют феромоны, чтобы запутать или обмануть муравьёв-хозяев во время проникновения в гнездо королевы. Некоторые просто сражаются, чтобы попасть внутрь гнезда .
Конфликты между самцами и самками встречаются у некоторых видов муравьёв и связаны, по-видимому, с конкуренцией между производителями потомства. В экстремальных формах конфликт включает в себя производство клонального потомства. Крайняя форма межполового конфликта наблюдается у малых огненных муравьёв , чьи царицы производят диплоидных дочерей однополым размножением , а самцы являются клонами своих отцов, так как диплоидное яйцо теряет материнский вклад у гаплоидных самцов .
Некоторым муравьям свойственен клептопаразитизм : они могут воровать пищу у таких хищных растений, как росянка или жирянка . Многие муравьи питаются другими общественными насекомыми. Одни виды ( Megaponera и Termitopone ) специализируются на термитах, а другие ( Cerapachyinae ) на муравьях ( мирмекофагия ) . Некоторые виды термитов, включая , объединяются с другими муравьями для противостояния хищным муравьям . Тропическая оса Mischocyttarus drewseni покрывает ножку своего гнезда химическим муравьиным репеллентом . Предполагается, что многие тропические осы могут строить свои гнёзда на деревьях и покрывать их репеллентом для защиты от муравьёв. Ядовитые железы жалящего аппарата пчёл из родов Trigona и Melipona вырабатывают химические вещества, используемые для защиты от муравьёв .
На основании временного социального паразитизма в ходе эволюции у некоторых видов муравьёв возникла своеобразная форма паразитизма , получившая название «рабовладельчества» . Муравьи некоторых видов грабят соседние гнёзда муравьёв, где они добывают куколок и приносят их в своё гнездо. Из этих куколок воспитывают «рабов» . Сходство с рабовладением в человеческом обществе тут чисто внешнее. «Рабы» муравьёв выполняют в гнезде «рабовладельца» те же работы, что они выполняли бы и в родном гнезде, только выращивают расплод не своего, а чужого вида . У кроваво-красного муравья-рабовладельца собственные рабочие добывают пищу, охраняют гнездо и частично занимаются уходом за потомством, хотя в основном эту работу выполняют «рабы». У муравьёв-амазонок рабочие занимаются только добыванием куколок «рабов» и не способны даже самостоятельно питаться . В целом социальный паразитизм у муравьёв характерен в основном для подсемейств мирмицины и формицины , а рабовладение встречается в родах Polyergus , Raptiformica , Rossomyrmex и Strongylognathus . Обнаружено более 200 паразитических видов муравьёв . Обитающая в Южной Франции ( Epimyrma vandeli ), проникая в гнездо хозяина — муравья вида Leptothorax recedens — убивает его самку. Рабочие особи принимают её. Своих рабочих особей у эпимирмы нет, и из отложенных самкой яиц выходят только крылатые самки и самцы .

Муравьи имеют симбиотические связи с рядом видов, в том числе с другими муравьями, насекомыми, растениями, грибами. Некоторые виды членистоногих проводят часть своей жизни в гнёздах муравьёв, где охотятся на них, их личинок и яйца, на сохранённую муравьями пищу или прячутся от хищников. Эти инквилины могут иметь сходство с муравьями. Природа муравьиной мимикрии (мирмекоморфии) различна. В некоторых случаях это пример бейтсовской мимикрии , в других — васманновой мимикрии .

Трофобиотические отношения с разными насекомыми широко распространены среди муравьёв. Так, тли и другие членистохоботные насекомые выделяют сладкую жидкость, называемую падью , когда они питаются соком растений. Сахар пади является высококалорийной пищей, которую собирают многие виды муравьёв , прежде всего из высокоразвитых подсемейств ( долиходерины , формицины , мирмицины ) . В некоторых случаях тля выделяет падь в ответ на прикосновения муравья своими антеннами. Муравьи, в свою очередь, оберегают скопления тлей от хищников и перемещают их на лучшие растения для кормления. При переходе на новое место многие семьи берут с собой тлей, чтобы обеспечить себе бесперебойный источник сахаров. Муравьи также собирают сладкие выделения мучнистых червецов . Помимо равнокрылых (Homoptera: Aphididae , Coccidae , Pseudococcidae , Membracidae , Cicadellidae , Aleyrodidae и др.) различные виды трофобиотических отношений развиты у муравьёв с некоторыми бабочками ( голубянки , риодиниды , листовёртки ) и клопами ( краевики , настоящие щитники , полушаровидные щитники ) , а также с представителями отряда перепончатокрылых — личинками пилильщиков Blasticotoma filiceti .

У ряда видов бабочек мирмекофильные гусеницы живут в муравейниках , находясь с муравьями, например, с родом мирмика , в симбиотических отношениях . Примерно половина всех видов голубянок так или иначе связана в цикле своего развития с муравьями . Взаимосвязи между муравьями и голубянками по своему типу варьируют от факультативных до облигатных и от мутуализма до паразитизма . Гусеницы и куколки голубянок обладают комплексом химических и акустических сигналов для контролирования поведения муравьёв. Также они выделяют из дорзальной нектароносной железы сладкую жидкость, привлекающую муравьёв. Муравьи, ощупывая усиками этот орган, вызывают рефлекторное выделение гусеницей жидкости, которая, вероятно, содержит феромоны лихневмоны , обусловливающие поведение муравьёв. Некоторые гусеницы голубянок и риодинид обладают также звукоиздающими органами, влияющими на поведение муравьёв , — одни виды имеют только шаровую щетинку на теле гусеницы, другие — вместе со звуковой ресничкой, при отсутствии нектароносной железы . Все виды мирмекофильных голубянок приурочены к луговым муравейникам .
Другие гусеницы эволюционировали от мирмекофилов в мирмекофагов : они выделяют феромон , который заставляет муравьёв думать, что гусеница является одной из личинок муравьёв. Такие гусеницы проникают в муравейники и питаются личинками самих муравьёв .
Представители мирмекофильных разнокрылых бабочек рода Cyclotorna (из надсемейства Zygaenoidea , эндемики Австралии ) являются единственными среди бабочек эктопаразитами . Гусеницы (первоначально похожие на уплощённых мокриц, а на последних стадиях более ярко окрашенные) являются эктопаразитами равнокрылых насекомых , таких как цикадки ( Cicadellidae ) и листоблошки ( Psyllidae ). Высосав жертву, молодые гусеницы покидают её и становятся хищниками личинок мясных муравьёв ( Iridomyrmex purpureus ), по-видимому, используя химические вещества ( феромоны ), индуцируют муравьёв для переноски гусениц в свои муравейники (перед окукливанием покидая их гнёзда) .
Большинство мирмекофилов питается различными отбросами муравьёв или гниющим строительным материалом гнезда, но многие являются «нахлебниками» . У многих из них выработались специальные структуры для сожительства с муравьями. У многих из них на теле имеются специальные железы, выделяющие вещества, которые муравьи слизывают. Например, у пауссид они находятся на усиках, у жуков-ощупников — по бокам тела .

К мирмекофилам относятся также некоторые другие жуки — например, карапузики , стафилины и прочие. Так, 19 родов стафилинов сожительствует с бродячими муравьями, 17 родов — с огненными муравьями и 15 родов — с представителями подсемейства формицины . Стафилины выпрашивают у муравьёв-хозяев корм, прикасаясь к нижней губе, и защищаются в случае нападения, выделяя репелленты. Стафилины, сожительствующие с бродячими муравьями, мимикрируют под муравьёв формой и окраской своего тела (пример бейтсовской мимикрии). Стафилины из рода Dinarda в муравейниках питаются мёртвыми муравьями, отходами и клещами , обитающими на живых муравьях . Нимфы клещей-уроподид прикрепляются к телу рабочих и питаются слюной, выделяемой муравьями тогда, когда они облизывают друг друга .
В гнёздах нескольких родов муравьёв ( формика , лазиус , мирмика ) встречается мелкий (3 мм) бескрылый сверчок-муравьелюб ( Myrmecophilus acervorum ) , который постоянно облизывает своих хозяев, питаясь выделениями их кутикулы .
Пример жуков- комменсалов муравьёв — некоторые виды бронзовок , например Potosia metallica , личиночные стадии которой проходят в муравейниках рыжих и малых лесных муравьёв, красногрудых муравьёв-древоточцев и некоторых других .
Конвергенция (возможно, одна из форм мимикрии ) видна на примере яиц палочника , имеющих съедобные элайосомоподобные придатки. Муравьи приносят их в гнёзда, где выводятся личинки палочников . Кочевые муравьи переносят с собой клещей рода Larvamima ( Larvamimidae ), так как те формой своего тела напоминают мелких личинок муравьёв .

Муравьи-грибководы из трибы Attini , в том числе муравьи-листорезы, выращивают некоторые виды грибов из родов Белошампиньонов и Белонавозников семейства Шампиньоновые . В этом симбиозе муравьёв и грибов выживание каждого из видов зависит от другого. Муравьи Allomerus decemarticulatus развили трёхсторонний симбиоз с растением-хозяином и липким грибом, которого они используют как ловушку для насекомых .
Myrmelachista schumanni создают сады дьявола , убивая окружающие растения и расчищая место для Duroia hirsuta , в стеблях которых они живут. Такое модифицирование леса обеспечивает муравьям больше места для гнёзд . Некоторые деревья имеют дополнительные внецветочные нектарники, которые служат пищей для муравьёв, а те, в свою очередь, защищают растения от травоядных насекомых .
Многие тропические виды деревьев имеют семена, распространяемые муравьями . Мирмекохория широко распространена в Африке и Австралии . Выживание и распространение некоторых растений в пожароопасных пастбищных экосистемах особенно зависит от муравьёв. Мирмекохория свойственна и многим лесным травам умеренного пояса (копытень, кислица, некоторые фиалки, ожика и др.). Многие мирмекохорные семена имеют специальные внешние питательные придатки, элайосомы, которыми муравьи питаются .


У ряда видов тропических деревьев способ привлечения муравьёв достиг высокого совершенства: они обладают не только экстрафлоральными нектарниками, содержащими сахара, но и подходящими местами для устройства муравьями гнёзд и даже обеспечивают муравьям твёрдые белковые и жировые корма. Многие виды акации ( , , ) привлекают муравьёв при помощи «пищевых телец» (тельца Бельта) , находящихся на кончиках листьев, и наличием полостей в специализированных вздутых колючках, в которых находятся места для устройства гнёзд. В них селятся муравьи одного из видов рода псевдомирмекс , которые защищают своё «гнездовое» дерево от насекомых-фитофагов .
У хищных растений рода Непентес наблюдаются почти все формы отношений с муравьями: нейтральные, хищник — жертва (муравьи как важнейшая добыча) и мутуалистические. Муравьи Camponotus schmitzi не только живут в стеблях непентеса , но и способны воровать из его кувшинчиков упавших туда насекомых . Муравьи также нападают на млекопитающих и очищают от других растений определённый участок вокруг дерева, уничтожая тянущиеся к нему ветви. Обнаружено несколько мутуалистических систем «муравьи-растения» , например Crematogaster — ; Tetraponera — ; Ocotea — Myrmelachista ; — Pseudomyrmex . Муравьи подсемейства долиходерины — виды из рода Azteca — селятся на деревьях рода Cecropia , предоставляющих им жильё и пищу . В данных примерах высшей ступени мутуалистических взаимоотношений между муравьями и растениями первые обеспечивают дереву защиту, получая, в свою очередь, территорию, удовлетворяющую потребностям в пространстве обитания и пище . Исследования с помощью изотопных маркировок показали, что растение также получает азот от муравьёв . Другим примером такого эктосимбиоза является дерево , связанное с остробрюхими муравьями . Тропическое растение Costus woodsonii выделяет из прицветников нектар , и на растении, сменяя друг друга, в зависимости от времени года, обитают два вида муравьёв: и малый огненный муравей .
Кочевые муравьи , ведущие кочевой образ жизни, известны своими «рейдами» . Подобные скопления муравьёв вида эцитон Бурчелли привлекают птиц, следующих за муравьями , таких как древолазовые или полосатые муравьеловки , которые используют муравьёв как загонщиков насекомых и других мелких членистоногих .

Более чем 250 видов птиц занимаются муравлением . Они отдыхают на муравьиных гнёздах, где муравьи, забравшись к ним на крылья и перья, удаляют эктопаразитов. Другие птицы «купаются» в муравейниках или раздавливают муравьёв и смазывают ими оперение , преследуя аналогичные цели . При этом птицы выбирают муравьёв (из подсемейств формицины и долиходерины ), которые выбрасывают струи муравьиной кислоты , обладающей инсектицидными свойствами.
Грибы из родов Кордицепс и Ophiocordyceps заражают муравьёв, заставляя их забираться на растения и повисать на своих мандибулах . Грибок убивает муравья, на его останках прорастает мицелий и плодовые тела. Возможно, гриб изменяет поведение муравьёв, чтобы помочь рассеять свои споры в местах обитания, которые лучше всего подходят для гриба . Также на представителях рода формика паразитируют два других гриба — и . Случаи заражения ими начинаются поздним летом. Вечером заражённые муравьи покидают свои гнёзда, забираются на верхушки растений, цепляясь за стебли жвалами и ножками. Вскоре после этого они погибают и приклеиваются к стеблям растений при помощи клейких выделений из сочленений груди. На следующий день сквозь тело муравьёв прорастает мицелий гриба, а на третий день развиваются конидии .

Веерокрылые также паразитируют и управляют муравьями, заставляя их подниматься на стебли травы, чтобы помочь паразитам в поисках партнёра для спаривания .
Тетрадонематиды Myrmeconema neotropicum , инфицирующие Cephalotes atratus , изменяют чёрный цвет брюшка рабочих на красный. Паразит также изменяет поведение муравья, заставляя высоко держать своё брюшко. Птицы поедают таких муравьёв. Помётом заражённых птиц, собранным другими муравьями, кормят молодняк, и цикл повторяется .
Также известно несколько видов гельминтов , паразитирующих на муравьях (представители Allantonematidae , Mermithidae и других). Например, ланцетовидная двуустка ( Dicrocoelium lanceatum ) и рабдитиды вида , паразитирующие на представителях рода формика , некоторые представители мермитид (в том числе род ). Они проходят весь цикл развития в одном хозяине-муравье. Так, муравьи могут поедать цисты ланцетовидной двуустки. Попадая в зоб муравьёв, церкарии пробуравливают его стенку и выходят в полость тела. Одна из них проникает в подглоточный ганглий, где образует цисту — так называемый «Hirnwurm» или «мозговой червь» в области отхождения нервов к ротовым органам . В дальнейшем происходят резкие изменения поведения заражённого муравья: он выходит из гнезда и подолгу висит, прицепившись мандибулами к верхушкам травянистых растений. Здесь заражённого муравья и поедают дефинитивные хозяева ланцетовидной двуустки .
Все виды наездников семейства Eucharitidae являются паразитоидами муравьёв . Каждый из видов паразитирует только на одном виде муравьёв. Наездники откладывают яйца на посещаемые муравьями растения. Вылупляющиеся личинки прикрепляются к рабочим особям муравьёв и переносятся на них в гнездо, где ищут личинок муравьёв, к которым прикрепляются внешне или проникают внутрь, после чего развитие прекращается до окукливания личинки муравья. После окукливания личинка наездника мигрирует в полость тела куколки, где начинает питаться, а затем окукливается. Имаго наездников могут какое-то время оставаться в муравейнике под защитой муравьёв, а затем выходят из гнезда для спаривания . Муравьи не способны отличить личинок паразитоидов от собственного расплода благодаря химической мимикрии .
Жук из семейства пластинчатоусых обитает в гнёздах муравьёв рода Solenopsis и изредка поедает куколок муравьёв . Также в муравейниках паразитируют жуки из рода ломехуза , имаго и личинки которых питаются яйцами и личинками муравьёв. Ломехузы способны постепенно приводить семьи муравьёв к деградации и гибели. Муравейники, заражённые ломехузами, нельзя использовать для искусственного расселения в целях защиты леса от вредителей .
Мухи Старого света из рода Bengalia охотятся на муравьёв и клептопаразитируют , выхватывая у них из челюстей добычу или выводок . Бескрылые и безногие самки малайзийских горбаток живут в гнёздах муравьёв рода Aenictus , где о них заботятся муравьи . Горбатки из рода Pseudacteon и некоторые представители рода апоцефалус являются паразитами муравьёв. Взрослые самки мух с помощью яйцеклада во время быстрой воздушной атаки откладывают в муравья яйцо. Личинка определённое время развивается, а затем мигрирует в голову, где она продолжает питаться гемолимфой и тканями, вызывая постепенную гибель муравья. Окукливание происходит там же, после чего у муравья отпадает голова . Личинки муравьиных журчалок обитают в муравейниках и питаются муравьиным расплодом или падалью. Самка мухи откладывает свои яйца недалеко от гнезда муравьёв, а затем, используя химическую мимикрию, личинки проникают внутрь . Среди двукрылых насекомых ( Diptera ) ассоциация с муравьями отмечена среди многих видов горбаток (Phoridae), у некоторых мух-журчалок ( Syrphidae ) из подсемейства Microdontinae , а также у длинноусых из семейств комары ( Culicidae ), мокрецы ( Ceratopogonidae ), галлицы ( Cecidomyiidae ) и почвенные комарики ( Sciaridae ) . Также к настоящим паразитам следует относить гамазовых клещей , которые питаются гемолимфой муравьёв .

Южноамериканские древолазы питаются в основном муравьями, а токсины на их коже могут состоять из яда муравьёв . Некоторые ящерицы также питаются исключительно муравьями — например, молох питается только муравьями-фуражирами , в основном , которых ловит с помощью липкого языка. Многие птицы, например дятлы , певчие птицы , пищухи , также любят лакомиться муравьями . Личинки муравьиных львов ловят насекомых, в том числе и муравьёв в свои ловчие ямки конической формы, выкопанные в песке .
Муравьеды , панголины и некоторые виды сумчатых Австралии (например, сумчатый муравьед ) специализируются на питании только муравьями и для этого выработали целый ряд приспособлений — длинный липкий язык для захвата насекомых и сильные когти, чтобы разрывать муравьиные гнёзда. Трубкозуб питается муравьями родов , Camponotus , Crematogaster , Dorylus . Бурые медведи также питаются муравьями, и около 12 %, 16 % и 4 % от их объёма фекалий, соответственно, весной, летом и осенью составляют муравьи .
Муравьи — это почти 2 % от всего видового многообразия животного мира и до 80 % биомассы насекомых (около 20 % биомассы всех животных) . По оценкам биологов в мире обитает около 20 квадриллионов муравьёв (20 × 10 15 ) муравьев с общей биомассой в 12 мегатонн сухого углерода. Это превышает совокупную биомассу диких птиц и млекопитающих и составляет 20 % биомассы человека. Численность муравьев распределяется на Земле неравномерно, достигая максимума в тропиках и в шесть раз различаясь по местам обитания . В лесах умеренного пояса и тропических лесах муравьи обеспечивают перемещение и аэрацию грунта. Многие земляные виды муравьёв являются почвообразователями, рыхлящими, перемешивающими, удобряющими почву . Их почвообразующая деятельность заключается в перемешивании почвы, изменении её механического и химического состава. В дождевых тропических лесах 99,9 % питательных веществ оставалось бы в верхнем слое почвы, если бы не переносилось вглубь, в том числе и муравьями . Муравьи вместе с термитами могут увеличивать урожайность в регионах с сухим и жарким климатом, где отсутствуют дождевые черви. В экспериментальных условиях в Австралии они увеличивают урожай пшеницы на 36 % .
Многие муравьи также представляют собой важные звенья в пищевых цепях как хищники, питающиеся беспозвоночными . Муравьи, поселяющиеся в древесине или устраивающие свои гнёзда в старых пнях, принимают участие в механическом разрушении мёртвой древесины , ускоряя процессы её разложения. Муравьи, питающиеся семенами, непосредственно способствуют расселению многих видов растений. Семена некоторых растений (так называемых мирмекохорных растений ) расселяются только муравьями ( копытень , фиалки , пролесок и другими). В степях и пустынных районах семена многих растений разносятся исключительно муравьями . Муравьи — единственные насекомые, которые рассеивают семена в больших количествах, на всех континентах и практически во всех экосистемах . Всего в мире насчитывается около 3000 видов мирмекохорных растений (около 1 % флоры) , а в Европе более 275 таких видов .
Также муравьи служат пищей для многих животных.
Муравьи — одни из самых мощных биологических агентов эрозии минералов и рекордсмены по улавливанию углекислого газа из атмосферы. Под воздействием муравьёв процесс превращения таких минералов, как силикаты магния и кальция в карбонат кальция и магния (MgCO 3 и CaCO 3 , последний это основной компонент известняка, мрамора и мела) ускоряется от 50 до 300 раз. Такая обусловленная муравьями эрозия минералов могла сыграть важную роль в кайнозойском похолодании на Земле .
Из 42 космополитных видов муравьёв двенадцать имеют большое экологическое, сельскохозяйственное или иное (как вредители в домах) значения: Anoplolepis gracilipes , Linepithema humile , Monomorium pharaonis , , Paratrechina longicornis , Pheidole megacephala , Solenopsis geminata , Solenopsis invicta , Tapinoma melanocephalum , Technomyrmex difficilis , Trichomyrmex destructor и Wasmannia auropunctata .
| Регион |
Количество
видов |
|---|---|
| Неотропика | 2162 |
| Неарктика | 580 |
| Европа | 180 |
| Африка | 2500 |
| Азия | 2080 |
| Меланезия | 275 |
| Австралия | 985 |
| Полинезия | 42 |
Муравьи обитают по всему миру, кроме Антарктики , Исландии , Гренландии и некоторых удалённых островов. Благодаря широкому распространению, многочисленности и хорошо заметным гнёздам муравьи повсеместно известны людям . Муравьи занимают различные экологические ниши и в состоянии использовать широкий ассортимент пищевых ресурсов, будучи прямыми или косвенными потребителями листвы, или падальщиками и хищниками. Большинство видов муравьёв всеядны, но некоторые специализируются на определённой пище. Их экологическое господство может быть измерено значением их доли в общей биомассе . Различные оценки показывают, что муравьи составляют от 10 % до 20 % (в тропиках до 25 %) от общей биомассы наземных животных, превосходя долю позвоночных . Более 200 видов муравьёв встречаются далеко за пределами своего естественного нативного ареала, а 42 вида муравьёв стали космополитными благодаря человеческой коммерции. Двадцать из них имеют закрепившиеся популяции во всех 7 биогеографических регионах (кроме Антарктики): Афротропика, Палеарктика, Неарктика, Неотропика, Индо-Малайзия, Австралазия и Океания. Из 42 космополитных муравьиных видов 35 (83 %) родом из Старого Света , а 7 (17 %) вышли из Нового Света .
Пять видов муравьёв ( Anoplolepis gracilipes , Linepithema humile , Pheidole megacephala , Solenopsis invicta и Wasmannia auropunctata ), распространившихся в мире с помощью коммерции человека, включены в список 100 самых опасных инвазивных видов организмов , деструктивно влияющих на экологические связи местных биот . Один вид ( фараонов муравей ) был завезён в 2013 году в Гренландию (аэропорт Kangerlussuaq/Sondre Stromfjord), где ранее муравьёв не находили .

Муравьи выполняют множество экологических функций, полезных для человека, в том числе регулирование количества насекомых-вредителей и аэрацию почвы . Благодаря активному хищничеству муравьи активно уничтожают многие виды насекомых-вредителей (включая гусениц бабочек и ложногусениц пилильщиков), отдают предпочтение видам, встречающимся в больших количествах. Исключением можно считать выкормки дубового шелкопряда , где, поедая гусениц шелкопряда, муравьи могут приносить серьёзный вред данной отрасли. Использование муравьёв-ткачей при выращивании цитрусовых на юге Китая считается одним из старейших применений биологической борьбы с вредителями . В Южной Африке муравьи используются для сбора урожая ройбуша , имеющего маленькие семена, применяемые для производства травяного чая. Растение разбрасывает свои семена далеко, что усложняет их ручной сбор. Чёрные муравьи собирают и хранят их и другие семена в своём гнезде, откуда люди могут собирать семена. До 150—200 г семян можно собрать с одного муравейника .
С другой стороны, муравьи могут причинять неудобства, когда они вторгаются в жилища или наносят экономический ущерб . Так, трофобиоз муравей с тлями, червецами и некоторыми другими насекомыми , считающимися вредителями, носит негативное значение в оценках экономического значения муравьёв. Вред от разведения тлей в некоторой степени может компенсироваться тем, что падь является одним из важных источников дополнительного питания для хищных и паразитических насекомых.
Некоторые виды, например фараонов муравей , способствуют распространению бактериальных и вирусных заболеваний человека . Другие виды могут служить промежуточными хозяевами ряда паразитов : например, муравьи рода формика для ланцетовидной двуустки , вызывающей дикроцелиоз у скота .
В некоторых частях света (в основном в Африке и Южной Америке ) большие муравьи, особенно кочевые , используются при зашивании ран во время хирургических операций. Челюстями муравьёв захватывают края раны и фиксируют их на месте. Тело затем отрезают, а голова и челюсти остаются, способствуя сведению краёв раны .
Способность муравьёв эффективно использовать ресурсы нередко приводит к их конфликту с людьми, так как муравьи могут повреждать культурные растения (например, разводя на них тлей) или вторгаться в жилища человека.
Значение муравьёв в природе велико, однако некоторые виды питаются растениями, плодами и ягодами, наносят вред сельскому хозяйству , и считаются вредителями . В тропических странах плантациям могут вредить муравьи-листорезы .
Муравьи могут забираться в жилые и хозяйственные помещения, квартиры, где портят сладкие и мясные продукты. Также они могут иногда заползать в ульи и вредить пчеловодству . В плодовых садах они часто являются спутниками других вредителей — тлей , медяниц , червецов , цикад , выделяющих большие количества пади .
Таким образом, ряд видов могут считаться вредителями сельского и лесного хозяйства, а также бытовыми вредителями .
Из-за высокой приспособляемости муравьиных семей ликвидировать всю популяцию практически невозможно. В связи с этим управление численностью муравьёв-вредителей — это вопрос контроля местных популяций, а не уничтожения всех семей, и большинство попыток контроля являются временными решениями.
Среди муравьёв к вредителям причисляют дерновых муравьёв , жёлтых сумасшедших муравьёв , фараоновых муравьёв , муравьёв-древоточцев , в частности, Camponotus consobrinus , аргентинских муравьёв , Tapinoma sessile , красных огненных муравьёв , рыжих мирмик . Популяции муравьёв контролируются с помощью инсектицидов-приманок в жидком виде или в гранулах. Приманки собираются муравьями как продовольствие и доставляются в гнездо, где яд распространяется на других членов семьи через обмен пищей . Борная кислота и боракс часто применяются как инсектициды.
Один из видов муравьёв, являющийся вредителем — это мелкий, длиной 3 мм, жёлтого цвета, фараонов муравей , ставший космополитом, расселившимся по всему земному шару в процессе перевозки грузов. Кроме мясных и молочных продуктов, он может питаться хлебом, мучными изделиями, сахаром, повреждать энтомологические коллекции . Эти муравьи могут являться механическими переносчиками ряда инфекционных заболеваний .
Некоторые муравьи из подсемейств Myrmeciinae и Ponerinae имеют яд, опасный для человека. Среди них вид Paraponera clavata , род Dinoponera из Южной Америки , а также Myrmecia из Австралии . Смертельно опасными могут быть ужаления муравья Myrmecia pilosula . Красный огненный муравей может вызывать сильную аллергическую реакцию у высокочувствительных людей и анафилактический шок .
Хотя большинство видов муравьёв обладают стойкостью ко многим попыткам людей их уничтожить, некоторые виды находятся под угрозой снижения численности или вымирания. В основном это островные виды, развившие в ходе эволюции специализированные черты, включая Aneuretus simoni с Шри-Ланки и Adetomyrma с Мадагаскара . В Красный список угрожаемых видов МСОП занесены 149 видов муравьёв . Восточный лиометопум внесён в Красную книгу России (категория 2 — сокращающийся в численности вид) . К 2009 году в 15 субъектах Российской Федерации в региональные Красные книги (или в соответствующие нормативно-правовые акты об охране редких и исчезающих видов организмов) было включено 30 видов муравьёв . В отдельных странах Европы охраняют и расселяют гнёзда рыжих лесных муравьёв . С 1971 года в РСФСР проводилась Всероссийская операция «Муравей» по картированию, охране и расселению полезных видов лесных муравьёв рода Formica . В России уничтожение (разорение) муравейников является административным правонарушением, за которое предусмотрен штраф от 300 до 500 рублей или предупреждение .


Муравьи и их личинки съедобны и употребляются в пищу в различных частях света. «Яйца» (куколки, личинки) двух видов муравьёв из рода Лиометопум служат основой для приготовления блюда, известного в Мексике под названием « » . Их рассматривают как форму икры (или яиц насекомых) и продают по высокой цене, примерно по 90 долларов за кг, из-за того, что они являются сезонными и трудно обнаруживаемыми ресурсами. В Южной Америке (в департаменте Сантандер Колумбии ) собирают крупнотелых муравьёв, известных у местного населения как «hormigas culonas» ( муравьи-листорезы вида Atta laevigata ), которых жарят живыми и употребляют в пищу .
В различных частях Южной ( Индия ) и Юго-Восточной Азии ( Мьянма , Таиланд ) паста из зелёных муравьёв-ткачей ( Oecophylla smaragdina ) служит приправой к овощному (или мясному) блюду карри . Яйца и личинки муравьёв-ткачей , так же, как и их взрослые особи, используются при приготовлении тайского салата «yum» ( тайск. ยำ ), в блюдах под названием «yum khai mod daeng» ( тайск. ยำไข่มดแดง ) или салате из яиц красных муравьёв (блюдо происходит из региона Исан на северо-востоке Таиланда). Муравьёв-ткачей также используют в пищу аборигены Австралии (Северный Квинсленд) . Мексиканские индейцы едят рабочих муравьёв, известных как «медовые бочки» ( Myrmecocystus ) .
Завоеватель Колумбии Гонсало Хименес де Кесада в своём докладе «Краткое изложение завоевания Нового Королевства Гранада» ( 1539 , отредактировано анонимным автором в 1548—1549 годах) сообщил о традиции выращивания муравьёв (различных видов) для приготовления хлеба:
Эта земля плодородна и обеспечена пропитанием и едой большая её часть, потому что другая её часть менее изобильна, а ещё одна, намного меньше, и доходит до такой нужды в каком-либо месте у панчей, что когда их покорили, то натолкнулись на тех, кто обитал в земле Тунхи меж двух многоводных рек, в неких горах, на провинцию людей, не очень малочисленных, чьим пропитанием были одни только муравьи, и из них они готовили для еды хлеб, перемешивая их [в муку]. Коих муравьёв превеликое изобилие в самой провинции, и их выращивают для этой цели в загонах. И [эти] загоны представляют собой некие перегородки, построенные из широких листьев; и даже есть там, в той провинции, разнообразие муравьёв: одни — крупные, а другие — мелкие.
— Гонсало Хименес де Кесада. «Краткое изложение завоевания Нового Королевства Гранада»

Мирмекологи исследуют муравьёв как в лабораторных, так и в полевых условиях. Разнообразное поведение этих общественных насекомых делает их модельными организмами для научных экспериментов. Исследователи проверяют на муравьях разнообразные гипотезы в областях экологии и социобиологии , и они особенно важны в исследовании предсказаний теорий отбора сородичей ( Кин-отбор ) и эволюционно устойчивых стратегий развития . У муравьёв было впервые обнаружено восприятие ультрафиолетового света (John Lubbock, 1881) . Для исследования отдельных индивидуумов муравьёв, которых содержат в лабораторных муравейниках ( формикариях ), метят специальными разноцветными красками . С 1963 года в России (ранее — СССР) раз в 4 года проходят Всероссийские мирмекологические симпозиумы .
Несколько раз муравьи доставлялись на космические станции. Например, 12 января 2014 года по программе НАСА колония из 800 рабочих особей дерновых муравьёв , разделённая на 8 отсеков, была доставлена на МКС для исследования организации их поведения в условиях невесомости .

В шумеро-аккадской мифологии красные муравьи считались посланцами богини подземного мира Эрешкигаль , а сражение красных муравьёв в чьём-либо доме рассматривалось в числе предзнаменований .
По известному греческому мифу, Зевс превратил муравьёв острова Эгина в людей, сделав их царём своего сына Эака — так появилось племя мирмидонян , участвовавших в Троянской войне под предводительством Ахилла . По другому варианту, мирмидоняне произошли от героя Мирмидона , сына Зевса , который зачал его от Евримедусы, превратившись в муравья .
Автором обращённого к Индре гимна « Ригведы » (X. 99) считается Вамра, или Вамрака («муравей») из традиции Вайкханаса , а в его тексте (а также в гимне I. 51, 9) упоминается миф о превращении самого Индры в муравья, проникшего во вражескую крепость . В русской былине есть сходный сюжет: Волх Всеславьевич обращает своих дружинников в муравьёв, чтобы они смогли пробраться сквозь стены и ворота «индейского царства» . В «золотого муравья» превращается также юноша в осетинской сказке . В русской народной сказке «Хрустальная гора» Иван-царевич, обернувшись муравьём, проникает через маленькую трещину в хрустальную гору и освобождает царевну .
Немало упоминаний о муравьях разбросано в мифологии индейцев. Например, в мифе индейского народа, известного как « Черноногие », муравьи предстают в качестве изобретателей искусства вышивания по коже .
Геродот рассказывает, что огромные муравьи («величиной почти с собаку») в пустыне добывают золотой песок, который и забирают индийцы , эта легенда получила достаточно широкое распространение в античной и средневековой литературе, вплоть до « Фауста » Гёте . Упоминание о «золоте муравьёв», которым обладают цари, живущие по берегам реки Шайлоды, содержит и « Махабхарата » .
Ацтеки верили, что муравьи показали Кетцалькоатлю местонахождение маиса , а народы центрального Вьетнама рассказывали о муравье Хмоч Кенту, который принёс людям рис после всемирного потопа .
По индийской легенде, отражённой в стихотворении Леконта де Лиля «Смерть Валмики» , великий поэт Вальмики в старости впал в аскезу и был съеден муравьями (само имя valmīki означает «муравейник» и родственно имени Вамра, русскому, греческому и другим названиям муравья в индоевропейских языках).

Муравьи часто использовались в сказках, баснях и детских рассказах в качестве образца трудолюбия и взаимопомощи. Например, в басне Ивана Крылова « Стрекоза и Муравей » и басне Эзопа «Муравей и кузнечик» одним из главных героев является муравей. Муравьи также упоминаются в религиозных текстах, включая Библию ( Книга притчей Соломоновых ), Коран и другие .
Пойди к муравью, ленивец, посмотри на действия его, и будь мудрым. Нет у него ни начальника, ни приставника, ни повелителя; но он заготовляет летом хлеб свой, собирает во время жатвы пищу свою.
— Библия. «Книга притчей Соломоновых»
Муравьи — народ не сильный, но летом заготовляют пищу свою
— Библия. «Книга притчей Соломоновых»
В еврейском мидраше муравей обличает царя Соломона . На вопрос: есть ли на свете кто-либо более великий, чем он, Соломон, Муравей отвечает, что считает себя более великим, и царь не вправе слишком возноситься .
Также муравьи присутствуют в следующих литературных произведениях: Юрий Никитин — « Мегамир », « », Бернар Вербер — « Муравьи », « День муравья », «Революция муравьёв», Анна Старобинец — «Переходный возраст», Андре Моруа — «Муравьи», Клиффорд Саймак — « Город », сказки для детей Ондржея Секоры — « », «Приключения муравья Ферды», — «Муравей Красная Точка», Виктор Пелевин — « Жизнь насекомых », Валерий Медведев — « Баранкин, будь человеком! », Ян Ларри — « Необыкновенные приключения Карика и Вали ». Вымышленные гигантские доисторические муравьи присутствуют в фантастическом романе Владимира Обручева « Плутония » . Человеческое общество, организованное по принципу муравейника, описано в романе Фрэнка Герберта « Улей Хельстрома » («Hellstrom’s Hive», 1973) .
Приди ко мне, мой муравей,
Как рудокоп в туман приходит,
Лишь альтруисты в злате бродят,
Мой муравей…
Муравьи являются главными персонажами советских мультфильмов : « Путешествие муравья », снятого по мотивам сказки Виталия Бианки , « Слон и муравей », « Стрекоза и муравей », « Баранкин, будь человеком! » .
В качестве героев выступали муравьи в иностранных мультфильмах: « Муравей Антц », « Приключения Флика », « Гроза муравьёв ». Муравьи являются персонажами французского анимационного телесериала Minuscule , где они показаны целеустремлёнными и трудолюбивыми. Также муравьи являются эпизодическими персонажами мультипликационного флэш -сериала « Happy Tree Friends », где чаще всего они крайне жестоко расправляются с желающим полакомиться ими муравьедом «Ботаником» .
В кинематографе муравьи обычно появляются в фильмах ужасов , как наводящие страх существа, — « Они! », « Империя муравьёв », « Стеклянный муравейник », « Похититель костей », « Нашествие », « ». Кочевые муравьи из рода Dorylus под названием Siafu были показаны в фильме « Индиана Джонс и Королевство хрустального черепа » 2008 года. Они изображаются чрезмерно крупными по размеру, очень агрессивными и сильными, а также вне их естественной среды обитания: они не встречаются в Перу или Бразилии , где происходит действие фильма.
Муравей является символом музыкальной группы The Prodigy . Также у рок-группы Stone Temple Pilots имеется песня «Army Ants» (с англ. — «кочевые муравьи»), входящая в состав музыкального альбома « Purple ». В названиях британской группы Adam and the Ants («Адам и муравьи») и некоторых её песен обозначены муравьи (сингл « » был № 2 в январе 1981 года в британском чарте ).
Видеоклип немецкой музыкальной группы Rammstein на песню « Links 2-3-4 » рассказывает о сражении населения муравейника с жуками и был снят как при помощи натуральных съёмок, так и компьютерной анимации . У группы Аквариум в альбоме « Треугольник » есть композиция «Мой муравей» .



Муравьи стали символом творческого порыва Сальвадора Дали при написании его картин, а впоследствии и символом высоких духовных отношений и любви Дали и Галы . Сам он говорил, что однажды к нему во сне пришёл гигантский муравей с человеческим ртом и говорил с ним голосом Галы, которую он повстречал через несколько дней. На нескольких картинах Дали повсеместно встречаются муравьи .
В геральдике муравей, как и пчела , служит эмблемой трудолюбия и покорности .
Муравьи изображены на аверсе памятной монеты из медно-никелевого сплава Банка Латвии достоинством 1 лат , выпущенной в 2003 году .
Муравьи также изображены на лицевой стороне серебряных монет Боспорского царства . Существует предположение, что они были выпущены в античном городе Мирмекии , основанном греками в середине VI века до н. э. на берегу Керченского пролива .
|
|
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
|
Семейства жалящих перепончатокрылых (
Aculeata
)
|
||
|---|---|---|
|
||
|
Современные
пчёлы, осы и муравьи |
|

|
| † Вымершие | ||
|
|
||
|---|---|---|
|
||
| Современные |

|
|
| † Вымершие | ||
| Бывшие | ||
|
|
Эта статья входит в число
избранных статей
русскоязычного раздела Википедии.
|
|
|
Эта статья победила на конкурсе
статьи года
и была признана статьёй 2011 года русской Википедии.
|