Полужесткокрылые
- 1 year ago
- 0
- 0

Жесткокры́лые , или жуки́ ( лат. Coleoptera ) , — отряд насекомых , представители которого характеризуются видоизменением передних крыльев в твёрдые, сильно склеротизированные, либо кожистые надкрылья , лишённые жилкования , с сохранением перепончатых задних (нижних) крыльев , служащих для полёта и в спокойном состоянии сложенных под надкрыльями (очень редко крылья и надкрылья редуцированы) . Для представителей отряда характерны грызущий и жующий ротовой аппарат, развитая передняя часть груди, подвижное сочленение переднегруди с её средней частью . Форма и размеры тела весьма разнообразны: от 0,3—1,0 мм до 171 мм .
Развитие с полным превращением : имеются стадии яйца , личинки , куколки и имаго . Личинки с хорошо развитой хитинизированной головой и грызущим ротовым аппаратом, преимущественно червеобразные или камподеевидные, у большинства видов с членистыми грудными конечностями, без брюшных ног. Куколки преимущественно свободные, обычно мягкие, неподвижные (двигаться может только брюшко), неокрашенные .
Жуки являются крупнейшей группой среди насекомых и живых существ в целом. По состоянию на август 2013 года в отряде насчитывается 392 415 видов , включая 2928 вымерших видов , что составляет 40 % от всех известных видов насекомых . По данным на 2018 год, на территории России обитает 14 178 видов жесткокрылых из 155 семейств .
Раздел энтомологии , изучающий жесткокрылых, называется колеоптерологией .
Латинское название отряда «Coleoptera» происходит от др.-греч. κολεός — «ножны» и πτερόν — «крыло» .
Своё название — «жесткокрылые» , представители отряда получили в связи с видоизменением передней пары крыльев в хитинизированные твёрдые надкрылья .
Русское «жук» восходит к праслав. žukъ , образованному от той же звукоподражательной основы, что и слово «жужжать» .
По количеству описанных видов жуки являются наиболее многочисленной и разнообразной группой организмов на Земле . По состоянию на август 2013 года учёными описано 392 415 видов жуков, включая 2 928 ископаемых видов . Жесткокрылые очень разнообразны и большинство их видов плохо изучено, поэтому истинная оценка количества существующих видов является очень затруднительной. Некоторые из описанных видов известны только по находкам из одной единственной местности или даже по единственному экземпляру . Первая категоризация по системе Линнея предпринята в начале XIX века К. Г. Яблонским .
Жуки широко распространены по всему земному шару , в шести зоогеографических областях , кроме Антарктиды , Арктики и наиболее высоких горных вершин. Наиболее богато видами отряд представлен в тропических регионах .
Примерно 62 % всего разнообразия жуков составляют представители всего шести семейств : долгоносики (Curculionidae) — 60 000 видов , стафилиниды (Staphylinidae) — 47 700 , жужелицы (Carabidae) — 30 000 , пластинчатоусые (Scarabaeidae) — 27 800 , листоеды (Chrysomelidae) — 36 350 , и усачи (Cerambycidae) — 26 000 .

Размеры представителей группы колеблются в широких пределах. Мельчайшими в мире жуками являются перокрылки из трибы Nanosellini , которые имеют длину менее 1 мм; среди них самый маленький жук — Scydosella musawasensis (от 0,325 мм до 0,352 мм) , а также Vitusella fijiensis и Nanosella fungi . Это одни из мельчайших представителей непаразитических видов насекомых .
Одними из крупнейших жуков в мире считаются южноамериканские виды: дровосек-титан ( Titanus giganteus ), с длиной тела до 167 мм , а по некоторым неподтверждённым источникам даже до 210 мм ; жук-геркулес ( Dynastes hercules ), отдельные особи самцов которого достигают длины до 171 мм и крупнозуб оленерогий ( Macrodontia cervicornis ) с максимальной длиной самцов 170 мм (а по некоторым данным до 178 мм ). За ними следует вид Dynastes neptunus , с максимальной длиной самцов 158 мм и усач Xixuthrus heros с Фиджи , достигающий длины до 150 мм .
Крупнейшим видом жуком, обитающим на территории Европы , является жук-олень ( Lucanus cervus ), отдельные особи самцов номинативного подвида которого могут достигать длины 83—86 мм при средней длине самцов 70—74 мм. Крупнейшим жуком, обитающим на территории России , является реликтовый дровосек ( Callipogon relictus ), достигающий длины 110 мм .
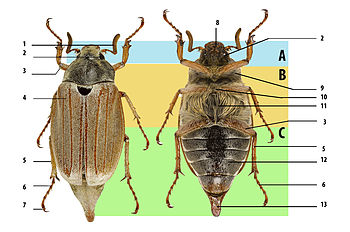

Тело взрослых жуков, или имаго , как и всех других насекомых, состоит из трёх основных отделов: головы, груди и брюшка . Придатки первых двух отделов обычно выступают наружу, тогда как придатки брюшка (прегенитальные и генитальные) чаще всего в той или иной мере втянуты внутрь.
Тело преимущественно овальное, удлинённо-овальное или удлинённое, более или менее выпуклое сверху, характеризуется сильно склеротизованными покровами и их придатками . Прочный экзоскелет обеспечивает пассивную защиту, преимущественно от воздействий субстрата при перемещении жуков внутри него .
О связи образа жизни и формы тела жуков можно судить преимущественно по крупным видам, в меньшей степени — по жукам средних размеров, а у мелких видов такая взаимосвязь часто не обнаруживается вообще . Например, обитатели водной среды — активно плавающие формы ( плавунцы , вертячки и т. д.) всегда характеризуются обтекаемой и компактной формой тела. Открыто живущие формы часто обладают сильно выпуклым телом. Передвигающиеся по поверхности субстрата хищные виды характеризуются довольно компактным телом с хорошо развитыми и сравнительно длинными конечностями ( жужелицы ). Роющие формы ( скарабеи , навозники ) обладают расширенным передним краем тела и более мощной переднеспинкой по сравнению с открыто живущими видами .
Голова состоит из собственно головной капсулы (которая в ходе эволюции образовалась путём слияния нескольких сегментов тела и и имеет округлую форму) и придатков — ротовых органов и усиков . В зависимости от положения головы жуков, она может быть прогнатической , когда ротовые части направлены вперёд (в большинстве случаев); гипогнатической (ортогнатической), когда ротовые части направлены вниз и опистогнатической (отклонены назад) . У представителей некоторых семейств ( долгоносики , трубковёрты , ложнослоники ) голова вытянута в головотрубку , на вершине которой находится ротовой аппарат .


Голова покрыта цельной капсулой. Своей задней частью, как правило, входит в переднегрудь . Иногда заходит в неё так глубоко, что сверху практически незаметна; порой, наоборот, свободно прикреплена к переднегруди .
На верхней стороне головы различают несколько слабо разделённых частей: шею , затылок , виски , темя , лоб и наличник . Головная капсула умеренно уплощена в дорсовентральном направлении и обычно отчасти втянута в переднегрудной склерит . Особенностью жуков является значительное удлинение нижней поверхности головной капсулы за ротовыми частями.
Голова несёт органы чувств : органы обоняния, осязания (усики и щупики ) и зрения (сложные, реже — простые, глаза). По бокам головы, размещены сложные фасеточные глаза (число фасеток может достигать 25 000 ). Обычно они хорошо развиты, часто довольно крупные. В редких случаях могут быть редуцированы (у пещерных, подземных и паразитических форм вплоть до полного исчезновения) . Иногда глаза могут быть смещены на дорсальную сторону, а у вертячек , ( Hydrophilidae ), Lamiinae и других разделены боковым краем, из-за чего каждый глаз состоит из одной половинки, смотрящей вверх, и другой, смотрящей вниз. Простые глазки у большинства видов отсутствуют. У некоторых семейств жуков они располагаются на темени ( Hydraenidae , , стафилиниды , кожееды , Phloeostichidae ), при этом непарные простые глаза у кожеедов и Metopsia имеют парное происхождение .
Между глазами, выше затылка находится темя , которое в передней части переходит в лоб . На лбу, между глазами, расположены усики (антенны) — членистые придатки, которые выполняют функции органов осязания и обоняния. Их основания находятся в усиковых впадинах. Обычно усики 11-члениковые, но их число может уменьшаться путём слияния до 2 ( Paussus ), либо увеличиваться до 40 путём деления отдельных члеников . Строение и форма усиков разнообразны и сильно варьируют у представителей различных семейств .
Усиковые ямки расположены между основанием мандибул и передним краем глаза и являются особой чертой, которая отличает жуков от многих насекомых с полным метаморфозом . У многих разноядных жуков и ряда жужелиц на нижней поверхности эпикраниума находятся усиковые бороздки, в которые вкладываются первые членики усиков .
Ротовые придатки не опущены в ротовую капсулу и состоят из непарной верхней губы ( лабрум ), пары нечленистых верхних челюстей ( мандибулы ), пары нижних челюстей ( максиллы ) и непарной нижней губы ( лабиум ), которая образовалась в результате слияния второй пары нижних челюстей. Нижние челюсти и нижняя губа несут органы осязания и вкуса — щупики . Для жуков характерен наиболее примитивный, грызущий ротовой аппарат .
Ротовые органы жуков выполняют функцию измельчения твёрдой пищи. Верхние челюсти имеют жевательный край с развитыми зубцами. Зубцы левой челюсти входят в углубления правой. Максиллы заканчиваются подвижно причленёнными жевательными лопастями — внешней и внутренней. Нижняя губа имеет основной и дистальный членик, на котором находятся две пары жевательных лопастей .
Грудь состоит из трёх сегментов. Из них наиболее развита переднегрудь , подвижно сочленённая со среднегрудью , которая неподвижно сочленена с заднегрудью , а та — с брюшком .
С дорсальной стороны эти сегменты называются спинками: передне-, средне- и заднеспинкой. Кутикула каждого сегмента представляет собой кольцо, которое делится на четыре склерита: спинной — тергит, грудной — стернит и два боковые — плейриты . Грудные сегменты несут на себе органы движения: средне- и заднеспинка — крылья , все нижние полусегменты — по паре ног .
Переднеспинка очень разнообразна по форме и своей скульптуре, а её строение имеет важное значение при определении жуков. Она может иметь боковые шипы, разнообразные выросты либо горбики .
Сверху видна только переднеспинка , а среднеспинка, кроме щитка, и заднеспинка прикрыты надкрыльями . Надкрылья крепятся к среднеспинке, а к заднеспинке — перепончатые крылья , используемые при полёте. Часть надкрылий, которая заходит на нижнюю сторону тела, называется эпиплеврами. С нижней стороны грудь состоит из передне-, средне- и заднегруди. Каждый из этих сегментов, в свою очередь, состоит из центральной части — диска, отделённых от него боковой ( эпистерн ) и задней (эпимер) сторон. В задней части каждого сегмента груди находятся тазиковые впадины, в которых расположены, собственно, тазики — часть конечности .
Ноги у жесткокрылых всегда хорошо развиты. Каждая из них состоит из пяти основных частей: тазиков, вертлугов, бёдер , голеней и . Тазики помещаются в тазиковых впадинах и представляют собой короткий базальный членик, с помощью которого нога подвижно прикрепляется к плейриту , они могут иметь шаровидную, конусовидную либо поперечно-продольную форму.
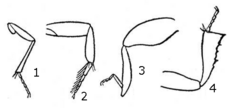

Передние и средние тазики всегда подвижные, в то время как задние обычно подвижны лишь в ограниченной мере, а иногда (у жужелиц , златок , плавунцов , щелкунов ) неподвижны и сочленены с заднегрудью . Тазик с бедром соединяет вертлуг. Суставы между тазиком и грудью и тазиком и вертлугом движутся в разных плоскостях, обеспечивая подвижность ноги. Бедро — самый сильный и, как правило, крупнейший членик конечности жука. К вершине бедра присоединяется голень, а к ней лапка с члениками. На вершине голени расположены шпоры, обычно две, а у некоторых видов по одной. Лапка состоит преимущественно из пяти члеников, хотя у разных семейств их количество варьирует, что является одним из систематических признаков. Число этих члеников может сокращаться до 4—3, очень редко до 2—1, и крайне редко их может совсем не быть (на передних ногах Scarabaeus , Onitis ) . Нижняя поверхность лапки может быть покрыта густыми волосками, щетинками. Последний членик лапки, у всех жуков, несёт два . У некоторых жуков между коготками имеется эмподий — подвижный кожистый придаток с 2 щетинками. Коготки лапки могут сильно варьировать по длине и форме. Остальные членики лапки могут быть сильно модифицированы: первый членик часто может сильно удлиняться (преимущественно на задних ногах); второй и третий могут быть сильно расширенными и вырезанными, двухлопастными, а четвёртый часто бывает недоразвитым .
В зависимости от способа жизни у жуков различают различные типы ног: бегательные, ходильные, копательные, прыгательные, плавательные, хватательные и т. д. Ходильные ноги отличаются строением лапки, которая является расширенной и приплюснутой, её подошва густо усажена волосками ( усачи , листоеды , долгоносики , и др.), 2-й или 3-й членики лапки обычно расширены. Бегательные ноги характеризуются стройностью и пропорциональность всех частей, довольно тонкой 5-члениковой лапкой ( жужелицы , чернотелки ). Копательные ноги (преимущественно бывают передними) отличаются расширенной и уплощённой голенью, наружный передний край которой снабжён зубцами ( пластинчатоусые , карапузики и др.). Плавательная нога отличается расширенной голенью и лапкой, края которых покрыты крепкими густыми волосками (у водяных жуков). Прыгательными принято называть задние ноги, обладающие мощным, очень утолщённым бедром, которое служит для совершения прыжков (некоторые листоеды, долгоносики и др.) . Хватательные ноги (только передняя пара) встречаются довольно редко (у Gyrinidae , самцов Labidostomis ), являются тонкими и сильно удлинёнными и используются для удержания добычи или самок во время спаривания .

У жуков передние крылья превратились в твёрдые склеротизованные надкрылья . Они обычно имеют такую же твёрдость, как и хитиновые покровы тела. Они крепятся на среднегруди. В покое надкрылья прикрывают собой среднеспинку (исключая щиток), заднеспинку и верхнюю сторону брюшка. Иногда они могут быть укороченными и не покрывают от 1 до 7 тергитов брюшка. Иногда надкрылья сильно недоразвиты, а у самок некоторых видов ( , и др.) могут совсем отсутствовать. Но порой, наоборот, надкрылья превосходят своей длиной брюшко. Линия соприкосновения надкрылий называется швом. У жуков с редуцированными или отсутствующими задними крыльями надкрылья могут срастаться. Иногда надкрылья, наоборот, не соприкасаются по шву (у некоторых веероносцев , нарывников , узкокрылок , сверлил и др.) . Крайне редко ( нарывники Meloe ) одно из надкрылий может немного налегать на другое в передней половине, а концы их при этом расходятся. Наружный край надкрыльев на всём своём протяжении часто подогнут на нижнюю сторону и частично прикрывает боковые части средне- и заднегруди и брюшка, образуя эпиплевры. У некоторых семейств ( жужелицы , темнотелки , щитоноски и др.) боковой край надкрылий может быть сильно уплощённым и выступающим, порой торчащим или отогнутым кверху . У жуков из нескольких родов короедов на вершине надкрылий имеется вдавление — т. н. «тачка», окружённая по краям зубцами и используемая для выбрасывания из ходов, выгрызаемых в древесине, буровой муки.
Скульптура надкрылий может быть «полированной гладкой», нести на себе разнообразные выросты, ямки, шипы. Часто на надкрыльях имеются продольные бороздки или ряды углублённых точек, отделённые друг от друга промежутками . Благодаря сильной склеротизации надкрылий и развития на их верхней поверхности различных скульптурных образований исходное жилкование надкрылий полностью неразличимо и скрыто у подавляющего большинства жесткокрылых. Только у ряда жуков с относительно мягкими покровами ( краснокрылы , род ) и у представителей примитивного семейства Cupedidae на верхней поверхности надкрылий выделяется первичное жилкование в виде продольных полос .

Крылья крепятся на заднегруди между тергитом и плейритом посредством сложной системы склеритов и мембран. Рядом с местом прикрепления крыло опирается на вырост плейрита, который является для него точкой опоры и образует рычаг с коротким осевым и длинным конечным плечами .
Задние (нижние) крылья перепончатые, в большей или меньшей степени прозрачные, слегка окрашенные либо бесцветные, с твёрдыми, более тёмно окрашенными жилками. Крылья могут быть редуцированы или вообще отсутствовать. Обычно крылья длиннее (порой значительно) и шире надкрылий, в состоянии покоя находятся скрытыми под надкрыльями (исключение составляют неполнокрылы , короткокрылы и немногие другие). При этом крылья перегибаются в продольном и поперечном направлениях .
У жуков различают три основных типа жилкования крыльев: карабоидный (1—2 поперечные жилки между ветвями медиальной жилки, образующие замкнутую ячейку), стафилиноидный (полное отсутствие поперечных ячеек и недоразвитие основания ветви М 1 ), кантароидный (наиболее распространённый) .
Брюшко состоит из сегментов. Каждый сегмент брюшка состоит из полуколец: спинного ( тергит ) и брюшного ( стернит ), соединённых плевральной перепонкой, на которой находится дыхальце . Иногда дыхальца располагаются по бокам тергитов брюшка. Обычно они скрыты под надкрыльями .

Количество сегментов непостоянно — их бывает не более 10; видимых сегментов обычно 7—9, стернитов 5—8 (так как стерниты первых 2 сегментов очень часто отсутствуют — редуцируются ), а 1—3 последние сегменты видоизменяются и преимущественно впячиваются внутрь брюшка . Сегменты брюшка с нижней стороны называются стернитами , а с верхней — тергитами . Тергиты обычно слабо хитинизованы , перепончатые, почти не окрашены, а выступающие из-под надкрылий (если они укорочены) являются такими же твёрдыми и окрашенными, как стерниты ( стафилиниды , некоторые дровосеки ). Последний из тергитов называется пигидием , а предпоследний — пронигидием . У некоторых видов самцы за пигидием имеют ещё и постпигидий (у Nitidulidae , Chrysomelidae , Pythidae и др.). Последние сегменты видоизменены, обычно находятся внутри брюшка .
Наружные гениталии самцов в спокойном состоянии находятся в генитальной камере и снаружи обычно не видны. Они выдвигаются из конца брюшка только во время спаривания. Мужской копулятивный орган ( эдеагус ) подвижно крепится на модифицированных IX и X сегментах брюшка. В типичных случаях он состоит из пениса, двух парамер и базального склерита. Внутри пениса у многих жуков находится эндофаллус , выворачивающийся во время спаривания и вводящий в половые пути самки сперматофор . Наружные гениталии самок образованы последними члениками брюшка и состоят из , представляющего собой полустернит X стернита брюшка .
Окраска тела представителей отряда является очень изменчивой и разнообразной. Окраска и рисунки на теле часто являются характерными для отдельных видов или различных таксономических групп .

У жуков различают структурную и пигментную окраску. Структурная окраска связана с некоторыми специфическими особенностями поверхностной структуры кутикулы (микроскопическими рёбрышками, пластинками, чешуйками и т. п.), которые создают эффекты интерференции , дифракции и дисперсии света . Это металлически блестящие оттенки некоторых жуков, в том числе многих тропических видов. Пигментная окраска обусловлена пигментами , которые часто содержатся в экзокутикуле, реже в клетках гиподермы или жирового тела . У многих жуков наблюдается сочетание структурной и пигментной окраски. Наиболее распространёнными пигментами жуков являются меланины , дающие тёмно-коричневую, коричнево-красную или чёрную окраску. А также каротиноиды , обеспечивающие жёлтый, оранжевый, красный окрас; флавоноиды , ответственные за жёлтый, белый, красный и коричневый цвета. Некоторые пигменты (особенно чёрные и бурые) отличаются большой стойкостью, а красные и зелёные неметаллические цвета обычно являются менее стойкими и после гибели жуков часто изменяют свою окраску на жёлтую, рыжую, буро-жёлтую .

Многие виды жуков обладают разнообразными, часто весьма яркими рисунками на различных частях своего тела. Преимущественно ярко окрашены жуки с дневной активностью, приуроченные к открыто расположенным растительным субстратам (цветки, листья), например бронзовки и златки , ряд усачей .
Для многих видов, перемещающихся по поверхности почвы или обладающих ночной активностью и многих обитающих в воде жуков характерна относительно однотонная и преимущественно тёмная окраска, хотя порой она сочетается с выраженными металлическими оттенками ( жужелицы и чернотелки ). Жуки мелких и средних размеров, а также виды, ведущие более или менее скрытый образ жизни, преимущественно обладают одноцветной окраской. Виды, обитающие в пещерах , либо не выходящие на поверхность почвы, обычно характеризуются бледно-коричневатой окраской тела. В некоторых группах жуков яркая окраска, либо броские рисунки на теле служат предупреждением для хищников о ядовитости (например, нарывники и божьи коровки ) .
Окраска может быть одноцветной, либо состоящей из комбинации нескольких цветов, преимущественно 2—3, реже — большего числа цветов. На общем фоне могут иметься пятна или другой рисунок иного цвета. У одного и того же вида окраска может порой сильно варьировать: например, в ряде случаев металлические тона могут быть представлены всевозможными оттенками от медно-красного до чёрно-синего. Пятна и полосы могут полностью исчезать или расширяться, сливаясь друг с другом .
Только что вышедшие из куколки жуки обладают бледной, более светлой окраской наружных покровов.
Поверхность тела может быть гладкой, почти полированной, либо наоборот — неровной и шероховатой. Часто тело многих жуков может быть покрыто пыльцой или налётом, которые обычно легко стираются. Они могут быть настолько густыми, что полностью скрывают фоновую окраску .
Половой диморфизм — различия во внешнем виде, окраске и/или форме тела самца и самки, является широко распространённым и порой резко выраженным явлением среди различных видов жесткокрылых. Самцы преимущественно несколько мельче и обладают более узким телом, чем самки. В других случаях, наоборот, самцы крупнее (за счёт сильно развитых мандибул или выростов-рогов на голове и переднеспинке: рогачи (Lucanidae), дупляки (Dynastinae), кравчики (Lethrus), Labidostomis ).
Очень часто (у большинства жужелиц и плавунцов , многих чернотелок , листоедов , некоторых хрущей и др.) передние, а порой и средние лапки самца сильно расширены и несут подошву из присасывательных волосков, служащую для удерживания самки во время спаривании. Порой самцы обладают сильно удлинёнными передними ногами (подсемейство Euchirinae , усач Acrocinus longimanus ). У других видов самцы характеризуются раздутыми задними бёдрами или искривлёнными, а часто и удлинёнными передними, средними или задними голенями .

Порой у самцов сильно развиты мандибулы: у рогачей ( Lucanidae ) и кравчиков ( Lethrus ). У представителей некоторых семейств, например у пластинчатоусых , голова и/или переднеспинка самцов несёт разнообразные «рога», выросты, бугорки, отсутствующие или слабо развитые у самок. Вторично-половые признаки могут также выражаться в виде модификации переднеспинки, переднегруди, ног и пигидия. Дупляки характеризуются очень резко выраженным половым диморфизмом. Самцы представителей трибы , как правило, значительно крупнее самок, с более или менее сильно развитыми рогами на голове и/или переднеспинке. Рог на голове у них нередко достигает очень значительной длины (род Dynastes , Megasoma и др.), снабжён зубцами, а у самок на голове вместо рогов обычно имеются лишь бугорки. Также у данных самцов передняя часть переднеспинки часто несёт рога, направленные вперёд .
Половой диморфизм также может проявляться в длине усиков ( усачи ), размерах и развитии их булавы (больше у самцов, чем у самок хрущей ), в отдельных случаях у самцов больше члеников на усиках, чем у самок.
Менее подвижный образ жизни самок порой обусловливает частое отсутствие у них крыльев (бескрылые самки светляков , ряда хрущей , некоторых дровосеков и др.), тогда как самцы данных видов всегда крылатые. Иногда и надкрылья самок могут быть недоразвиты или совсем отсутствовать (некоторые светляки). В крайне редких случаях (у некоторых короедов ), наоборот, самки всегда имеют крылья, тогда как у самцов крылья часто редуцированы .

Полиморфизм — появление различных форм, отличающихся по ряду признаков, внутри одного вида. Возникновение т. н. трофического полиморфизма у самцов жуков из семейства рогачей и подсемейства дупляков , связано с условиями развития личинки, количеством доступной кормовой базы, а также условиями окружающей среды . Более крупные особи представителей данных групп отличаются и наиболее сильно развитыми выростами и «рогами» на голове и/или переднеспинке самцов. При этом маленькие самцы отличаются, кроме слабого развития вторичных половых признаков, также некоторыми чертами, сближающими их внешне с самками . Трудно установить, что в данном случае имеет отношение к условиям, в которых развивались личинки, а что может быть объяснено генетически .
Также у ряда жуков, преимущественно с яркой «металлической» окраской (некоторые бронзовки, златки, усачи), могут возникать различные цветовые вариации внутри одного вида, при этом металлические тона их окраски могут быть представлены всевозможными оттенками, варьирующими от зелёных оттенков до медно-красного и чёрно-синего. У жуков-голиафов видов Goliathus orientalis и особенно Goliathus goliatus отмечаются многочисленные формы вариации окраски, которые порой резко отличаются друг от друга, либо являются очень сходными, или образующими ряд переходных форм . Тёмные вариации окраски преобладают у жуков из Центральной и экваториальной Африки , в то время как светлые вариации распространены у жуков из южных и западных районов экваториальной Африки. Изменчивость окраски Goliathus goliatus может быть результатом изменения целого ряда абиогенных факторов (степень инсоляции, влажность и т. п.) .
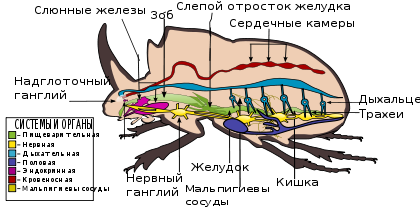
Внутреннее строение жесткокрылых является типичным для всех представителей класса насекомых . Полость тела жуков, как и всех насекомых и членистоногих в целом, представлена смешанной полостью тела, называемой миксоцелем . Она разделена двумя продольными горизонтальными перепонками ( диафрагмами ) на три отдела ( ). Верхняя диафрагма отделяет верхний, или перикардиальный синус, в котором расположен спинной кровеносный сосуд. Нижняя диафрагма отделяет нижний, или перинейральный синус, где содержится брюшная нервная цепочка. Средний синус лежит между диафрагмами; он называется висцеральным; в нём содержатся пищеварительная, выделительная и половая системы, а также большая часть жирового тела. Полость тела заполнена гемолимфой .

Центральная нервная система жуков, как и других насекомых, состоит из парного надглоточного ганглия , или головного мозга, окологлоточных коннектив и брюшной нервной цепочки. Первый ганглий цепочки (подглоточный) находится, как и надглоточный, в голове, остальные располагаются в туловище. Степень централизации брюшной нервной цепочки варьирует в широких пределах. Так, у краснокрыла Dictyopterus три грудных и восемь брюшных ганглиев; у жука-скакуна Cicindella — 3 грудных и 6 брюшных, у божьей коровки Coccinella septempunctata — 3 грудных и 2 брюшных, у Coccinella quinquepunctata — 3 грудных и 1 брюшной, у долгоносиков (Curculionidae) — 2 грудных и 2 брюшных, у вертячки Gyrinus — 2 грудных и 1 брюшной, а у скарабеиды Rhizotrogus — 1 грудной и 0 брюшных (все ганглии брюшной цепочки сливаются в один, расположенный в грудных сегментах) . У жуков хорошо развита вегетативная нервная система , состоящая из трёх отделов: стомато-гастрального, вентрального или брюшного и каудального .
Пищеварительная система жуков состоит из трёх отделов: передней, средней и задней кишки. Стенки всех отделов кишечника образованы однослойным эпителием , который снаружи покрыт продольными и кольцевыми мышечными волокнами. Их сокращение обеспечивает движение пищи в кишечнике. У жуков имеются мандибулярные, максиллярные и лабиальные (нижнегубные) железы. Глотка и пищевод обеспечивают проглатывание пищи и её прохождение в зоб , являющийся местом накопления пищи и начального её переваривания под действием ферментов слюны и пищеварительного сока из средней кишки. Задняя кишка не участвует в пищеварении, а её функции связаны с формированием экскрементов , водным обменом, выделением и осморегуляцией .

Кровеносная система крайне редуцирована. Представлена спинным сосудом , расположенным в перикардиальном синусе и подвешенным с помощью соединительнотканных тяжей к дорсальной стенке тела. Задняя его часть представлена сердцем , состоящим из ряда последовательных камер, а передняя представлена аортой .

Дыхательная система жуков представлена трахеями, которые открываются наружу несколькими парами дыхалец — стигм. Две пары стигм расположены соответственно на средне- и заднегруди, и первых пяти сегментах брюшка .
Выделительная система представлена мальпигиевыми сосудами , уратными клетками жирового тела , перикардиальными клетками .
Половая система самки состоит из пары яичников , пары яйцеводов , непарного яйцевода, придаточных половых желёз, семяприёмника. Половая система самца состоит из пары семенников , пары семяпроводов, семяпроводящего канала, придаточных половых желёз и копулятивного органа ( эдеагуса ) .
Жуки имеют хорошо развитые органы чувств: механорецепторы (воспринимающие прикосновение, вибрацию и звуковые волны); терморецепторы ; глаза . Глаза у жуков сложные ( фасеточные ), в состав которых входит большое количество, иногда несколько тысяч оматидиев, плотно прилегающих друг к другу. Каждый оматидий воспринимает только одну точку предмета находящегося перед ним, в результате чего, фасеточный глаз даёт мозаичное изображение, состоящее из множества отдельных точек. Также имеются проприорецепторы , которые сигнализируют нервной системе о положении, деформации и перемещении отдельных участков тела.
Звуковые колебания жуками воспринимаются при помощи Джонстоновых органов, которые состоят из модифицированных хордотональных сенсилл .
Тимпанальные органы слуха достоверно описаны у представителей двух семейств: жуков-скакунов рода Cicindela (на дорсальной поверхности первого абдоминального сегмента) и у представителей двух триб подсемейства Dynastinae (под щитом переднеспинки) .

Яйца жуков обычно овальной, круглой или продолговатой формы. Преимущественно светлоокрашенные (белого, желтоватого, серовато-жёлтого, серого, зеленоватого или светло-зелёного цвета, реже коричневато-буроватые), полупрозрачные. Их оболочка обычно тонкая либо кожистая, без чёткой наружной скульптуры .
Откладываются яйца преимущественно поодиночке либо небольшими кучками, чаще всего в укромные места: под различные укрытия, под камни, опавшую листву, мох , в стебли, почки , плоды растений, трещины коры деревьев, в землю. Порой для откладывания яиц самки подготавливают специальные полости (например, подземные норы навозников , заполненные навозом ; маточные ходы короедов ) или свёртывают определённым образом листья в виде сигар ( трубковёрты ) , в редких случаях, у хищных форм, самки откладывают яйца в гнёзда других насекомых (например, представители рода Trichodes откладывают яйца в гнёзда одиночных пчёл и ос, кубышки саранчовых ) .


Личинки жуков весьма разнообразны, однако обладают общими чертами строения: характеризуются развитой, сильно склеротизированной головой с грызущим ротовым аппаратом, преимущественно удлинённым мясистым телом с мягкими или склеротизированными покровами .
У скрытно живущих личинок окраска обычно белая или желтоватая, редко розовая, а у открыто живущих видов — тёмная: чёрная, серая, бурая, грязно-зелёная (личинки некоторых листоедов, питающиеся открыто на листьях); иногда на основном фоне могут находиться красные, жёлтые, бурые или металлически блестящие пятна.
По особенностям строения принято выделять несколько основных типов личинок жуков .
Личинки водных видов жуков принято разделять на 2 типа в зависимости от их образа жизни. Одни из них являются хищными ( Dytiscidae , Hydrophilidae ), характеризуются большой головой, острыми серповидными челюстями, довольно длинными ногами, порой покрытыми густыми волосками, с помощью которых быстро плавают. Они дышат атмосферным воздухом, для чего вынуждены периодически подниматься к поверхности воды. У других, в основном нехищных форм ( Gyrinidae , Haliplidae и многие водолюбы ), на теле имеются трахейные жабры (своеобразные выросты), используемые для дыхания под водой. Личинки данного типа гораздо менее подвижны, но среди них также встречаются хищные формы .
Существуют также и другие типы личинок: например, сильно волосатые личинки кожеедов , часто с кисточками длинных волосков на конце брюшка, или личинки, строение которых несколько отличается от основных типов ( Cicindelinae , Lymexylidae и некоторые другие) .

Куколка белая, свободная. Окукливание происходит преимущественно в почве, либо в лесной подстилке, даже если развитие личинки происходило в прочих местах: в древесине деревьев или под корой, внутри стеблей травянистых растений, в «сигарах» или пакетах, свёрнутых из листьев, и т. п. Личинки большинства водных видов также окукливаются в почве по берегам водоёмов. Однако многие личинки окукливаются в местах своего развития: в древесине деревьев (усачи, златки , короеды и др.), на листьях кормовых растений в минах ( долгоносики ), в пакетах из листьев (некоторые трубковёрты), либо открыто, непосредственно на листьях ( божьи коровки , некоторые листоеды ), внутри зерна ( зерновки ), в гнёздах перепончатокрылых (многие Cleridae ) .
Перед процессом окукливания личинки обычно устраивают особую «куколочную колыбельку», крайне редко (некоторые долгоносики) плетут настоящий кокон на поверхности листьев .
Во время фазы куколки происходят интенсивные процессы внутренней перестройки: гистолиз и гистогенез . Органы взрослого насекомого формируются из особых зачатков ( имагинальных дисков ). Это небольшие скопления недифференцированных клеток, расположенные в определённых местах тела личинки. Метаморфозы регулирует нейроэндокринный комплекс органов .

Жесткокрылые всегда являются раздельнополыми и почти всегда яйцекладущими . Жуки относятся к насекомым с полным превращением . Это означает, что из яйца появляется личинка, не похожая на взрослого жука, которая в конце своего роста превращается сперва в неподвижную куколку, а затем в имаго (взрослое насекомое). Таким образом, преобладающее большинство видов жуков проходят в своём развитии 4 фазы: яйцо, личинка, куколка и имаго . Однако имеются некоторые исключения — в некоторых семействах (например, нарывников ) наблюдается явление гиперметаморфоза с 5—6 и более фазами, которые представлены в виде дополнительных, особых личиночных стадий, непохожих на основную личинку и имаго. У нарывников рода Meloe и Mylabris данная дополнительная стадия называется « триунгулин » .
Другим исключением является неотения , представляющая собой способность некоторых организмов достигать половой зрелости и размножаться на стадии личинки . Примером могут служить самки рода Platerodrilus (семейство краснокрылов — Lycidae ). При этом в развитии самок исключаются сразу 2 фазы: куколка и имаго. Однако данное явление не исключает нормального развития самцов.

Партеногенез встречается у жуков довольно редко, но встречается во многих семействах, чаще всего у некоторых чернотелок , листоедов и особенно долгоносиков . При этом сохраняются все 4 фазы развития у самок, но среди имаго самцы могут полностью отсутствовать .
Живорождение (во всех случаях в виде яйцеживорождения) встречается крайне редко у нескольких филогенетически неродственных семейств. Первые виды жуков, у которых ещё в середине XIX века было обнаружено яйцеживорождение (у самок в яйцеводах в оболочках яиц находятся уже созревшие личинки), — стафилиниды Corotoca melantho ( Staphylinidae ; в подсемействе алеохарины ) и жуки- листоеды ( Chrysomelidae ; в трибе из номинативного подсемейства хризомелины ) . В 1913 году яйцеживорождение было обнаружено у жуков- микромальтусов ( Micromalthidae ) . В дальнейшем в этот список были включены жуки- жужелицы ( Carabidae ) и чернотелки ( Tenebrionidae ; в трибах Pedinini и из номинативного подсемейства Tenebrioninae ) . В 2016 году к этому списку добавились жуки-усачи ( Cerambycidae : Borneostyrax cristatus ) .
Своеобразна и сложна биология размножения жука микромальтуса ( Micromalthus debilis ), у которого известны три личиночных возраста. Личинки первого возраста очень подвижны, обладают длинными ногами и крупной головой, но не питаются. После линьки они превращаются в безногих личинок второго возраста, основной функцией которых является питание. Они, в свою очередь, превращаются в личинок третьего возраста, которые не питаются, но способны к размножению и рождают множество личинок первого возраста. Известны у данного вида и имаго , развивающиеся из личинок второго возраста, однако данный процесс изучен не полностью. У микромальтуса имеет место единственный среди насекомых случай сочетания гиперметаморфоза с размножением личинок — педогенезом .
Фаза яйца у жуков длится 2—3 недели. Развивающаяся личинка характеризуется 3—7 возрастами, разделяемыми линьками. Продолжительность развития личинок преимущественно зависит от климатических условий природной зоны и типа питания личинки. В умеренном климате у преобладающего большинства жуков за год развивается одно поколение, но у многих жуков, обитающих в почве и древесине ( хрущей , рогачей , щелкунов , златок , дровосеков , чернотелок и др.), развитие личинок продолжается от 2 до 6 лет, а при неблагоприятных условиях может затягиваться на более длительные сроки. При этом у божьих коровок, листоедов и некоторых других жуков на юге средней полосы и в субтропических районах может развиваться 2—3 поколения за год. У жуков, обитающих в тропических районах и вредителей запасов, обитающих в помещениях, за год обычно развивается несколько поколений, и развитие у них не прерывается на протяжении всего года .
Длительность стадии личинки и имаго у различных видов жуков неодинакова. Чаще (у большинства жужелиц , стафилинов , плавунцов , листоедов , некоторых долгоносиков, короедов и многих других представителей различных семейств) личинки заканчивают своё развитие за нескольких недель, а имаго могут жить значительно дольше, от нескольких месяцев до нескольких лет, повторно откладывая яйца и давая новые поколения жуков. В других группах жуков развитие личинок продолжается гораздо дольше, чем живут имаго (у ряда пластинчатоусых , щелкунов , рогачей , златок , некоторых усачей и др). Закончив откладывание яиц, данные жуки погибают. Иногда (у некоторых видов чернотелок ) личинки и имаго могут жить одинаково долго, порой по нескольку лет. Например, продолжительность жизни имаго чернотелки Polposipus herculeanus с Сейшельских островов может составлять до 8 лет . Имеет место также различная продолжительность жизни самок и самцов. У одних видов самцы появляются раньше самок и после спаривания быстро погибают, а самки продолжают жить, пока не отложат все яйца, созревающие у них порциями .
В умеренных широтах большинство жуков зимует в почве, лесной подстилке, некоторые — в древесине, под корой деревьев и т. п.; преимущественно в фазе имаго и личинки, гораздо реже на стадиях яйца или куколки. Фаза куколки у жуков является непродолжительной и обычно длится 2—3 недели .
Жуки характеризуются богатым набором проявлений инстинкта заботы о потомстве. Это обнаруживается, прежде всего, в заготовке запасов пищи для будущих личинок, либо в уходе за отложенными яйцами (или даже за личинками) .
Самое примитивное проявление заботы о потомстве заключается в обеспечении безопасности откладываемых яиц. Самым простым и распространённым является откладывание яиц в естественные убежища: трещины почвы или древесной коры, зарывание их в землю и т. п. Самки многих видов усачей выгрызают на коре кормовых деревьев специальные насечки и укладывают в них яйца . Ряд долгоносиков, например ( ), просверливают отверстия в плодах разных деревьев и откладывают в них яйца. Личинка при этом развивается внутри плода и выедает его мягкое ядро .

Более сложным способом является изготовление для яиц специальных чехольчиков. Листоеды из рода изготавливают защитную капсулу для яиц из собственных экскрементов, рода Galeruca склеивают специально выделяемым секретом яйца в защищённый «пакет», тропические щитоноски рода сооружают настоящие оотеки с несколькими ячейками. Калиновый листоед ( Galerucella viburni ) выгрызает на ветках своеобразное гнездо, откладывая туда несколько яиц и закрывая его крышкой из волокон древесины и экскрементов, скреплённых вместе выделяемым самкой секретом. Водные жуки, например водолюбы , обладают специальными паутинными железами. Секретом, выделяемым из этих желёз, они делают капсулы для своих яиц .
Ринхиты ( Rhynchites ) и близкие к ним роды откладывают яйца в ямки, просверлённые в различных сочных частях растений (молодых побегах, бутонах). Ещё одним способом защиты будущего потомства является изготовление из листьев т. н. «сигар» (или кульков-фунтиков) трубковёртами . Самки трубковёртов откладывают яйца в ткани листа, а затем делают на листьях хитроумные, математически точно рассчитанные надрезы, позволяющие свернуть из него плотную трубку. Затем самка обычно перегрызает главную жилку листа, после чего он увядает . Африканские долгоносики рода выгрызают укрытие для яиц в веточках, а затем обгрызают на их основании кору и луб — ветка погибает, а личинки получают корм. Похоже поступают некоторые американские усачи и африканские , способные обгрызть и ветки диаметром до 12 сантиметров .

Заготовка корма для личинок достигает наибольшей сложности у навозников и включает в себя как элементарное рытьё норок под кучами помёта, так и транспортировку его (после скатывания в шары) на большие расстояния от кучи в сложную систему подземных камер и ходов. Некоторые навозники-афодии , как, например, , самостоятельно не готовят пищу для своих личинок, а пользуются заготовленной другими видами: отыскивая туннели навозников Землерой обыкновенный , они откладывают свои яйца на подготовленные ими навозные лепёшки. Личинки афодия созревают раньше, выедая навоз и практически не оставляя пищи медленнее развивающимся личинкам вида-хозяина . Навозник из Австралии, находя высохшие экскременты животных, закапывает их так глубоко, что добирается до водоносных слоёв, где, отсыревая, они размягчаются и становятся пригодными для питания будущих личинок .
Жуки-кравчики (Lethrinae) заготавливают для своих личинок в подземных норках измельчённые травянистые растения — своеобразный силос . Мертвоеды , в частности жуки-могильщики , обеспечивают своих личинок пищей путём зарывания в землю трупов позвоночных животных. Вокруг мёртвого тела устраиваются подземные камеры, в которые откладываются яйца. После выхода личинок самка непрерывно кормит их, отрыгивая каждой личинке капли полупереваренной питательной смеси .
Некоторые жужелицы сооружают гнездовые камеры и охраняют яйцекладки до выхода личинок, а иногда вплоть до появления жуков из куколок. Сходное поведение наблюдается у самок навозников-копров (род Copris ).
Короеды не только делают под корой деревьев специальные маточные ходы, в которых откладывают яйца, но также заражают кормовые деревья особыми грибами, делающими древесину пригодной для питания личинок. Также самки короедов могут поддерживать в ходах определённую температуру и влажность, путём их вентиляции на протяжении всего времени развития личинок .
Некоторые жуки не сооружают убежища, а прикрывают своё потомство собственным телом. Так, самки водолюбов носят яйца на нижней стороне брюшка. А южноамериканские щитоноски из рода обладают расширенными боковыми краями надкрылий, под которыми укрываются до 20 молодых личинок .
Представители семейства пассалиды (Passalidae) отличаются высоко развитым инстинктом заботы о потомстве. Обитая в гнилой древесине, они часто встречаются в ней целыми скоплениями, образуя социальные группы . Самцы и самки живя парами, совместно выкармливают своё потомство пережёванной древесиной, смоченной слюной и прошедшей ферментацию с участием особых грибков. Эти жуки не только выкармливают личинок, но охраняют куколок и недавно вышедших из них имаго .
Межвидовое скрещивание у жуков не является редкостью. Для жуков-усачей индо-малайского рода Batocera зарегистрирован ряд гибридов , также известны межвидовые гибриды и в роде Monochamus , имеет место спаривание между различными видами рода Dorcadion и среди представителей подрода .
Во время развития жесткокрылых возможны генетические изменения, приводящие к появлению гинандроморфных жуков — полусамцов-полусамок, у которых одна сторона тела полностью мужская, а другая — женская (т. н. билатеральные гинандроморфы) .
Число хромосом у жесткокрылых сильно варьирует: диплоидные числа колеблются от 4 до 70. Существуют цитогенетические данные для более чем 4900 таксонов, и все жуки, размножающиеся половым путем, имеют гетерогаметных самцов, причем более 3000 видов обладают гетероморфными кариотипами половых хромосом XY. Из остальных видов более 700 являются XO и более 100 бесполыми. Подотряд Archostemata изучен по двум видам: Distocupes varians имеет 9 аутосом и детерминацию пола XO, в то время как Micromalthus debilis имеет диплоидное число хромосом 20 и циклический партеногенез, педогенез (размножение половозрелыми личинками) и гаплодиплоидию . Подотряд Myxophaga изучен по одному виду, . Диплоидное число у этого вида равно 20, а половые хромосомы имеют отдаленное спаривание XY. По подотряду Adephaga имеются данные по 1 221 виду из 7 семейств. Число хромосом у Adephaga самое низкое у , имеющего диплоидное число хромосом 8, и самое высокое у Dixus capito obscuroides , имеющего 70 хромосом. Крупнейший подотряд Polyphaga имеет наиболее распространенные половые хромосомы XY (более 1884 видов из 43 семейств). Половые хромосомы XO зарегистрированы в 18 семействах, а сложные системы половых хромосом обнаружены в 12 семействах. Число диплоидных хромосом колеблется от 4 у до 66 у .
Полиплоидия часто встречается у партеногенетических видов, а партеногенез выявлен в 16 семействах. Истинная гаплодиплоидия развилась по крайней мере один раз в подсемействе Scolytinae , и считается, что все (> 1200 таксонов) являются гаплодиплоидными, но это было исследовано только у нескольких видов. Другой вид короедов, не имеющий близкого родства с трибой Xylobrini, Hypothenemus hamperi , проявляет функциональную гаплодиплоидию в форме элиминации отцовского генома .


Среди жесткокрылых имеются представители большинства основных типов питания, которые известны в пределах класса насекомых . Жуки представлены как многочисленными хищниками (многоядными и специализированными), так и преобладающим большинством растительноядных форм ( фитофагов ). В числе последних имеются как виды, питающиеся листвой , так и потребители корней (ризофаги), цветов и пыльцы ( антофаги ), древесины и коры ( ксилофаги ), плодов либо семян ( ). Большинство фитофагов питаются живыми тканями растений, но некоторые также способны питаться сухой древесиной. Обширные биологические группы составляют жуки, питающиеся грибами ( ), а также виды, питающиеся гниющими и разлагающимися животными и растительными останками ( некрофаги и сапрофаги ) и детритофаги , которые питаются сухими или медленно разлагающимися веществами растительного и животного происхождения. Все перечисленные группы связаны множественными переходами .
Среди жуков (дровосеки, пластинчатоусые , щелкуны и другие семейства) существуют виды, имаго которых вовсе не питаются и живут за счёт запаса питательных веществ, накопленных на личиночной стадии развития. Но для большинства жуков, особенно долгоживущих, питание имаго («дополнительное питание») является необходимым для продления жизни и созревания яиц .
Характер питания личинок и имаго большинства групп жуков очень сходен, но у других питание этих фаз совершенно различно. Например, личинки бронзовок живут в почве, перегное , разлагающихся растительных останках, а жуки питаются на цветах или вытекающим соком деревьев. Жуки-усачи в личиночной фазе питаются, преимущественно в древесине или под корой деревьев, реже — в стеблях травянистых растений, а имаго многих видов встречаются на цветах и питаются пыльцой. В некоторых случаях личинки являются хищниками, тогда как имаго питаются растительными веществами .
Жуки, обитающие на земле, стволах деревьев, либо открыто на растениях, преимущественно передвигаются, ползая с разной скоростью, но некоторые из них (многие жужелицы , стафилиниды , златки , щелкуны , кожееды , грибоеды , пестряки и некоторые усачи ) могут быстро бегать. Жуки, обитающие на песчаных почвах (некоторые крупные жужелицы, скакуны и чернотелки ), а также лазающие по деревьям (многие усачи) обладают более-менее длинными, а в отдельных случаях изогнутыми ногами . Способность прыгать имеется у некоторых листоедов ( Alticinae , Sagra ), горбатки , , которые обладают сильно утолщёнными задними бёдрами. Прыжки ими используются не только в качестве способа передвижения, но и при спасении от хищников. Способность подскакивать имеется только у щелкунов , обладающих особой анатомической структурой на передне- и среднегруди. Подскок у них сопровождается характерным щелчком (порой достаточно громким) и используется для перевёртывания их тела на брюшную сторону, в случаях, когда жук оказывается лежащим на спинке, например, после падения на землю. Обладая короткими ногами, перевернуться иным способом щелкуны не могут .


Основной формой передвижения жуков в воздухе является машущий полёт . Перед взлётом жуки приподнимают надкрылья, а затем выдвигают и расправляют крылья . Именно поэтому жуки взлетают преимущественно медленно, а их полёт является сравнительно медленным. Быстрый взлёт и стремительный полёт отмечается только у мелких златок ( Agrilus ) и усачей ( Menesia ), а также скакунов .
Изменение направления движения в полёте сопровождается поворотом продольной оси тела. Скорость полёта может быть как низкой, так и высокой. Своеобразие полёта жуков заключается в низкой манёвренности и скорости, хотя последний показатель возрастает у крупных видов . Жуки и веерокрылые являются единственными насекомыми, использующими для полёта преимущественно заднюю пару крыльев. У летящего жука, в отличие от представителей других отрядов насекомых, основная часть тела находится впереди от работающих крыльев .
Некоторые жуки, как то бронзовки и Gymnopleurus , могут летать не приподнимая надкрылий, расправляя крылья через специальные выемки по бокам надкрылий . Отдельные представители жесткокрылых вообще не способны летать, так как у них крылья редуцированы или совсем отсутствуют .

Способность издавать звуки довольно широко распространена среди жуков и встречается у представителей примерно 20 семейств . Акустическая коммуникация является одним из важнейших средств общения у особей одного вида. При помощи звуков обеспечивается встреча самцов и самок, поддерживаются внутрипопуляционные и внутрисемейные отношения, достигается репродуктивная изоляция у близких видов. При контактах с другими видами звуковые сигналы преимущественно выполняют более узкие функции, как то отпугивания или угрозы. Большинство жуков, издающих звуки, делает это при помощи стридуляционного аппарата. Он наиболее часто встречаются у усачей , у которых звук возникает при движениях переднегруди относительно среднегруди, благодаря скольжению заднего края по стридуляционной площадке. Если держать усача за брюшко, то можно легко заметить, как он издаёт скрипучие звуки, двигая вверх-вниз передней частью тела. У ряда капюшонников на боках переднегруди находятся ряды мелких бугорков, а звуки издаются при проведении по этим бугоркам вершинами передних бёдер .
Представители некоторых семейств издают звуки, ударяя головой или концом своего брюшка по субстрату (последний способ характерен для чернотелок ). «Часы смерти» — именно так у многих народов называют «тикающие» звуки, издаваемые некоторыми мебельными и домовыми точильщиками ( Anobiidae ), которые селятся в изделиях из древесины . Эти виды точильщиков издают данные звуки, похожие на звук тиканья часов , ритмично ударяя головой о стенки своих ходов в древесине. Эти звуки являются способом привлечения самок .


Биолюминесцентное свечение характерно для светляков , некоторых тропических видов щелкунов , а также самок и личинок жуков семейства Фенгодиды . При этом количество, форма и расположение световых органов у различных видов варьируют. Так, у некоторых тропических видов мелкие световые органы находятся на дорсальной стороне каждого стернита брюшка. У щелкунов рода огненосных щелкунов ( Pyrophorus ) два небольших световых органа находятся на верхней стороне переднеспинки, также крупный светящийся орган находится на первом стерните брюшка. У большинства видов имеется только один крупный светящийся орган на вентральной стороне двух последних брюшных стернитов. В основе свечения лежит реакция окисления люциферина при участии специфического фермента люциферазы , а также АТФ и ионов магния .
У светляков встречаются сигналы, связанные с половым поведением (поисковые и призывные сигналы у самцов и самок), защитные и территориальные сигналы и даже световая мимикрия . Некоторые виды, например, обыкновенный светляк ( Lampyris noctiluca ), обладают только призывным сигналом. У преобладающего большинства видов рода и Photuris отсутствуют различия между поисковыми и призывными сигналами у самцов. В то же время только у самок рода Photuris описано явление световой мимикрии, когда самки испускают световые сигналы готовности к спариванию, характерные для видов рода Photinus , которые обитают на одной с ними территории. Самцы Photinus , привлечённые данными сигналами, «попадают в ловушку» и служат пищей для хищных самок рода Photuris .
У светляков выделяют несколько типов световой коммуникации. Особи одного пола (преимущественно не летающие самки) испускают видоспецифичные сигналы, привлекающие самцов (род Lampyris , Pyrophorus и др.), наличие собственных световых сигналов у летающих особей при этом необязательно. В другом случае, летающие особи одного пола (преимущественно самцы) производят видоспецифичные световые сигналы, в ответ на которые особи другого пола испускают световые «ответы» (подсемейство ). Существуют также виды с промежуточными вариантами описанного выше поведения .
Самки южноамериканских светляков из рода способны излучать свет зелёного и красного цвета, причём делая это поочерёдно либо одновременно .
Феромоны используются жуками для самых разнообразных целей. Самыми важными являются половые феромоны, облегчающие встречу самцов и самок. Обычно половые феромоны выделяются самками, но также отмечены случаи выделения их и самцами. Также жуки выделяют агрегационные феромоны, помогающие особям одного вида образовать скопление, необходимое для питания или зимовки .

Время лёта и его продолжительность весьма разнообразны у различных групп жуков и ограничены тёплым временем года в умеренных широтах. Наиболее ранними формами на юге средней полосы России являются преимущественно весенние виды. Наибольшее число видов появляется в средней полосе в июне — июле, в южных областях — в мае — июне. Также в средней полосе имеется несколько видов, лёт которых приходится на конец лета — начало осени. Таким образом, в средней полосе на протяжении вегетационного периода растений происходит постепенная смена фаунистического состава жесткокрылых, при этом некоторые виды живут в течение практически всего тёплого сезона — с весны до осени .
Жуки, питающиеся на растительных культурах и цветах, активны преимущественно днём, и наиболее подвижны в жаркие часы дня (например бронзовки , усачи Lepturinae и др.), а на ночь прячутся в укрытия. Другие же жуки, наоборот, активны вечером и в сумерках, а днём преимущественно прячутся в укромных местах (например усачи Prioninae , жужелицы и др.) . У одних видов более активными и склонными к перелётам являются самцы, у других — самки .
Божьи коровки могут образовывать массовые скопления в местах зимовок. Причины зимних скоплений и предшествующих им миграций, возможно, связаны с историческим становлением данного семейства, ведь скопления характерны именно для стран с холодной зимой, которые заселялись теплолюбивыми видами, изменившими свой тип питания и образ жизни .
Некоторые жуки также совершают групповые перелёты. Например, плавунцы , водолюбы иногда совершают перелёты довольно большими стаями при переселении в новые водоёмы .

Самцы многих рогачей и некоторых дупляков часто бывают агрессивными по отношению к другим самцам своего вида. Они вступают в драки между собой из-за мест питания (например, мест с вытекающим древесным соком) или самок. Такие сражения обычно происходят на стволах деревьев. Например, самцы многих рогачей, заметив противника, принимают угрожающую позу — приподнимают переднюю часть тела вверх и широко расправляют в стороны усики. Если угрозы не действуют, самец атакует противника. При этом самцы обычно высоко поднимаются на передних и средних ногах («вставая на дыбы»), широко раскрывают челюсти. В ходе таких драк, каждый из соперников пытается подцепить противника за надкрылья своими мандибулами, приподнять его в воздух, а затем скинуть вниз .
Во время лёта у крупных усачей ( чёрные усачи , Prionus и др.) иногда между самцами могут возникать драки, напоминающие брачные турниры, в ходе которых они откусывают друг другу усики и части конечностей . Также у скарабеев между жуками могут возникать драки за обладание готовым скатанным шариком навоза .
Жуки широко распространены по всему земному шару, в шести зоогеографических областях , кроме Антарктиды , ледниковой зоны Арктики и наиболее высоких горных вершин. Наиболее богато отряд представлен в тропических регионах .

Жесткокрылые населяют сушу и пресные водоёмы по всему земному шару — начиная от самых засушливых пустынь и заканчивая тропическими лесами , от экваториальных областей до тундры. Жуки отсутствуют только в областях, которые покрыты вечным ледниковым покровом ( Антарктида , некоторые районы Арктики и наиболее высокие горные вершины). На Кавказе жесткокрылые поднимаются на высоты более 3000 м, в горах Средней Азии до 4500 м, а в Гималаях и горных регионах тропиков до 5000—5500 м . Например, жужелица и ещё около 25 видов рода бегунчики в Гималаях населяют только пояс высот 4300—5000 метров над уровнем моря в зоне вечного снега .
В пределах области распространения жуки могут встречаться практически повсюду. Многие из них обитают в верхних слоях почвы, их личинки и куколки проводят в них свою жизнь, а имаго обычно на короткое время выходят на поверхность (исключением являются некоторые мелкие жужелицы , стафилины , долгоносики и др.). Не менее многочисленными являются обитатели растений. Одни жуки обитают под корой или в древесине , другие — на листьях (изредка даже внутри их тканей), в цветах и плодах , третьи — в корнях . На растениях также охотятся многие хищные жесткокрылые ( божьи коровки , ряд жужелиц ). Представители некоторых семейств селятся на древесных и наземных грибах , питаясь ими (мицетофаги), либо охотясь на других животных, обитающих здесь же. Множество жуков являются обитателями навоза или селятся на падали — одни из них питаются разлагающимися веществами, другие же хищничают , в частности, охотятся на личинок мух .
В пресных водоёмах также обитают многие жесткокрылые. Многие из них являются представителями специфических водных семейств ( плавунцы , вертячки , водолюбы и др.), другие являются специализированными представителями семейств, населяющих главным образом сушу. Жуки встречаются в пресных водоёмах любых типов (включая подземные), однако преобладающее большинство из них предпочитают небольшие, неглубокие, слабо проточные водоёмы с обильной растительностью. В некоторых семействах жуков практически все виды в любой стадии развития обитают в воде, а в других семействах в воде могут обитать только некоторые виды, и часто лишь в одной фазе своего жизненного цикла.

Некоторые плавунцы и вертячки встречаются также в солоноватой воде. Однако в морской воде нормальной солёности жесткокрылые не живут. Среди жуков также имеются специфические обитатели берегов пресных или солёных водоёмов . Среди последних существуют виды, встречающиеся исключительно на сильно засолённых почвах ( ) или предпочитающие её ( галофилы ) . А некоторые, даже могут погружаться в солёную воду. Например, вид Cicindis horni — плавающая хищная жужелица, которая обитает на побережьях солёных озёр в Аргентине . При этом она не только может плавать на поверхности воды, но и погружается в глубь в поисках добычи (мелких планктонных жаброногих рачков) .
Солончаковые биотопы и берега осолонённых водоёмов населены характерной и своеобразной фауной жуков, связанных в своём развитии с осолонёнными почвами или с солончаковой растительностью (мелкие жужелицы , включая род скариты , скакуны , эндемичные златки, усачи, долгоносики ) .
Жуки, населяющие пустыни и полупустыни , характеризуются наличием суточного ритма с преимущественно ночной активностью, выносливостью к высоким температурам и способностью обеспечивать водный баланс организма часто за счёт метаболической воды. Для многих видов характерна кратковременность стадии имаго и её афагия . Характерными пустынными обитателями являются чернотелки и некоторые жужелицы .
Особый интерес представляет жук Zaitzevia thermae , являющийся эндемиком , обитающим в термальном источнике штата Монтана ( США ). Жуки длиной тела всего 2 мм и их личинки живут в необычно тёплой воде с температурой 29 °C .
Большую и разнообразную группу составляют жуки, постоянно живущие в муравейниках , называемые мирмекофилами . Также обширную биологическую группу составляют жесткокрылые- синантропы (спутники человека), способные поселяться в жилище человека, хозяйственных постройках, продовольственных складах, зернохранилищах и т. д. ( кожееды , некоторые виды пестряков , притворяшек , плоскотелок , чернотелок , зерновок и др.) .
Взаимоотношения жуков с другими организмами очень разнообразны и включают мимикрию , комменсализм , паразитизм и мутуализм .

Примером комменсализма , с выгодой извлекаемой жуками, могут служить некоторые виды кожеедов , связанные с пауками , в гнёздах которых они питаются сухими остатками пойманных ими насекомых .
Частным случаем комменсализма у жуков является форезия — перенос на насекомом-хозяине других организмов. У жуков примером этого явления могут служить «путешествующие» на мертвоедах , жуках-могильщиках , навозниках , карапузиках мелкие клещики из рода и семейств Acaridae , , Parasitidae , Uropodidae и других . А у южноамериканского усача Acrocinus longimanus имеются симбиотические взаимоотношения с ложноскорпионами рода , живущими под надкрыльями жуков, и питающимися клещами - паразитами .
Среди жуков существует целый ряд мирмекофилов , поддерживающих тесные связи с муравьями . Они питаются различными отбросами муравьёв или гниющим строительным материалом гнезда, но многие являются «нахлебниками» . У многих из них выработались специальные структуры для сожительства с муравьями. У некоторых на теле имеются специальные железы, выделяющие вещества, которые слизывают муравьи. Например, у пауссид они находятся на усиках, а у жуков-ощупников по бокам тела . К мирмекофилам относятся также и некоторые карапузики , стафилины и прочие. Так, 19 родов стафилинов «сожительствует» с бродячими муравьями, 17 родов с огненными муравьями и 15 родов с представителями подсемейства формицин . Стафилины выпрашивают у муравьёв-хозяев корм, прикасаясь к нижней губе, и защищаются в случае нападения, выделяя репелленты . Стафилины, обитающие с бродячими муравьями , имитируют муравьёв формой и окраской своего тела (пример бейтсовской мимикрии).
Также среди жуков существует ряд термитофилов , поддерживающих связи с термитами или обитающих в термитниках . Они имеют как морфологические, так и поведенческие реакции, позволяющие им вступать в тесные симбиотические связи с термитами. Полностью термитофильными являются несколько узкоспециализированных триб и родов алеохарин (Corotocini, Feldini , Pseudoperinthini , Termitopaedini , Termitocharina, Termitocupidina, Termitogastrina, Trichopseniini) . Термитофилия отмечена у южноамериканского рода из трибы , у личинок жужелиц и и некоторых других жуков. А Coatonachthodes ovambolandicus — вид мелких термитофильных стафилинов обладает уникальным по форме брюшком, которое сверху похоже по форме на рабочего термита с отростками, напоминающими ноги и имитирующими усики .
Примером комменсализма муравьёв и жуков являются некоторые виды бронзовок , например, бронзовка металлическая , личиночные стадии которой проходят в муравейниках рыжих и малых лесных муравьёв, красногрудых муравьёв-древоточцев и некоторых других .
Комменсалами ряда пилильщиков и орехотворок являются некоторые долгоносики рода , которые откладывают свои яйца в их галлы на различных древесных растениях, например: в галлы орехотворок на дубе , в листовые галлы рода Nematus .
Ряд южноамериканских видов пластинчатоусых жуков из рода являются эктокомменсалами млекопитающих и живут в шерсти некоторых обезьян . Виды родов и обнаружены в шерсти ленивцев или у заднего прохода центральноамериканского тапира . В Австралии в аналогичных условиях на кенгуру живут представители рода .
Многие кожееды связаны с насекомыми и частично с пауками . Значительная группа видов относится к числу симбионтов различных перепончатокрылых . Некоторые из них развиваются в гнёздах шмелей , другие обитают в норах ос и пчёл .
Большинство грибоядных и потребляющих мёртвую древесину жуков имеют кишечных симбионтов , относящихся к дрожжевым или дрожжеподобным грибам из Ascomycota . Ключевым аспектом взаимоотношений жуков и грибов является наличие у первых таких особенностей, которые способствуют распространению спор, конидий и частиц мицелия , которые переносятся жуками на поверхности своего тела . Некоторые жуки из подсемейства Scolytinae и Platypodinae разводят в проделанных в древесине ходах «грибные сады»: рассеивают в них споры грибка, который становится пищей для их личинок .
Для многих представителей долгоносиков из рода Gymnopholus (особенно для подрода Symbiopholus ) характерен симбиотический рост некоторых растений , водорослей , лишайников , мхов-печёночников и грибов на поверхности кутикулы в верхней части модифицированных надкрылий .
Паразитизм среди жуков встречается довольно редко. Среди жуков существуют паразитоиды , являющиеся эктопаразитами. Личинки нарывников , некоторых пестряков паразитируют в гнёздах одиночных пчёл и ос, в кубышках саранчовых . Также личинки некоторых жужелиц рода Lebia паразитируют на куколках листоедов или других групп жуков . Личинки веероусых жуков являются эктопаразитами личинок жуков-точильщиков и некоторых перепончатокрылых . Также имеют место отдельные случаи перехода к эндопаразитизму личинок некоторых родов веероусых жуков, которые живут в телах тараканов .
Имаго некоторых жуков являются эктопаразитами или полупаразитами млекопитающих , например, жуки Platypsyllus castoris паразитируют на речных бобрах , вероятно питаясь их отмершей кожей, род на полёвках , на выхухоли . Также некоторые южноамериканские стафилины являются эктопаразитами грызунов , а их тропическая группа в стадии имаго паразитирует на позвоночных .
Жук из семейства пластинчатоусых обитает в гнёздах муравьёв рода Solenopsis и изредка поедает куколок муравьёв . Также в муравейниках паразитируют жуки из рода ломехуза , имаго и личинки которых питаются яйцами и личинками муравьёв .
Образ жизни и жизненные формы жуков являются настолько разнообразными, что у представителей отряда можно встретить почти все известные у насекомых защитные механизмы .
Для многих видов жуков, как средство защиты, характерен танатоз — временная неподвижность, при которой жуки притворяются мёртвыми. При вероятной опасности жуки обычно замирают и падают с растений на подстилку. Данное поведение характерно для многих групп жуков, в том числе долгоносиков , листоедов , пилюльщиков и др .

Ряд видов спасаются от хищников быстрыми передвижениями: бегом ( жужелицы ), полётом ( бронзовки ), плаваньем ( вертячки ). Скакуны используют моментальный взлёт. Рогачи и скариты при опасности используют угрожающие движения и позы: например, самцы жуков-оленей при опасности приподнимают переднюю часть тела вверх, раскрывают мандибулы и широко расправляют в стороны усики .
Некоторые жуки, например, многие виды усачей , могут издавать резкие скрипящие звуки, воспроизводимые путём трения рёбрышка на заднем крае переднегруди о шершавую поверхность среднегруди. Данные скрипящие звуки используются жуками в случае нападения хищников и носят отпугивающий характер .
Многие виды обладают покровительственной окраской тела. Некоторые виды чернотелок (Tenebrionidae), (Georyssidae), водолюбов (Hydrophilidae) покрывают себя грязью, глиной, илом или песком. Для жуков также характерен — одна из форм мимикрии, достигаемая путём сходства с деталями окружающей обстановки или заведомо несъедобными предметами. Например, сходство формы или окраски тела щитоносок (Cassidinae) с галлами на листьях, щелкунов рода и долгоносиков рода Alcides с птичьим помётом, пилюльщиков ( Byrrhidae ) с высохшим козьим помётом или камешками и т. п. Примером также может служить окраска некоторых долгоносиков из рода , имитирующих своим видом высохшее насекомое с отверстием, оставленным после выхода из него эндопаразита .

Апосематизм — предостерегающая окраска и форма тела. Классическим примером является яркая окраска, преимущественно представленная как сочетание красного или жёлтого цветов с чёрным, у жуков, обладающих ядовитой гемолимфой : у божьих коровок ( Coccinellidae ), нарывников ( Meloidae ), краснокрылов ( Lycidae ) и многих других. Примером такого явления также может служить выпячивание, в случае опасности, красных пузырей по бокам тела у малашек (род Malachius ) .
Синапосематизм — ложная или мюллеровская мимикрия — согласованная, сходная окраска и форма тела у нескольких различных видов, обладающих развитыми прочими средствами защиты от хищников .
Псевдоапосематизм — истинная, или бейтсова мимикрия . При данной форме мимикрии виды, не обладающие защитными механизмами, обладают такой же окраской и формой тела, как один или несколько защищённых видов . Ряд видов жуков-усачей ( Cerambycidae ) часто подражает жалящим перепончатокрылым . Интересно, что кроме сходства в окраске и форме тела, иногда у них имеется и сходство в поведении: усачи двигаются быстро и порывисто, «ощупывая» субстрат вытянутыми вперёд усиками, подражая своим поведением осам .
Многие златки , бронзовки и другие обладают очень твёрдыми и прочными покровами тела, защищающими их в той или иной степени от хищников. Ряд жуков обладает устрашающими и порой весьма небезопасными челюстями: рогачи , жужелицы , некоторые усачи . Некоторые группы характеризуются наличием острых и длинных шипов на переднеспинке и надкрыльях — усачи, листоеды ( Chrysomelidae : Hispinae ), грибовики .

Среди жуков довольно часто встречаются виды, обладающие ядовитой гемолимфой . Самыми распространёнными ядовитыми компонентами являются кантаридин и педерин . Наиболее ядовитые (при поедании их хищником) жуки обычно относятся к представителям нарывников , божьих коровок , краснокрылов , мягкотелок , малашек , листоедов , стафилинид .

Некоторые обладают железами с ядовитыми и пахучими секретами. Самым ярким примером подобного способа защиты являются жуки-бомбардиры ( Brachininae ) . Они обладают железами, выделяющими смесь химических веществ, которые, вступая во взаимодействие друг с другом в специальной камере брюшка, вызывают экзотермическую реакцию и разогрев смеси до 100 °C. Образующаяся смесь веществ выбрасывается наружу через отверстия на кончике брюшка . Представители подсемейства пауссин ( Paussinae ) обладают менее подвижным брюшком и при необходимости атаковать врага, находящегося спереди, они выпускают горячую жидкость на специальные выступы надкрылий , направляющие её вперёд . Лучше всего эти выступы видны у жуков трибы . выбрасывает непульсирующую струю со скоростью 2,4 м/с . Более примитивный механизм защиты описан у представителей трибы — они не формируют струи, как остальные бомбардиры, а выделяют пузырящуюся и брызгающуюся в разные стороны жидкость .
Жужелицы рода Carabus способны также выбрызгивать очень едкую жидкость, которая может вызвать раздражение кожных покровов человека. Медляки рода Blaps при опасности занимают определённую позу и выделяют из специальных желёз жидкость с неприятным запахом. Ядовитый секрет с неприятным запахом выделяется также грудными железами плавунцов ( Dytiscidae : Dytiscus ) .
Ряд видов практикует сожительство с защищёнными животными . Примером может служить мирмекофилия — выгодное для жуков сожительство с муравьями в их гнёздах, где они находят не только защиту, но и пищу (некоторые виды ощупников ( Pselaphidae : ), стафилинид (Staphylinidae), карапузиков (Histeridae)). Некоторые виды жуков предпочитают вести скрытный образ жизни, обитая в труднодоступных местах, надёжно защищённых от врагов ( короеды (Scolytidae), а также обитающие в почве виды). Другие ведут ночной образ жизни, который действенно предохраняет их от возможных нападений птиц и других дневных хищников, включая муравьёв. Примеры жуков, активных ночью, можно найти среди большинства семейств .
Жуками могут питаться различные членистоногие — пауки , скорпионы , многоножки , и другие насекомые . Также жуки служат пищей многим видам земноводных , пресмыкающихся . Рацион многих птиц , например, серой вороны , сороки , а также чеглока , лугового луня , сизоворонки , сов и многих других включает жуков . Ряд млекопитающих — насекомоядные , летучие мыши , мелкие хищники и некоторые другие — могут поедать жесткокрылых .
На жуках паразитируют наездники -терзилохины ( , , , , и ). Наиболее часто в качестве хозяев выступают жуки-долгоносики ( Curculionidae ). Наездники эфиальты (подсемейство Ephialtinae ) из семейства ихневмонид ( Ichneumonidae ) паразитируют на личинках усачей и златок. Личинки сколий являются паразитоидами личинок различных жуков: гигантская сколия на личинках жука-носорога и жука-оленя , сколия степная на личинках других пластинчатоусых жуков . Найдя личинку, самка сколии парализует её уколом жала в брюшной нервный ганглий, после чего откладывает на неё одно яйцо. Вышедшая из него личинка сколии питается живой, но парализованной личинкой, начиная поедать её с наименее важных жизненных органов.

В силу огромного многообразия, большой численности и широкого распространения, роль жесткокрылых в природе исключительно велика. Имаго и личинки видов, населяющих почву и лесную подстилку, принимают активное участие в процессах почвообразования, гумификации мёртвой древесины .
В естественных и, в незначительной мере, изменённых экосистемах , жуки- ксилофаги ( усачи , златки и др.) выполняют санитарную роль, элиминируя мёртвую древесину и утилизируя ослабленные, сухостой , поражённый корневым грибком ( Heterobasidion annosum ), трутовиками , корневой гнилью деревья, в определённой мере регулируя распространение этих грибов . Особую роль они играют на буреломах и вырубках, где ускоряют разложение мёртвого дерева . Элиминация таких деревьев освобождает место для молодой поросли и способствует восстановлению экосистем.
Многие жуки также выступают опылителями цветковых растений , ведь значительная часть этих насекомых характеризуется развитием антофилии . Имаго подобных видов часто встречаются на цветах, где в отличие от «классических» опылителей ( пчёлы , шмели , чешуекрылые , двукрылые и т. п.) они проводят значительно больше времени, и, соответственно, эффективность опыления оказывается выше. Также эти виды являются регуляторами численности тех цветочных растений, которые они опыляют . Это связано с тем, что взрослым насекомым для жизнедеятельности требуется питание пыльцой , при этом жуки часто поедают гинецей и андроцей , уменьшая продукцию семян растений .
Крупные представители подсемейства Scarabaeinae могут быть промежуточными хозяевами ряда гельминтов , в том числе патогенных для домашних животных и порой для человека . Также скарабеины являются главнейшими природными санитарами, очищающими поверхность почв от разнообразных экскрементов . Утилизация жуками масс навоза способствует их перемещению в нижние слои почв, которые разрыхляются и удобряются . Виды, питающиеся различными разлагающимися веществами ( стафилины , мертвоеды , карапузики и др.) выполняют санитарную роль и способствуют утилизации животных и растительных остатков .

Древнейшие палеонтологические находки жуков, по одним данным, относятся к концу каменноугольного периода (320—300 млн лет назад) , по другим — к пермскому периоду (299 млн лет назад) .
На сегодняшний день признаётся 166 семейств жесткокрылых, живущих в настоящее время. Ещё 67 семейств известны из отложений палеозоя и мезозоя , но только 19 из них вымерли до кайнозоя (в том числе все палеозойские семейства) .
Практически все надсемейства жуков сформировались до или в течение кризиса мелового периода . Устойчивость многих групп жуков может объясняться их происхождением как подкорных и скважных обитателей ксероморфной флоры голосеменных на рубеже карбона и перми , а также сохранением связи с исходными местами обитания на протяжении длительного периода времени .
Считается, что исходной группой, из которой в дальнейшем сформировался отряд Жесткокрылых, было семейство чекардоколеиды ( Tshekardocoleidae ), которых относят к подотряду . Представители подотряда возникли в каменноугольном периоде . К началу пермского периода от них отделилась эволюционная ветвь, которых выделяют в надсемейство . Пермокупедоидеи просуществовали до начала мелового периода (120—145 млн лет назад) и вымерли. Однако ещё в нижнем триасовом периоде — 240—250 млн лет назад, от этой группы отделилась эволюционно очень успешная группа архостемат (Archostemata), которые существуют и сейчас. Архостематы образуют отдельный подотряд в отряде жесткокрылых и являются самыми древними из ныне живущих представителями этого отряда. Для архостемат характерно развитие личинок в древесине, которая служит для них источником пищи, а поскольку условия среды в древесине в ходе эволюции изменялись мало, жуки этого подотряда не испытывали воздействия резких изменений климата и не вымерли. Ранним триасом датируются также и первые находки повреждений древесины насекомыми — в окаменевших стволах голосеменных видны ходы личинок . Пермокупедоидеи включали семейство адемосинид ( ), которые вымерли вместе с ними в меловом периоде, однако в среднем триасе это семейство дало начало подотряду всеядных жуков (Polyphaga) .

К пермским жукам также относится ещё два надсемейства: и . Если последнее надсемейство просуществовало до начала юрского периода и вымерло, то первое в верхнем триасе развилось в инфраотряд шизофориформ ( ), от которого произошли два важных подотряда жуков — миксофага (Myxophaga) и плотоядные жуки (Adephaga), успешно существующие и сегодня .

Палеонтология на данный момент не может дать достоверного ответа на вопрос о происхождении жуков, поэтому он остаётся доподлинно неизвестным. Переходных форм между ними и какими-то другими группами насекомых, имеющих полное превращение, пока не обнаружено, хотя существует определённое сходство между древнейшими жуками и насекомыми с неполным превращением . В геологических отложениях каменноугольного и пермского периодов палеозоя встречаются две группы насекомых, очень похожие на древних жуков, — это и тараканы .
Проведённые в 2007 году исследования ДНК ныне живущих жуков и схемы вероятной эволюции жесткокрылых, свидетельствовали, что они, вероятно, возникли в нижней перми около 299 млн лет назад . Однако в 2009 году ископаемый жук возрастом 318 млн лет был описан из в США . Находки жуков, которые датируются как наиболее древние, обнаружены в отложениях пермского периода в Азии и Европе, например в красных сланцах Нидермошель , вблизи Майнца, Германия . Однако многие учёные сомневаются в правильности определения этих находок в качестве жуков. Научно задокументированы древнейшие находки жуков сакмарского яруса пермского периода с Босковицкого Грабена, Восточная Чехия . Эти находки являются очень разнообразными и относятся к 8 родам, принадлежащим к семейству чекардоколеиды ( Tshekardocoleidae ). Их надкрылья характеризуются решётчатой структурой с жилкованием, похожим на таковое у древних , с которыми, возможно, эти жуки имеют общее происхождение. Их крылья также были неспособны складываться на спине и лежали вдоль брюшка, а надкрылья были значительно длиннее брюшка. Другие признаки, как например, подвижная переднегрудь, слитные и скрытые стерниты заднегруди, погружённые в грудь тазики задних ног, склеротизированные базальные членики брюшка и т. п. не отличались от современных жуков.
| Coleoptera |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Находки жуков с кунгурского яруса пермского периода на Урале , возле деревни Чекарды в Пермском крае , Россия , принадлежащих к чекардоколеидам, свидетельствуют о том, что структура надкрылий значительно изменяется — жилки выпрямлены, с меньшим количеством ячеек. В середине пермского периода надкрылья жуков структурно не отличаются от современных — жилки расширяются, полностью заполняя пространство ячеек, а последние уменьшаются до полых колонок-колумел. Процесс становления надкрылий происходит двумя путями. Первый — у купедоид жилки преднадкрылий образуют механически совершенную решётчатую структуру, и только после этого начинают разрастаться, вытесняя ячейки, которые приобретают вид штриховых бороздок. Второй — у схизофороидов разрастание жилок происходит без промежуточных стадий специализированного жилкования. Одновременно во всех группах жуков возникают гладкие надкрылья: считается, что это является приспособлением к жизни в водной среде, поскольку гладкие надкрылья вызывают меньшую турбулентность водного потока и обеспечивают эффективное движение в толще воды.
В начале пермского периода жуки являются чрезвычайно редкими животными, даже в массовых захоронениях насекомых они составляют не более 0,1—0,6 % от всей энтомофауны. А в многочисленных отложениях артинского яруса пермского периода Северной Америки, где найдено много остатков насекомых, жуки вообще отсутствуют, и это в то время, когда Европа и Северная Америка составляли единый континент .
Эволюция подсемейства пластинчатоусых жуков Scarabaeinae была неразрывно связана с эволюцией крупных позвоночных животных: прежде всего млекопитающих , а на ранних этапах, возможно, и растительноядных динозавров — надо полагать, что подсемейство сформировалось уже в меловом периоде , а частично уже в юре . Представители семейства усачи в ископаемом состоянии известны уже из отложений юрского периода .
Отряд Жесткокрылые включает четыре подотряда: Архостемата ( Archostemata ), Миксофага ( Myxophaga ), Плотоядные жуки ( Adephaga ) (или Хищные жуки), и Разноядные жуки ( Polyphaga ). Подотряды жуков-архостемат и миксофагов являются наименьшими, а самым крупным — Всеядные жуки, которых насчитывается около 300 000 видов. Подотряд Хищные жуки насчитывает около 50 000 видов. Традиционные классификации (или системы) отряда жуков включают около 150 семейств . Согласно последним данным на 2011 год в отряде Жесткокрылые признано 24 надсемейства, 211 семейств, 541 подсемейство, 1663 трибы и 740 подтриб . По числу видов преобладают стафилиниды (Staphylinidae, >56000 видов) и долгоносики (Curculionidae, >51 000 видов), а по количеству родов лидируют усачи (Cerambycidae, >5232 родов) и долгоносики (Curculionidae, >4600 родов) .
Личинки жуков потребляются в пищу человеком значительно чаще, чем взрослые особи, их обычно едят сырыми или жареными . В Лаосе , Мьянме , Индии , Таиланде и на Филиппинах крупные личинки и куколки ряда видов пластинчатоусых жуков являются деликатесом в местной кухне . Личинок усачей употребляют в пищу в Юго-Восточной Азии , Шри-Ланке и Папуа — Новой Гвинее . У местных племён Фиджи деликатесом считаются личинки некоторых усачей , а у племён парагвайских индейцев личинки пальмовых долгоносиков (род Rhynchophorus ). Мучных хрущаков широко употребляют в пищу в Южной Америке , в Индии, на Ближнем и Среднем Востоке . Жаренных личинок майских жуков едят во Франции , где из них также варят супы .
Личинки и куколки листоедов африканского рода Diamphidia выделяют яд диамфотоксин , используемый бушменами для смазывания наконечников отравленных охотничьих стрел .

Многие племена Африки , Южной Америки и Океании , используют засушенных жуков (например златок ) или их блестящие ярко окрашенные надкрылья для изготовления различных украшений, преимущественно бус . Также в качестве украшений могут носить живых жуков. Для изготовления «живых брошей» используются некоторые виды жуков из семейства зоферид ( Zopheridae ), надсемейства Тенебрионоидных , которые водятся в Мексике и на юге США . Их украшают искусственными или настоящими драгоценными камнями , а к надкрыльям прикрепляют тонкие цепочки . В Мексике обычай использовать жуков одновременно и как украшения, и как домашних питомцев существует на протяжении многих веков . В конце XIX века огненосные щелкуны вошли в моду в Париже в виде вечерних украшений, названных «живые алмазы» . Данная мода впервые возникла ещё среди мексиканских женщин, которые украшали себя живыми жуками, которых клали в специальные тюлевые мешочки, прикалываемые к платьям . Первые европейские переселенцы в Южной Америке использовали огненосных щелкунов для освещения своих жилищ, а местные племена издавна привязывали их к пальцам ног, путешествуя ночью через джунгли , чтобы освещать дорогу .

В судебно-медицинской экспертизе имеется метод определения давности убийства по составу жуков- некрофагов и их личинок, обитающих на трупе .
Амброзиевый полосатый листоед ( Zygogramma suturalis ) был интродуцирован в СССР , ряд стран Европы, Китай и Австралию из Канады и США для борьбы со злостными сорняками — амброзиями ( Ambrosia artemisiifolia , ) . Для борьбы с рядом вредителей лесных культур эффективными являются божьи коровки , например, удивительная коровка используется в борьбе с тополёвым листоедом ( Chrysomela populi ). Личинки других божьих коровок истребляют тлей , кокцид , листоблошек . Красотел пахучий используется в качестве биологического метода борьбы с гусеницами непарного шелкопряда , и, начиная с 1906 года, с этой целью данный вид неоднократно интродуцировали из Европы в США .



Отрицательное значение жуков также заметно. Личинки и жуки ряда видов пластинчатоусых являются вредителями лесных, плодовых и сельскохозяйственных культур. Например, некоторые виды майских жуков являются одними из основных вредителей плодово-ягодных и лесных насаждений в Центральной Европе, лесостепной зоне, на Северо-Востоке Украины. В годы массового размножения эти жуки могут полностью объедать листья деревьев . Некоторые тропические виды из рода Oryctes и других вредят кокосовым пальмам , таро , сахарному тростнику и т. п.
Большую группу вредителей образуют листоеды, наносящие вред посевам и молодым посадкам различных лесных и сельскохозяйственных растений. Особо нужно отметить колорадских жуков и их личинок, питающихся исключительно мякотью листьев растений семейства паслёновых , преимущественно картофеля .
Большое значение имеют жуки как вредители леса. Среди них имеется довольно много листогрызущих видов, которые могут систематически или периодически размножаться в массе. Более разнообразны и многочисленны вредители древесины ( жуки-усачи , короеды , златки , жуки-точильщики ), среди которых есть опасные вредители, повреждающие здоровые деревьях и вызывающие их ослабление и усыхание, а также технические вредители, снижающие качество заготавливаемой древесины .
Множество видов личинок жуков-щелкунов являются вредителями культурных растений . Некоторые виды прогрызают в клубнях картофеля отверстия и повреждают молодые стебли , другие вредят винограду , объедая почки и молодые побеги .
Многие виды кожеедов являются опасными вредителями запасов животного и растительного происхождения, шелководства и музейных коллекций. Часто повреждают кожевенное сырьё, меха, перо, шерстяные изделия, мясные продукты, сушёную и копчёную рыбу, музейные экспонаты, чучела животных, зоологические и энтомологические коллекции, гербарии , зерновые продукты. В шелководстве уничтожают грону и сильно повреждают коконы тутового шелкопряда . Различные виды точильщиков могут также повреждать пищевые продукты, книги, мебель, деревянные конструкции. А некоторые из них обладают необычным рационом: личинки табачного жука ( Lasioderma serricorne ) питаются сухими листьями табака и являются злостными вредителями табачной промышленности . Хлебный точильщик может питаться даже лекарственными средствами , включая такой яд, как стрихнин .
Личинки крупных плавунцов и сами жуки, а также личинки крупных видов водолюбов , могут, порой, питаться мальками рыб, нанося местами вред рыбному хозяйству .

Крупные представители подсемейства скарабеинов могут являться промежуточными хозяевами ряда гельминтов , в том числе патогенных для домашних животных и иногда человека . В Южной Индии и Шри-Ланке среди местного населения иногда возникает периодическая болезнь кишечника, получившая название « скарабиаз ». Она вызывается некоторыми мелкими видами Onthophagus и Caccobius . Эти жуки проникают в кишечник человека через задний проход , особенно детей, живущих в антисанитарных условиях .
Гемолимфа представителей некоторых семейств содержит ядовитые вещества, в частности кантаридин . Её попадание в организм человека перорально может вызвать серьёзные отравления и даже летальный исход . Особенно ядовита гемолимфа божьих коровок , краснокрылов , малашек и нарывников . Представители последнего семейства могут выделять из сочленений бёдер и голеней капли жёлтой гемолимфы, которая при попадании на кожные покровы вызывает химические ожоги и похожие на нарывы . Также ядовита гемолимфа жуков рода Paederus , обитающих на берегах водоёмов, и которые случайно могут попасть на рыбу , лежащую на месте вылова. Случайное съедание вместе с рыбой такого жука может стать причиной сильного отравления .
Высушенные жуки шпанской мушки ( Lytta vesicatoria ) ранее применялись в аптекарском деле для изготовления т. н. «нарывного пластыря» и использовались в качестве природного афродизиака . Также в медицинских целях применяется спиртовая вытяжка из стафилинидов-синекрылов рода Paederus , повышающая активность клеточного иммунитета . В нетрадиционной медицине для лечения целого ряда заболеваний используется чернотелка . Однако научных доказательств эффективности подобной терапии нет .
|
ḫpr
иероглифами |
||
|---|---|---|
|


В Древнем Египте особо почитался священный скарабей , в Гелиополе считавшийся воплощением бога Атума . Для египтян он олицетворял жизнь и самовозрождение, его название хепри было созвучно глаголу хепер («быть, становиться»). Фигурки скарабеев, сделанные из драгоценных или поделочных камней, служили печатями, амулетами и украшениями. Фигурку скарабея с написанными на нём заклинаниями вкладывали на место сердца при мумификации людей и священных быков . Греческие авторы приписывали египтянам представление, что у скарабеев нет самок, а только самцы, рождающиеся из скатываемого «родителем» навозного шара . Это почитание пародийно преобразуется в комедии Аристофана « Мир », главный герой которой Тригей откармливает навозного жука, чтобы взлететь на небо, к престолу Зевса. При этом сам Зевс иногда почитался в виде жука .
В космогоническом мифе семангов навозный жук собирает землю возле ветви дерева рамбутана , которое воздвигла богиня Манои посреди мирового океана . Амбивалентно воспринимался жук в средневековой ирландской культуре. Например, в гимне святому тот назван «благородным, неописуемым жуком» . Жук выступает как вредоносное, ядовитое существо, ассоциируется с чёрным, но также с чем-то сверкающим, переливающимся (ярко-синего или голубого цвета), неоднократно название жука встречается в составе личных имён и прозвищ. В двух легендах повествуется об огромном жуке-паразите, прогрызающем бок человека (от одного страдает святая Ита, от другого — некий король, исцелённый лекарем Айрмнахом). Привлекательные чёрные брови могут сравниваться со «спинкой жука» .
Как и многим животным необычного вида, жуку-оленю в прошлом иногда приписывали магическую силу. Так, древние римляне носили засушенные головы этих жуков на шее как ожерелье в качестве амулетов , а также надевали его детям для защиты от болезней . В средневековой Англии, этих жуков боялись крестьяне, которые считали, что они могли принести непогоду и погубить урожай .
«Жук на грецком орехе» является одной из традиционных японских нэцкэ . Сам жук при этом символизирует предвестника счастливых событий .
В 1910 году Владислав Александрович Старевич решил снять документальный фильм о жуках-оленях , в частности битву двух самцов-рогачей за самку. Однако выяснилось, что при необходимом для съёмки освещении самцы становятся пассивны. Тогда Старевич придумал сделать из панцирей рогачей муляжи и снять нужную ему сцену покадрово. Снятый им таким образом фильм « Lucanus Cervus » был первым в мире кукольным анимационным фильмом . В той же технике Старевич снял в 1912 году короткометражный фильм « Прекрасная Люканида, или Война усачей с рогачами », в котором жуки разыгрывали сцены, пародирующие сюжеты из рыцарских романов. Фильм пользовался успехом у зрителей.
В последние годы сразу в нескольких мультфильмах в качестве главных или второстепенных героев выступали жуки, например навозник-копр, божья коровка и светляки в мультфильме « Приключения Флика » (1998) . Жуки также являются персонажами французского анимационного телесериала « Minuscule » (2006) : божья коровка показана гиперактивным существом, любящим дразнить других насекомых, особенно мух, и затем быстро удирающей от них, а жук-навозник в нём старается постоянно ухватить шарик навоза побольше. В кинематографе жуки обычно появляются в фильмах ужасов , как наводящие страх существа, в виде нереалистично крупных и свирепых насекомых: в фильмах « Подземная ловушка » ( англ. Caved In: Prehistoric Terror ; 2006) , « Звёздный десант » ( англ. Starship Troopers ; 1997) и другие. В фантастическом фильме ужасов « Мумия » 1999 года показаны плотоядные жуки-скарабеи , нападающие на людей и пожирающие их заживо, что целиком не соответствует действительности (на самом деле они питаются навозом ).
Символом американской рок-группы Journey , образованной в 1973 году , является скарабей , который также изображён (обычно летящим, с расправленными крыльями) на обложках большинства из её альбомов.

Жуки не часто изображаются на полотнах художников. Однако жука-оленя на своих полотнах не анималистической тематики часто изображал Альбрехт Дюрер . Среди многочисленных набросков Леонардо да Винчи имеется рисунок жука-усача . Жуки довольно часто присутствуют на картинах-натюрмортах голландских и фламандских художников эпохи барокко — Яна Давидса де Хема , Яна Брейгеля и Яна Брейгеля младшего , Бальтазара ван дер Аста и , . Немецкий художник Георг Флегель на многих своих натюрмортах изобразил насекомых, в частности жуков. На картинах другого немецкого художника Ханса Хофмана жуки не только изображены, но также фигурируют в их названиях.

Жук-олень упоминается у греческого драматурга Софокла (496—406 до н. э.) — в сатировской драме «Следопыты» говорится о сходстве между лирой и головой жука-оленя .
Жуки-точильщики и связанные с ними «часы смерти» фигурируют в художественных произведениях Марка Твена , Эдгара По , Джорджа Оруэлла , Яна Флеминга , Рэя Брэдбери и других писателей. Джеральд Даррелл в главе «Человек с золотыми бронзовками» своей повести « Моя семья и другие звери » рассказывает о странном чудаке, над головой которого с громким гудением кружились золотисто-зелёные жуки-бронзовки , привязанные к шляпе с помощью длинных ниток. Также Даррелл в своих повествованиях рассказывает о встрече с жуками-голиафами и их гигантскими личинками.
Жуки также встречаются в следующих литературных произведениях: К. Г. Паустовский — «Похождения жука-носорога», Эдгар По — «Золотой жук», Ян Ларри — « Необыкновенные приключения Карика и Вали », Пьер Грипари — «Мадемуазель Скарабей» и многих других. Также майский жук является одним из второстепенных персонажей сказки Ханса Кристиана Андерсена « Дюймовочка ».
Уже среди самых первых монет, отчеканенных в конце VII века до н. э. , присутствуют изображения жуков — на аверсе монеты 1/24 статера 600—550 гг. до н. э. изображён крылатый скарабей . На серебряном оболе , чеканившемся в Аттике в 545—515 гг. до н. э., также изображён скарабей. На современных монетах также имеются изображения жуков. На серебряной монете достоинством 1 новозеландский доллар , выпущенной на Островах Кука , изображена златка .
В 1997 году в Польше была выпущена монета номиналом 2 злотых , изготовленная из бронзы , с изображением жука-оленя. Также существует разновидность этой монеты, изготовленная из серебра, номиналом 20 злотых . В Приднестровье в 2006 году была выпущена серебряная монета достоинством 100 рублей с изображением самца и самки жука-оленя. Стоит заметить, что в надписи на монете сделана ошибка в родовом названии — «Lukanus» вместо «Lucanus» .
Четыре альпийских усача ( Rosalia alpina ) изображены на аверсе серебряной монеты достоинством 500 крон , выпущенной в 2001 году .

В геральдике образ жуков используется крайне редко. Так, водолюбы изображены на гербе французской коммуны в Провансе , майские жуки изображены на муниципальном гербе посёлка ( Чешская Республика ) , на гербе коммуны Бельмон-сюр-Лозанн и гербе муниципалитета Ромаирон ( Швейцария ) , на гербе поселения Розенталь в Нижней Саксонии ( Германия ) .
Жуки являются относительно популярными объектами в современной культуре . Их изображения, в той или иной интерпретации, часто появляются как на обложках научных книг , посвящённых природе , так и художественных произведений; на некоторых рекламных плакатах, на обложках музыкальных дисков , на керамических и ювелирных изделиях , а также почтовых марках .
Сегодня жуки также активно используются в энтомодизайне — оформлении интерьеров жилых и рабочих помещений, профессионально собранными коллекциями насекомых .
В странах Юго-Восточной Азии широко распространено и пользуется популярностью развлечение и азартный вид спорта, называемый « ». Он представляет собой состязание между самцами жуков ( дупляков , реже — рогачей ), хозяева которых натравливают их друг на друга (часто в качестве стимула используется самка). Наблюдатели делают ставки на победу одного из жуков . В Таиланде для этих целей используют самцов , а в Японии популярны поединки между самцами жуков, относящихся как к одному, так и разным видам.
В некоторых странах увлечение содержанием и разведением живых жуков стало распространённым хобби . Особенно широкое распространение оно получило в Японии и на Тайване .





Жуки являются одной из наиболее популярных групп насекомых для коллекционеров. Это обусловлено в первую очередь огромным количеством видов, их распространённостью по всей Земле, а также большим количеством представленных среди них форм, размеров и цветов .
Как и все энтомологические коллекции , их стоит разделять на частные и научные. Частные коллекции в большей мере носят любительский характер и ориентируются на эстетичность, разнообразие и эффектность собранных экземпляров. Жуки в такие коллекции подбираются по цветовой гамме окраски, размерам, разнообразию форм тела, личным предпочтениям и материальным возможностям коллекционеров. В последнее время многие частные коллекции становятся выставочными, принося дополнительный доход их владельцам.
Научные коллекции представлены фондами музеев , университетов, научно-исследовательских институтов и т. д. Такие коллекции собираются научными работниками, которые проводят специализированные фаунистические, эволюционные и экологические исследования жуков. Научные коллекции принято разделять на выставочные — предназначенные для всеобщего обзора — и фондовые, доступ к которым имеют лишь научные сотрудники. Основу фондов этих музеев составляют частные коллекции, завещанные коллекционерами или приобретённые у их наследников .
Крупнейшей на территории России и стран бывшего СССР является коллекция жуков Зоологического института Российской Академии наук . Она включает в себя около 6 млн экземпляров. Начало коллекции было положено в 1714 году Петром I при создании Кунсткамеры .
В результате хозяйственной деятельности человека: вырубки лесов, распашки целинных степей, численность многих видов жуков значительно сократилась, а некоторые из них находятся на грани вымирания. На размерах популяции сказываются изменения в видовом разнообразии растительности мест их обитания. В первую очередь в группе риска находятся жуки, обитающие на ограниченной территории, личинки которых питаются растениями только одного вида .
Научно-директивной основой организации охраны редких и находящихся под угрозой исчезновения видов жесткокрылых служит Красная книга . Во 2-е издание « Красной книги СССР » были включены 32 вида жуков, из которых только 25 видов встречаются на территории России. На сегодняшний день Красная книга Российской Федерации включает 36 видов жуков .
В Красный список угрожаемых видов ( The IUCN Red List of Threatened Species ), публикуемый Международным союзом охраны природы и природных ресурсов (МСОП), внесено 174 редких и исчезающих вида жесткокрылых, из них 12 видов — вымершие , 18 видов — находящиеся на грани исчезновения , 86 видов — вымирающие , 58 видов — уязвимые . Всего на сайте МСОП перечислено 1141 вид жуков с учётом видов, информации о которых недостаточно (Data Deficient, 438 видов), близки к уязвимому положению ( Near Threatened , 73 вида) или чьё положение под наименьшей угрозой ( Least Concern , 456 видов) .
Перечень жесткокрылых, экспорт , реэкспорт и импорт которых регулируется в соответствии с Конвенцией о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения ( СИТЕС ), включает в себя вид Dynastes satanas и род африканских рогачей Colophon .
Привитие широкой общественности современных экологических представлений о необходимости охраны определённых видов жуков просто необходимо. Экологические проекты по охране жесткокрылых должны информировать общественность о важности защиты как самих видов, так и мест их обитания. Только осознание необходимости охраны видов и мест их обитания даст возможность предпринять конкретные действия для решения существующих проблем .
|
Современная
систематика жуков
(
) по подотрядам
|
|
|---|---|
|
|
| † Alphacoleoptera |
|
| Архостематы |
|
| Миксофага | |
| Плотоядные жуки | |
| Разноядные жуки | |
|
|
Эта статья входит в число
избранных статей
русскоязычного раздела Википедии.
|
|
|
Некоторые
внешние ссылки
в этой статье
ведут на сайты, занесённые в
спам-лист
|
| Жесткокрылые | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

Коллекция тропических жуков из Музея Виктории и Альберта (Лондон). |
|||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен:
Царство:
Подцарство:
Без ранга:
Без ранга:
Без ранга:
Без ранга:
Тип:
Подтип:
Надкласс:
Класс:
Подкласс:
Инфракласс:
Надотряд:
Отряд:
Жесткокрылые
|
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Coleoptera Linnaeus , 1758 | |||||||||||||||||||||||||||||||||||||||||
| Подотряды | |||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||
|
Геохронология
появился 318 млн лет
◄
Наше время
◄
Мел-
палеогеновое вымирание
◄
Триасовое вымирание
◄
Массовое
пермское вымирание
◄
Девонское вымирание
◄
Ордовикско-силурийское вымирание
◄
Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||