Interested Article - Азотобактер

- 2020-04-27
- 1
Азотоба́ктер ( лат. Azotobacter ) — род бактерий , живущих в почве и способных в результате процесса азотфиксации переводить газообразный азот в растворимую форму, доступную для усваивания растениями .
Род азотобактер принадлежит к грамотрицательным бактериям и входит в группу так называемых свободноживущих азотфиксаторов. Представители рода обитают в нейтральных и щелочных почвах , воде и в ассоциации с некоторыми растениями . Образуют особые покоящиеся формы — цисты .
Играет важную роль в круговороте азота в природе, связывая недоступный растениям атмосферный азот и выделяя связанный азот в виде ионов аммония в почву. Используется человеком для производства азотных биоудобрений, является продуцентом некоторых биополимеров .
Первый представитель рода, Azotobacter chroococcum , был открыт и описан в 1901 году голландским микробиологом и ботаником Мартином Бейеринком . На данный момент в род входят шесть видов.
Биологические свойства
Морфология
Клетки бактерий рода Azotobacter относительно крупные (1—2 мкм в диаметре), обычно овальные, но обладают , то есть могут иметь разную форму — от палочковидной до сферической . На микроскопических препаратах клетки могут располагаться одиночно, парами, неправильными скоплениями или, изредка, цепочками различной длины. Формируют особые покоящиеся формы — цисты , не образуют спор .
В свежих культурах клетки подвижны за счёт многочисленных жгутиков . В более поздних культурах клетки теряют подвижность, приобретают почти кокковидную форму и продуцируют толстый слой слизи , формирующий капсулу клетки. На форму клетки также оказывает влияние химический состав питательной среды — пептон , например, вызывает плеоморфизм и, в том числе, индуцирует образование так называемых «грибоподобных» клеток. Индуцирующее влияние на плеоморфизм в культурах представителей рода Азотобактер в составе пептона оказывает аминокислота глицин .
При микроскопии в клетках наблюдаются включения, часть из которых окрашивается, а часть остаётся бесцветными. В начале XX века считалось, что прокрашиваемые включения являются «репродуктивными гранулами», или , и принимают участие в размножении клетки, являясь своеобразными « зародышевыми » клетками , однако затем было доказано, что гранулы не принимают участия в размножении клеток и не являются «малыми, коккоподобными репродуктивными клетками» бактерий — гонидиями . Прокрашиваемые гранулы состоят из волютина , неокрашивающиеся же гранулы являются каплями жира. Гранулы являются резервным источником питания.
Цисты
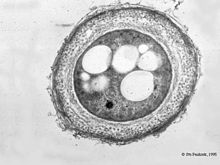
Цисты представителей рода Azotobacter более устойчивы к действию неблагоприятных факторов внешней среды, чем вегетативные клетки — так, цисты в два раза более устойчивы к действию ультрафиолетового излучения чем вегетативные клетки, устойчивы к высушиванию, гамма-излучению , солнечной иррадиации, действию ультразвука , однако не являются устойчивыми к действию высоких температур.
Формирование цист индуцируется изменением концентрации питательных веществ в питательной среде и добавлением некоторых органических веществ (например, этанола , н -бутанола и β-гидроксибутирата). Цисты редко образуются в жидких питательных средах . Инцистирование может быть индуцированно химическими факторами и сопровождается метаболическими сдвигами, изменениями в катаболизме и дыхании, изменениями в биосинтезе макромолекул . Определённое значение в индукции инцистирования имеет альдегиддегидрогеназа , а также регулятор ответа AlgR .
Циста азотобактера — сферическое тело, состоящее из так называемого центрального тела (уменьшенной копии вегетативной клетки с большим количеством вакуолей ) и двуслойной оболочки , внутренняя часть которой называется интима и имеет волокнистое строение , а внешняя называется экзина и представлена ровной, отражающей структурой, имеющей гексагональное кристаллическое строение . Экзина частично гидролизуется трипсином и устойчива к действию лизоцима , в отличие от центрального тела . Центральное тело может быть изолировано в жизнеспособном состоянии некоторыми хелатирующими агентами . Главными компонентами внешней оболочки цисты являются алкилрезорцинолы, состоящие из длинных алифатических цепей и ароматических колец. Алкилрезорцинолы встречаются также у других бактерий, животных и растений .
Прорастание цист
Циста представителей рода Azotobacter является покоящейся формой вегетативной клетки, необходимой для переживания неблагоприятных факторов внешней среды, и не служит для размножения . После возобновления оптимальных условий, таких, как оптимальное значение pH, температуры и поступления доступного источника углерода , цисты прорастают, образовавшаяся вегетативная клетка вновь размножается путём простого деления клетки . При прорастании цист экзина цисты повреждается, и высвобождается большая вегетативная клетка.
Микроскопически первым проявлением прорастания спор является постепенное понижение преломления света цистами при фазово-контрастной микроскопии . Прорастание цист — медленный процесс и длится около 4—6 часов, на протяжении которых центральное тело увеличивается и происходит захват гранул волютина , прежде находившихся в интиме. Затем экзина лопается и вегетативная клетка высвобождается из экзины, имеющей характерную подковообразную форму . При прорастании цисты отмечаются метаболические изменения. Сразу после прибавления источника углерода к среде цисты начинают поглощать кислород и выделять двуокись углерода , скорость дыхания повышается до максимальных значений через 4 часа после прибавления глюкозы . Синтез белков и РНК также начинается после прибавления источника углерода к среде, однако интенсификация синтеза макромолекул отмечается лишь через 5 часов после прибавления источника углерода. Синтез ДНК и фиксация азота инициируются через 5 часов после прибавления глюкозы к безазотистой питательной среде .
Во время прорастания цист отмечаются изменения в интиме, видимые на электронно-микроскопических препаратах. Интима состоит из углеводов , липидов и белков и занимает почти такой же объём в клетке, что и центральное тело. Во время прорастания цист интима гидролизируется и используется клеткой для синтеза компонентов клетки .
Физиологические свойства
Получают энергию в ходе окислительно-восстановительных реакций, используя в качестве донора электронов органические соединения. Для роста необходим кислород, но способны расти при пониженных концентрациях кислорода , образуют каталазу и . Способны использовать различные углеводы , спирты и соли органических кислот в качестве источников углерода . способны фиксировать по крайней мере 10 мкг азота на грамм потреблённой глюкозы , фиксация азота зависит от наличия ионов молибдена , отсутствие молибдена может быть частично замещено ионами ванадия . В качестве источников азота могут использовать нитраты , ионы аммония и аминокислоты . Оптимум pH для роста и фиксации азота 7,0—7,5, способны расти в диапазоне pH от 4,8 до 8,5. Возможен также зависимый от водорода миксотрофный рост представителей рода Azotobacter на безазотистой питательной среде, содержащей маннозу . Водород доступен в почве, поэтому не исключена возможность миксотрофии у представителей рода Azotobacter в природных условиях.
Культуральные свойства
Представители рода Azotobacter способны использовать углеводы (например маннит , сахарозу , глюкозу ), спирты (в том числе этанол и бутанол) и соли органических кислот, в том числе и бензоаты , в качестве источника углерода и энергии. Представители рода растут на безазотистых средах, предназначенных для выделения свободноживущих азотфиксирующих и олигонитрофильных организмов, например на , содержащей источник углерода (маннит, сахароза или глюкоза) и необходимые микроэлементы (источник фосфора , серы и т. д.), или на , содержащей больше микроэлементов , а также на жидкой .
На плотных питательных средах представители рода образуют плоские, слизистые колонии пастообразной консистенции диаметром 5—10 мм, в жидких питательных средах образуют плёнки. Характерно также пигментирование , колонии представителей рода могут быть окрашены в тёмно-коричневый, зелёный и других цветов, или же могут быть бесцветными в зависимости от видовой принадлежности. Представители рода Azotobacter являются мезофильными микроорганизмами и растут при температуре 20—30 °C.
Пигменты
Представители рода Azotobacter продуцируют пигменты. Например, типовой вид рода Azotobacter chroococcum продуцирует тёмно-коричневый водорастворимый пигмент (в видовом эпитете как раз отражена эта способность) меланин . Продукция меланина у Azotobacter chroococcum наблюдается при высоких уровнях дыхания во время фиксации азота и, предположительно, также защищает нитрогеназную систему от действия кислорода при аэроадаптации Другие виды рода Azotobacter также продуцируют пигменты от жёлто-зелёного до пурпурного цвета. Также представители рода способны продуцировать зеленоватый флюоресцирующий пигмент, флюоресцирующий жёлто-зелёным светом и пигмент, флюоресцирующий бело-голубым светом.
Геном
Частично завершено определение нуклеотидной последовательности хромосомы Azotobacter vinelandii штамма AvOP. Хромосома Azotobacter vinelandii — кольцевая молекула ДНК размером 5 342 073 пар нуклеотидов и содержит 5043 генов , из которых 4988 кодируют белки , составляет 65 %. Отмечено изменение плоидности представителей рода Azotobacter на протяжении жизненного цикла: по мере старения культур количество хромосом в клетках и содержание ДНК увеличивается — в стационарной фазе роста культуры могут содержать более 100 копий хромосомы на клетку. При пересеве на свежую питательную среду первоначальное содержание ДНК (одна копия) восстанавливается Кроме хромосомальной ДНК, у представителей рода Azotobacter обнаружены плазмиды , доказана и возможность трансформации представителей рода Azotobacter экзогенной плазмидной ДНК .
Распространение
Представители рода Azotobacter распространены повсеместно в нейтральных и слабощелочных почвах и не выделяются из кислых почв. Были они обнаружены и в экстремальных условиях почв северного и южного полярного региона , несмотря на короткие местные сезоны роста и относительно низкие значения pH, — в арктическом регионе в глине и суглинках (в том числе торфянистых и песчанистых суглинках), в антарктическом регионе — в грунте побережья В сухих почвах представители этого рода способны сохраняться в виде цист до 24 лет.
Также представители рода Azotobacter были выделены из водных местообитаний, в том числе из пресноводных водоёмов , солоноватоводных болот . Некоторые представители рода Azotobacter ассоциированы с растениями и обнаружены в ризосфере , вступая с растением в определённые взаимоотношения — представители рода были выделены из ризосферы мангровых деревьев совместно с другими азотфиксирующими и денитрифицирующими бактериями .
Некоторые штаммы также обнаружены в коконах дождевых червей Eisenia fetida .
Фиксация азота
Представители рода Azotobacter являются свободноживущими азотфиксаторами, то есть в отличие от представителей рода Rhizobium фиксируют молекулярный азот из атмосферы , не вступая в симбиотические отношения с растениями, хотя некоторые представители рода вступают в ассоциацию с растением-хозяином. Фиксация азота ингибируется наличием доступных источников азота, например ионов аммония , нитратов .
Представители рода Azotobacter имеют полный комплекс ферментов , необходимый для осуществления азотфиксации: ферредоксины , гидрогеназы и важнейший фермент — нитрогеназу . Процесс азотфиксации энергозависим и требует притока энергии в виде АТФ . Процесс фиксации азота крайне чувствителен к присутствию кислорода , поэтому у представителей рода Azotobacter выработался особый механизм защиты от действия кислорода — так называемая дыхательная защита, осуществляемая путём значительной интенсификации дыхания, снижающего концентрацию кислорода в клетках. Также имеется особый белок Shethna, защищающий нитрогеназу и участвующий в предотвращении гибели клетки, вызванной кислородом: мутанты, не вырабатывающие этот белок, гибнут в присутствии кислорода во время азотфиксации в отсутствие источника азота в среде Определённую роль в процессах азотфиксации у Azotobacter играют -ионы.
Нитрогеназы
Нитрогеназный комплекс является важнейшим ферментом , участвующим в азотфиксации. У представителей рода Azotobacter обнаружено несколько типов нитрогеназ — Mo - Fe - нитрогеназа и альтернативные нитрогеназы: Ванадий -содержащая, не зависимая от ионов молибдена , более активная чем Mo-Fe-нитрогеназа в условиях пониженных температур — так, эффективная фиксация азота не прекращалась V-нитрогеназой вплоть до понижения температуры до 5 °C, активность V-нитрогеназы понижалась при понижении температуры в 10 раз меньше, чем у Mo-Fe-нитрогеназы , и Fe -содержащая, менее активная, чем обычная нитрогеназа . Важную роль в образовании активной нитрогеназы играет созревание Р- кластера Mo-Fe-нитрогеназы , также как и предшественник Mo-Fe- кофактора нитрогеназы , шаперон GroEL, играет важную роль в завершающей перестройке нитрогеназы . Регуляция активности нитрогеназы может осуществляться образованием осадка аргинина Синтез нитрогеназы осуществляется под контролем т. н. nif-генов. Фиксация азота регулируется nifLA опероном , продукт NifA регулирует транскрипцию nif-генов, NifL имеет антагонистичное действие по отношению к действию NifA в ответ на поглощённый азот и в зависимости от уровня поступления кислорода в клетку, экспрессия nifLA оперона регулируется по механизму позитивной регуляции. NifL является флавопротеином , модулирующим активацию транскрипции генов азотфиксации путём редокс -зависимого переключения. Двухкомпонентная система регуляции, состоящая из двух белков ( NifA и сенсора NifL), образующих комплексы между собой, является атипичной и не распространённой среди других организмов системой регуляции экспрессии генов.
Значение
Азотфиксация играет большую роль в круговороте азота в природе. Азотфиксация является важнейшим источником азота, и представители рода Azotobacter играют важнейшую роль в круговороте азота почвы, осуществляя фиксацию молекулярного азота. Также представители рода синтезируют некоторые биологически активные вещества, в том числе и некоторые фитогормоны , например ауксины , тем самым стимулируя рост и развитие растений , являясь биологическим стимулятором роста растений и синтезируя факторы, необходимые для роста растений . Экзополисахариды представителей рода способствуют мобилизации тяжёлых металлов в почве, способствуя самоочищению почв, загрязнённых тяжёлыми металлами, например кадмием , ртутью и свинцом . Некоторые представители рода Azotobacter также способны к биодеградации некоторых хлорсодержащих ароматических соединений , например ( ) — ранее использовавшегося инсектицида , фунгицида и гербицида , имеющего мутагенное и канцерогенное действие и являющегося ксенобиотиком и поллютантом .
Использование человеком
Благодаря своей способности фиксировать молекулярный азот, тем самым повышая плодородие почвы и стимулирования роста растений представители рода Azotobacter используются в сельском хозяйстве для получения азотных биоудобрений, в том числе , также представители рода являются продуцентами полисахарида — альгиновой кислоты (E400) , использующегося в медицине (в качестве антацида ), в пищевой промышленности (в качестве пищевой добавки к мороженому, пудингам и кремам) и в биосорбции металлов и поли(3-гидроксибутирата) ( ) . является продуцентом рестриктазы Abe I, узнающей несимметричную гептануклеотидную последовательность CCTCAGC.
Систематика
Род Azotobacter был описан в 1901 году голландским микробиологом и ботаником, одним из основоположников экологической микробиологии Мартином Бейеринком на основании впервые выделенного и описанного им Azotobacter chroococcum , первого аэробного свободноживущего азотфиксатора.

(1851—1931),
первооткрыватель бактерий рода Azotobacter
В 1903 году Липман ( Lipman ) описал Azotobacter vinelandii Lipman, 1903 , а годом позже Lipman, 1904 , названный им в честь самого Мартина Бейеринка. В 1949 году русский микробиолог Николай Александрович Красильников описал вид Krasil'nikov, 1949 , в 1981 году разделённый Томпсоном ( Thompson ) и Скирманом ( Skerman ) на два подвида : Azotobacter nigricans subsp. nigricans Krasil'nikov, 1949 и Azotobacter nigricans subsp. achromogenes Thompson and Skerman, 1981 , в том же году Томпсон и Скирман описали вид Thompson and Skerman, 1981 . В 1991 Пейдж ( Page ) и Шивпрасад ( Shivprasad ) описали микроаэрофильный , зависимый от ионов натрия аэротолерантный вид Azotobacter salinestris Page and Shivprasad 1991 .
Ранее представители рода принадлежали к семейству Azotobacteraceae Pribram, 1933 , но затем были перенесены в семейство Pseudomonadaceae на основании изучения нуклеотидных последовательностей 16S рРНК . В 2004 году было проведено филогенетическое исследование и выяснено, что Azotobacter vinelandii входит в одну кладу с бактерией Pseudomonas aeruginosa . В 2007 году было сделано предположение о близости родов Azotobacter , и и о возможной синонимичности .
- Таксономическая схема
| царство Бактерии | ||||||||||||||||||||||
| тип Протеобактерии | ещё более двадцати типов, в том числе Актиномицеты , Фирмикуты , Цианобактерии , Aquificae , Dictyoglomi | |||||||||||||||||||||
| класс Gamma Proteobacteria | классы (род Риккетсии и др.), (род Neisseria и др.), (роды , Desulfovibrio и др.), | |||||||||||||||||||||
| порядок | ещё около пятнадцати порядков, в том числе Enterobacteriales (роды Иерсинии , Сальмонеллы , Эрвинии и др.), (род Alcanivorax и др.), ( Гемофильная палочка и др.) | |||||||||||||||||||||
| семейство Pseudomonadaceae | семейство | |||||||||||||||||||||
| род Азотобактер | ещё около пятнадцати родов | |||||||||||||||||||||
| шесть видов | ||||||||||||||||||||||
К роду Azotobacter ранее принадлежали также виды Azotobacter agilis (перенесён в 1938 году Виноградским в род ), Azotobacter macrocytogenes (перенесён в 1981 году в род и в 1982 году в род ) и Azotobacter paspali (перенесён в 1981 году в род ).
См. также
Примечания
- Азотобактер // А — Ангоб. — М. : Советская энциклопедия, 1969. — ( Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 1).
- Gandora V. , Gupta R. D. , Bhardwaj K. K. R. // Journal of the Indian Society of Soil Science. — 1998. — Т. 46 , № 3 . — С. 379—383 . — ISSN . 6 сентября 2012 года. CODEN JINSA4
- Martyniuk S., Martyniuk M. // Polish Journal of Environmental Studies. — 2003. — Т. 12 , № 3 . — С. 371—374 . 15 июля 2011 года.
- Tejera N., Lluch C., Martínez-Toledo M. V. , González-López J. // Plant and Soil. — 2005. — Т. 270 , № 1—2 . — С. 223—232 . — ISSN . 3 марта 2016 года.
- Kumar R., Bhatia R., Kukreja K., Behl R. K. , Dudeja S. S. , Narula N. // Journal of Basic Microbiology. — 2007. — Т. 47 , № 5 . — С. 436—439 . 4 апреля 2020 года.
- Baillie A., Hodgkiss W., Norris J. R. // Journal of Applied Microbiology. — 1962. — Т. 25 , № 1 . — С. 116—119 . 4 апреля 2020 года.
- Vela G. R. , Rosenthal R. S. // Journal of Bacteriology. — 1972. — Т. 111 , № 1 . — С. 260—266 .
- Jones D. H. // Journal of Bacteriology. — 1920. — Т. 5 , № 4 . — С. 325—341 .
- Lewis I. M. // Bacteriological Reviews. — 1941. — Т. 5 , № 3 . — С. 181—230 .
- Lewis I. M. // Journal of Bacteriology. — 1937. — Т. 34 , № 2 . — С. 191–205 .
- Socolofsky M. D. , Wyss O. // Journal of Bacteriology. — 1962. — Т. 84 . — С. 119—124 .
- Layne J. S. , Johnson E. J. // Journal of Bacteriology. — 1964. — Т. 87 , № 3 . — С. 684—689 .
- Sadoff H. L. // Microbiological Reviews. — 1975. — Т. 39 , № 4 . — С. 516—539 .
- Gama-Castro S., Núñez C., Segura D. , Moreno S., Guzmán J., and Espín G. // Journal of Bacteriology. — 2001. — Т. 183 , № 21 . — С. 6169—6174 .
- Núñez C., Moreno S., Soberón-Chávez G. , Espín G. // Journal of Bacteriology. — 1999. — Т. 181 , № 1 . — С. 141–148 .
- Pope L. M. , Wyss O. // Journal of Bacteriology. — 1970. — Т. 102 , № 1 . — С. 234—239 .
- Page W. J. , Sadoff H. L. // Journal of Bacteriology. — 1975. — Т. 122 , № 1 . — С. 145—151 .
- Lin L. P. , Sadoff H. L. // Journal of Bacteriology. — 1969. — Т. 98 , № 3 . — С. 1335—1341 .
- Parker L. T. , Socolofsky M. D. // Journal of Bacteriology. — 1968. — Т. 91 , № 1 . — С. 297—303 .
- Funa N., Ozawa H., Hirata A., Horinouchi S. // Proceedings of the National Academy of Sciences of the United States of America : Сб.. — 2006. — Т. 103 , № 16 . — С. 6356–6361 .
- Wyss O., Neumann M. G. , Socolofsky M. D. // Journal of Biophysical and Biochemical Cytology. — 1961. — № 10 . — С. 555—565 . 20 ноября 2008 года.
- Loperfido B., Sadoff H. L. // Journal of Bacteriology. — 1973. — Т. 112 , № 2 . — С. 841—846 .
- Lin L. P. , Pankratz S., Sadoff H. L. // Journal of Bacteriology. — 1978. — Т. 135 , № 2 . — С. 641—646 .
- Part B: The Gammaproteobacteria // / Editor-in-Chief: George M. Garrity. — 2nd Edition. — New York: Springer, 2005. — 2816 p. — ISBN 0-387-95040-0 . 31 января 2010 года.
- Wong T.-Y., Maier R. J. // Journal of Bacteriology. — 1985. — Т. 163 , № 2 . — С. 528–533 .
- Большой практикум по микробиологии / Под общей ред. проф. Г. Л. Селибера. — М. : Высшая школа, 1962. — С. 190—191.
- Теппер Е. З., Шильникова В. К., Переверзева Г. И. Практикум по микробиологии. — 2-е изд., перераб. и доп. — М. : Колос, 1979. — 216 с.
- Shivprasad S., Page W. J. // Applied and Environmental Microbiology. — 1989. — Т. 55 , № 7 . — С. 1811–1817 .
- Jensen H. L. // Bacteriological Reviews. — 1954. — Т. 18 , № 4 . — С. 195–214 .
- Johnstone D. B. // Journal of Bacteriology. — 1955. — Т. 69 , № 4 . — С. 481–482 .
- . Дата обращения: 25 ноября 2019. 22 марта 2022 года.
- Maldonado R., Jimenez J., Casadesus J. // Journal of Bacteriology. — 1994. — Т. 176 , № 13 . — С. 3911—3919 .
- Maia M., Sanchez J. M., Vela G. R. // Journal of Bacteriology. — 1988. — Т. 170 , № 4 . — С. 1984—1985 .
- Glick B. R., Brooks H. E., Pasternak J. J. // Journal of Bacteriology. — 1985. — Т. 162 , № 1 . — С. 276—279 .
- Yamagata U., Itano A. // Journal of Bacteriology. — 1923. — Т. 8 , № 6 . — С. 521—531 .
- Boyd W. L., Boyd J. W. // Journal of Bacteriology. — 1962. — Т. 83 , № 2 . — С. 429–430 .
- Moreno J., Gonzalez-Lopez J., Vela G. R. // Applied and Environmental Microbiology. — 1986. — Т. 51 , № 1 . — С. 123—125 .
- Johnstone D. B. // Ecology. — 1967. — Т. 48 , № 4 . — С. 671—672 .
- Dicker H. J., Smith D. W. // Applied and Environmental Microbiology. — 1980. — Т. 39 , № 5 . — С. 1019—1025 .
- van Berkum P., Bohlool B. // Microbiological Reviews. — 1980. — Т. 44 , № 3 . — С. 491—517 .
- Flores-Mireles A. L., Winans S. C., Holguin G. // Applied and Environmental Microbiology. — 2007. — Т. 73 , № 22 . — С. 7308–7321 .
- Zachmann J. E., Molina J. A. E. // Applied and Environmental Microbiology. — 1993. — Т. 59 , № 6 . — С. 1904—1910 .
- Kass D. L., Drosdoff M., Alexander M. // Soil Science Society of America Journal. — 1971. — № 35 . — С. 286—289 . 24 июля 2008 года.
- Bürgmann H., Widmer F., Sigler W. V, Zeyer J. // Applied and Environmental Microbiology. — 2003. — Т. 69 , № 4 . — С. 1928—1935 .
- Берцова Ю. В., Демин О. В., Богачев А. В. // Успехи биологической химии : Сб.. — 2005. — Т. 45 . — С. 205—234 . 22 июля 2011 года.
- Maier R. J., Moshiri F. // Journal of Bacteriology. — 2000. — Т. 182 , № 13 . — С. 3854—3857 .
- Durrant M. C., Francis A., Lowe D. J., Newton W. E., Fisher K. // Biochemistry Journal. — 2006. — Т. 397 , № 2 . — С. 261–270 .
- Howard J. B., Rees D. C. // Proceedings of the National Academy of Sciences of the United States of America. — 2006. — Т. 103 , № 46 . — С. 17088–17093 .
- Bellenger J. P., Wichard T., Kraepiel A. M. L. // Applied and Environmental Microbiology. — 2008. — Т. 74 , № 5 . — С. 1478–1484 .
- Rüttimann-Johnson C., Rubio L. M., Dean D. R., Ludden P. W. // Journal of Bacteriology. — 2003. — Т. 185 , № 7 . — С. 2383–2386 .
- Robson R. L., Eady R. R., Richardson T. H., Miller R. W., Hawkins M., Postgate J. R. // Nature. — 1986. — № 322 . — С. 388—390 . 12 декабря 2008 года.
- Miller R. W., Eady R. R. // Biochemistry Journal. — 1988. — Т. 256 , № 2 . — С. 429–432 .
- Fallik E., Chan Y.-K., Robson R. L. // Journal of Bacteriology. — 1991. — Т. 173 , № 1 . — С. 365—371 .
- Pau R. N., Mitchenall L. A., Robson R. L. // Journal of Bacteriology. — 1989. — Т. 171 , № 1 . — С. 124–129 .
- Hu Y., Fay A. W., Lee C. C., Ribbe M. W. // Proceedings of the National Academy of Sciences of the United States of America. — 2007. — Т. 104 , № 25 . — С. 10424–10429 .
- Hu Y., Fay A. W., Lee C. C., Ribbe M. W. // Proceedings of the National Academy of Sciences of the United States of America. — 2005. — Т. 102 , № 9 . — С. 3236—3241 .
- Ribbe M. W., Burgess B. K. // Proceedings of the National Academy of Sciences of the United States of America. — 2001. — Т. 98 , № 10 . — С. 5521—5525 .
- Martinez-Argudo I., Little R., Dixon R. // Proceedings of the National Academy of Sciences of the United States of America. — 2004. — Т. 101 , № 46 . — С. 16316—16321 .
- Curatti L., Brown C. S., Ludden P. W., Rubio L. M. // Proceedings of the National Academy of Sciences of the United States of America. — 2005. — Т. 102 , № 18 . — С. 6291—6296 .
- Mitra R., Das H. K., Dixit A. // Applied and Environmental Microbiology. — 2005. — Т. 71 , № 7 . — С. 3716—3724. .
- Hill S., Austin S., Eydmann T., Jones T., Dixon R. // Proceedings of the National Academy of Sciences of the United States of America. — 1996. — Т. 93 , № 5 . — С. 2143—2148 .
- Money T., Barrett J., Dixon R., Austin S. // Journal of Bacteriology. — 2001. — Т. 183 , № 4 . — С. 1359—1368 .
- Ahmad F., Ahmad I., Khan M. S. // Turkish Journal of Biology. — 2005. — № 29 . — С. 29—34 . 15 апреля 2010 года.
- Oblisami G., Santhanakrishan P., Pappiah C. M., Shabnugavelu K. G. // Acta Horticulturae (ISHS). — № 108 . — С. 44—49 . 2 декабря 2008 года.
- Rajaee S., Alikhani H. A., Raiesi F. // Journal of Science and Technology of Agriculture and Natural Resources. — 2007. — Т. 11 , № 41 . — С. 297 . (недоступная ссылка)
- Chen J. H., Czajka D. R., Lion L. W., Shuler M. L., Ghiorse W. C. // Environmental Health Perspectives. — 1995. — Т. 103 , № 1 . — С. 53—58 .
- Li D. Y., Eberspächer J., Wagner B., Kuntzer J., Lingens F. // Applied and Environmental Microbiology. — 1991. — Т. 57 , № 7 . — С. 1920—1928 .
- / edited by Neeru Narula. — New Delhi, 2000. — 162 p. — ISBN 81-239-0661-7 . 20 ноября 2008 года.
- Волова Т. Г. 6.3. Биологические удобрения // / Под ред. академика И. И. Гительзона. — Новосибирск: Издательство СО РАН, 1999. — С. 190—193. — ISBN 5-7692-0204-1 . (недоступная ссылка)
- Galindo E., Peña C., Núñez C., Segura D., Espín G. // Microbial Cell Factories. — 2007. — Т. 6 , № 7 .
- Page W. J., Tindale A., Chandra M., Kwon E. // Microbiology. — 2001. — № 147 . — С. 483—490 . 17 февраля 2009 года.
- Ahmed M., Ahmed N. // Current Genomics. — 2007. — Т. 8 , № 3 . — С. 191–202 .
- Emtiazia G., Ethemadifara Z., Habibib M. H. // African Journal of Biotechnology. — 2004. — Т. 3 , № 6 . — С. 330—333 . 22 июля 2011 года.
- Pettinari M. J., Vázquez G. J., Silberschmidt D., Rehm B., Steinbüchel A., Méndez B. S. // Applied and Environmental Microbiology. — 2001. — Т. 67 , № 11 . — С. 5331—5334 .
- Vitkute J., Maneliene Z., Janulaitis A. -CCTCAGC-3[prime](-/-2)] // Nucleic Acids Research. — 1998. — Т. 26 , № 21 . — С. 4917—4918 . 18 января 2006 года.
- Beijerinck M. W. Ueber Oligonitrophile Mikroben // Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene. Abteilung II. — 1901. — № 7 . — С. 561—582 .
- Page W. J., Shivprasad S. // International Journal of Systematic Bacteriology. — 1991. — Т. 41 , № 3 . — С. 369—376 . (недоступная ссылка)
- Rediers H., Vanderleyden J., De Mot R. // Microbiology. — 2004. — № 150 . — С. 1117—1119 . 15 октября 2008 года.
- Young J. M., Park D.-C. // International Journal of Systematic and Evolutionary Microbiology. — 2007. — № 57 . — С. 2894—2901 . 24 июля 2008 года.
Ссылки
- (англ.) . MicrobeWiki. — The student-edited microbiology resource. Дата обращения: 13 сентября 2008. 20 августа 2011 года.
- J. P. Euzéby. (англ.) . List of Prokaryotic names with Standing in Nomenclature . Дата обращения: 13 сентября 2008. 20 августа 2011 года.
- (англ.) . — Сайт проекта по исследованию генома Azotobacter vinelandii. Дата обращения: 13 сентября 2008. 20 августа 2011 года.
- Amy Crum. (англ.) . SOIL MICROBIOLOGY BIOL/CSES 4684 . Дата обращения: 13 сентября 2008. 23 января 2012 года.
- (англ.) . John Innes Centre — Molecular Microbiology Department. Дата обращения: 13 сентября 2008. 20 августа 2011 года.
- (англ.) . JGI. Дата обращения: 13 сентября 2008. 20 августа 2011 года.
- Iwao WATANABE (JICA/Cantho Univ. expert Mar-Apr. 2000). (англ.) . Lecture in Cantho University, Vietnam (30 марта 2000). Дата обращения: 13 сентября 2008. 20 августа 2011 года.
- (англ.) . Microbiology Video Library . MicrobiologyBytes. Дата обращения: 13 сентября 2008. 20 августа 2011 года.
- 2020-04-27
- 1
| Азотобактер | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|

Клетки представителей рода Azotobacter : видны палочковидные и кокковидные клетки различного размера. Окраска железным гематоксилином по Гейденгайну; ×1000 |
||||||||||
| Научная классификация | ||||||||||
|
Домен:
Тип:
Класс:
Порядок:
Семейство:
Род:
Азотобактер
|
||||||||||
| Международное научное название | ||||||||||
| Azotobacter Beijerinck 1901 | ||||||||||
| Виды | ||||||||||
|
||||||||||