Interested Article - Метаботропные рецепторы

- 2020-08-04
- 1
Метаботропные рецепторы — это подтип трансмембранных рецепторов в эукариотических клетках , воздействие на которые приводит к первичным, непосредственным изменениям метаболизма в клетке (откуда, собственно, и название «метаботропные» рецепторы). Эффект всех без исключения метаботропных рецепторов опосредуется через те или иные системы вторичных посредников . Метаботропные рецепторы могут быть расположены как на поверхностной мембране клетки, так и на мембранах внутриклеточных везикул.
На основании их структурных и функциональных характеристик, все рецепторы к нейромедиаторам и гормонам могут быть разделены на две большие категории: метаботропные и ионотропные рецепторы. Ионотропные рецепторы непосредственно связаны с ионными каналами клетки. А метаботропные рецепторы не имеют прямой связи с ионными каналами на поверхности клетки или её везикул и осуществляют передачу сигнала на них через те или иные внутриклеточные сигнальные механизмы, нередко — через G-белки . В частности, все G-белок-связанные рецепторы по определению являются метаботропными. Другие известные примеры метаботропных рецепторов включают в себя рецепторные тирозинкиназы и гуанилатциклазы.
Оба типа рецепторов — метаботропные и ионотропные — активируются при связывании с ними специфического лиганда — или экзогенного агониста , обычно нейромедиатора или гормона. Когда активируется ионотропный рецептор, он открывает ионный канал, который позволяет вход внутрь клетки или, напротив, выход из клетки ионов , таких, как ионы натрия , калия , кальция или хлора , что приводит к изменениям электрического потенциала клетки. Когда же активируется метаботропный рецептор, активируются связанные с ним внутриклеточные сигнальные каскады (системы вторичных посредников) и запускается цепочка тех или иных внутриклеточных событий, которые могут приводить или не приводить к открытию или закрытию ионных каналов клетки.
Примеры
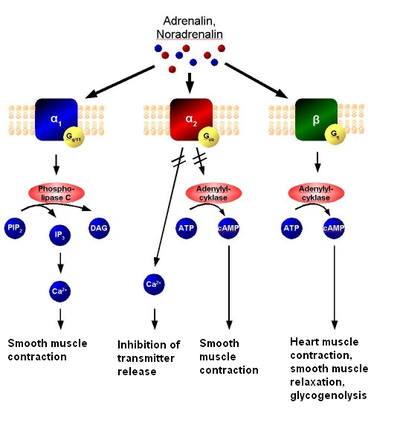
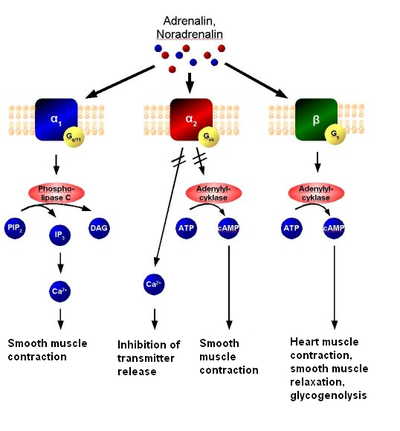
Этот класс рецепторов включает в себя метаботропные глутаматные рецепторы, мускариновые ацетилхолиновые рецепторы, ГАМК -Б рецепторы, и большинство подтипов серотониновых рецепторов (за исключением 5-HT 3 -рецептора , который является ионотропным), а также рецепторы к норадреналину , адреналину , гистамину , дофамину , различным нейропептидам , гормонам, факторам роста, эндогенным опиоидам ( эндорфинам ) и каннабиноидам .
Структура
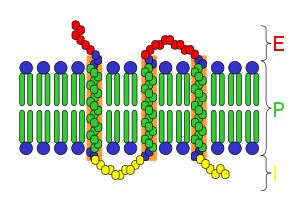
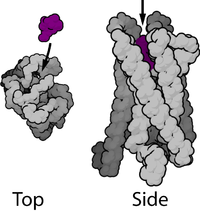
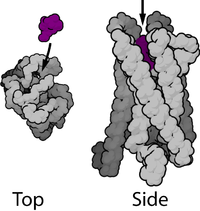
Наиболее распространённый тип метаботропных рецепторов — G-белок-связанные рецепторы — имеют семь гидрофобных трансмембранных доменов. Большинство из них являются (или могут являться) мономерными белками, хотя ГАМК-Б рецепторы требуют гетеродимеризации для своего правильного функционирования. N-терминальный конец белка при этом расположен во внеклеточном пространстве, а C-терминальный конец — во внутриклеточном.
Эти семь трансмембранных доменов, с наружным N-терминальным концом, часто имеют (или предполагается, что имеют) α-спиральную структуру. Полипептидная цепочка типичного G-белок-связанного рецептора обычно состоит из приблизительно 450—550 аминокислотных оснований. Эти белки часто подвергаются гликозилированию , а также пальмитированию . Пальмитирование обеспечивает повышение их гидрофобности и локализацию их на поверхности мембраны клетки в специфических местах.
Функция
Эндогенными лигандами (агонистами) метаботропных рецепторов являются те или иные нейромедиаторы или гормоны, которые, связываясь с рецептором, изменяют его пространственную конфигурацию, что приводит к его активации и запуску тех или иных внутриклеточных сигнальных каскадов (причём связанных с одним и тем же рецептором сигнальных каскадов может быть несколько, то есть более одного). Это, в свою очередь, может приводить к открытию или закрытию тех или иных ионных каналов и к другим изменениям в метаболизме и жизнедеятельности клетки, например к изменению транскрипции генов , изменениям в фосфорилировании тех или иных белков. Когда лиганд-агонист (нейромедиатор или гормон) связывается с белком рецептора (первичным передатчиком сигнала), этот белок-рецептор активирует внутриклеточные первичные эффекторные белки (например, G-белок и через него так называемую гормончувствительную аденилатциклазу ). Это, в свою очередь, приводит к образованию так называемых вторичных посредников (например, цАМФ ) и к активации вторичных эффекторных белков (например, протеинкиназы A ). Вторичные эффекторные белки, в свою очередь, запускают множественные третичные эффекторы и далее нисходящий эффекторный каскад вплоть до эффекторов N-го порядка. Поскольку открытие и закрытие ионных каналов при помощи метаботропных рецепторов требует активации целой системы вторичных посредников и эффекторов, то ионные каналы, активность которых косвенно регулируется этими рецепторами, требуют больше времени для своего открытия или закрытия, чем ионные каналы, ассоциированные с ионотропными рецепторами. Поэтому метаботропные рецепторы не вовлечены в механизмы, которые требуют от клетки очень быстрой реакции. Однако эффекты, опосредуемые метаботропными рецепторами, держатся дольше. Метаботропные рецепторы, как правило, сохраняют активированное состояние в течение секунд или минут после связывания с агонистом. Поэтому они имеют более длительные эффекты, чем ионотропные рецепторы, которые открываются почти мгновенно (время реакции ~10 мкс), но остаются открытыми лишь в течение нескольких миллисекунд. Кроме того, эффект ионотропных рецепторов заключается преимущественно в локальных изменениях электрического потенциала мембраны в окрестности рецептора, в то время как метаботропные рецепторы могут оказывать влияние на метаболизм и жизнедеятельность всей клетки.
Метаботропные рецепторы могут как открывать, так и закрывать ионные каналы клетки (в отличие от ионотропных рецепторов, активация которых всегда приводит к открытию в норме закрытого ионного канала). Они могут вызвать повышение возбудимости клеточной мембраны при помощи закрытия калиевых каналов клетки, что приводит к удержанию положительных зарядов внутри клетки и уменьшению тока, необходимого для возникновения потенциала действия . Метаботропные рецепторы на пресинаптической мембране могут ингибировать (чаще) или стимулировать (реже) выделение нейромедиаторов из пресинаптического нейрона. Метаботропные рецепторы могут быть субклассифицированы в зависимости от используемых ими механизмов передачи сигнала и используемых систем вторичных посредников и эффекторов на рецепторные тирозинкиназы, G-белок-связанные рецепторы и гуанилатциклазы.
Вторичные посредники метаботропных рецепторов
Примечания
- ↑ Hoehn K., Marieb E. N. // (неопр.) . — San Francisco: Pearson Benjamin Cummings, 2007. — ISBN 0-8053-5910-9 . 19 февраля 2015 года.
- ↑ Williams, S. J.; Purves, Dale. Neuroscience (неопр.) . — Sunderland, Mass: , 2001. — ISBN 0-87893-742-0 .
- ↑ Jessell T. M., Kandel E. R., Schwartz J. H. Principles of Neural Science (неопр.) . — New York: McGraw-Hill Education , 2000. — ISBN 0-8385-7701-6 .
- Schmitz D., Mellor J., Nicoll R. A. Presynaptic kainate receptor mediation of frequency facilitation at hippocampal mossy fiber synapses (англ.) // Science : journal. — 2001. — March ( vol. 291 , no. 5510 ). — P. 1972—1976 . — doi : . — .
Для дополнительного чтения
- Zimmerberg, B. 2002. Dopamine receptors: A representative family of metabotropic receptors. Multimedia Neuroscience Education Project
- 2020-08-04
- 1