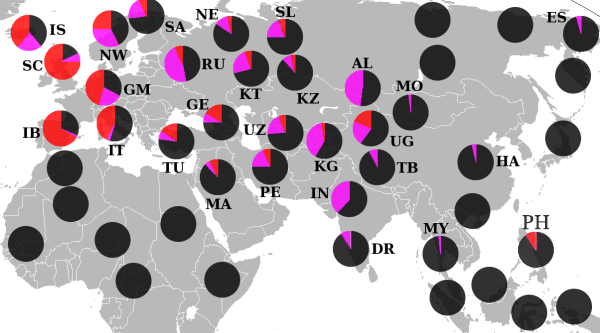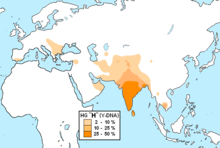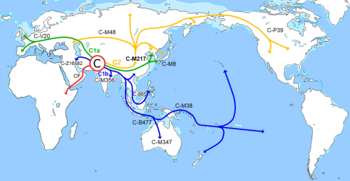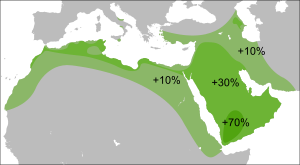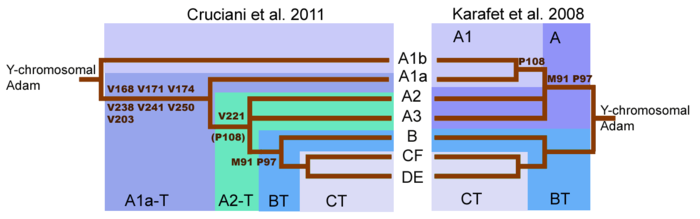
Гаплогруппа R (Y-ДНК)
- 1 year ago
- 0
- 0

|
|
Этот перевод статьи с другого языка
требует улучшения (см.
Рекомендации по переводу
)
.
|
Гаплогруппа R1a (М420) — Y-хромосомная гаплогруппа , распространённая в Восточной и Центральной Европе , Средней и Южной Азии и Южной Сибири .
R1a возникла около 22 тысяч лет назад (по другим данным — около 25 тыс. лет назад ), предположительно в Азии (исходя из распространения линий R2 и R*). R1a-М420 делится на ветви R1a1-M459 и R1a2-YP4141.
R1a2-YP4141 (18200-12300) делится на две ветви R1a2a-YP5018 (1230-5700) и R1a2b-YP4132 (12300-6300).
YP5018 5700 лет назад распалась на Y45596, потомки которой обитают на Ближнем Востоке: в Турции, Армении, Грузии, Ираке, Катаре, Бахрейне. Вторая ветвь FTA11514 попала в Европу, где 1250 лет назад начали ветвиться в Германии и Италии.
Потомки YP4132 последние 6300 лет обитают восточнее в Пакистане, Индии, Иране. Откуда ветвь YP4131 (4600-1450) попала на Британские острова.
Мутация SNP R1a-M420 была обнаружена после R1a1a-M17, что привело к реорганизации линии, в частности, к созданию новой парагруппы (обозначенной R-M420*) для относительно редких линий, которые не находятся в субкладе R1a1-M459/SRY10831.2 , ведущем к линии R1a1a-M512/M17. Распределение субкладов R1a1a1b1a-Z282 (включая R1a1a1b1a2-Z280) в Центральной и Восточной Европе и R1a1a1b2-Z93 в Азии предполагает, что R1a1a-M17 выделилась в пределах евразийских степей. Место происхождения и развития этих субкладов значимо для определения происхождения и путей расселения протоиндоевропейцев .
В качестве прародины R1a, относят регион Южной Сибири.
R1a1a1-M417 это корневая и первая ветвь, пошедшая от гаплогруппы R1a-M420. Она разделилась между 25 и 22 тысячами лет назад. Это период последнего ледникового максимума. В исследовании 126 современных популяций со всей Евразии от 2014 года, проведённом Питером А. Андерхиллом и соавторами с участием 16 244 человек, авторы пришли к предположению о том, что «начальные эпизоды диверсификации гаплогруппы R1a, вероятно, произошли в окрестностях современного Ирана ». Согласно расчётам Андерхилла и др. (2014), субклад R1a1a1-M417, диверсифицировался на линии, ведущие к субкладам R1a1a1b1a-Z282 и R1a1a1b2-Z93, около 5800 лет назад . Ранне энеолитический образец NEO113 из региона Среднего Дона ( Голубая Криница ) возрастом 7300 л. н. помещается в базальный субклад гаплогруппы R1a вместе с ранними образцами, связанными с комплексом шнуровой керамики, poz81 (R1a1a1a-CTS4385>CTS4385*, 2880-2630 до н. э., Obłaczkowo, Poland) и RISE446 (R1a1a1a-CTS4385>L664>S3479, 2829-2465 до н. э., Bergrheinfeld, Bavaria, Germany), что делает его самым ранним наблюдением этой базальной линии, о котором сообщалось до сих пор .
R1a показывает сильную корреляцию с индоевропейскими языками Южной и Западной Азии, Центральной и Восточной Европы . Наибольшее распространение она имеет в Восточной Европе, Западной и Южной Азии и Центральной Азии. В Европе преобладает R1a1a1b1a-Z282, в то время как в Азии доминирует R1a1a1b2-Z93. Связь между Y-хромосомной гаплогруппой R1a1a-M17 и распространением индоевропейских языков впервые была отмечена Татьяной Зерджаль и её коллегами в 1999 году .
Взрывообразный рост числа потомков основателя субклады R1a1a1b2-Z93 произошёл в Южной Азии примерно 4,5—4 тыс. л. н. Расширение маркеров R1a1a1b2-Z93 в промежуток между 4500 и 4000 лет назад на несколько столетий предшествует угасанию и краху Индской цивилизации .
Согласно исследованию древней ДНК В. Нарасимхана и др. (2018), степные скотоводы являются вероятным источником Y-хромосомной гаплогруппы R1a в Индии .
По данным на 2017—2018 год по исследованиям, проводимым Европейскими генетиками, известно следующее [ источник не указан 942 дня ] .
1 — Гаплогруппа R1a через гаплогруппу R1 происходит от старшей по возрасту гаплогруппы R . Гаплогруппа R произошла в Северной Азии перед Последним Ледниковым максимумом , в промежутке между 26 599 и 19 000 лет назад. Наиболее древний найденный образец данной гаплогруппы (R*) найден у мальчика со стоянки Мальта́ в Иркутской области . Этот мальчик жил в племени охотников на северных оленей . В силу постоянных миграций эти носители гаплогруппы R доходили до современной Европы и Южной Азии и таким образом способствовали её распространению. В современный период именно эти регионы наиболее богаты данной гаплогруппой и её производными — R1b в Западной Европе , R1a в Восточной Европе , Центральной и Южной Азии и R2 в Южной Азии [ источник не указан 942 дня ] .
2 — В результате серии мутаций корневой гаплогруппы R появилась Y-хромосомная гаплогруппа R1 и из неё — R1a . Срок возникновения — приблизительно после последнего ледникового максимума. Точное место возникновения именно гаплогруппы R1a на данном этапе изучения не известно. Разброс возможных географических регионов — Пакистан , Северо-Западная Индия , Балканы — данные регионы имеют наибольшее генетическое разнообразие этой гаплогруппы.
3 — Существует версия, что на Балканы гаплогруппа R1a попала от миграционных потоков, идущих из Евразийских степей, и в силу того, что миграции шли волнами, было обеспечено высокое разнообразие мутаций. Генетики предполагают, что гаплогруппа R1а могла возникнуть либо в Средней Азии , либо в Сибири [ источник не указан 942 дня ] .
При первом варианте R1a с носителями сначала попала в Восточную Европу. Второй вариант — предварительно люди мигрировали в период палеолита на юг, в Среднюю Азию, затем — в Иран , затем — на Кавказ во время неолита , после Кавказа через Каспийские степи уже попали в Восточную Европу . За данный вариант говорит то, что в Иране обнаруживаются очень старые субклады гаплогруппы R1a-М420, в регионе Загрос [ источник не указан 571 день ] . При третьем варианте развития событий люди из места происхождения гаплогруппы разделились на два потока — один в Казахстанские степи и затем в Восточную Европу, другой отклонился и попал в Иран.
4 — Ряд теорий, упиравших на то, что гаплогруппа R1a попала в Европу через Анатолию в период миграции неолитических земледельцев, натолкнулись на несостыковки. 99 % современной R1a происходит от гаплогруппы R1a1a1-M417 — а она, в свою очередь, имела расширение в период бронзового века , но не в период неолита. Филогенетика данной гаплогруппы выводит к месту её происхождения — Восточной Европе. Большая часть R1a на Ближнем Востоке — глубокая субклада, отделившаяся от гаплогруппы R1a1a1b2-Z93, имевшей место происхождения в России [ источник не указан 942 дня ] .
С другой стороны, к северу от этих ближневосточных регионов, уровни R1a1a начинают расти на Кавказе неравномерно. Несколько изученных популяций не показали никаких признаков R1a1a, в то время как самые высокие уровни, обнаруженные до сих пор в регионе, среди карачаевцев ок. 40 % и среди балкарцев ок. 35 % представлены доминирующей гаплогруппой R1a1a .
При исследовании образцов, максимально близких к неолитическим земледельцам, не обнаружено мутаций, связывающих их с R1a, даже относительно поздний земледелец Эци , живший 5 300 лет назад, не несёт в себе никаких признаков гаплогруппы R1a (у него G2a), так же R1a отсутствует у современных сардинцев , являющихся наиболее близкими потомками земледельцев, мигрировавших в период неолита. Данные факты указывают, что гаплогруппа могла произойти именно в Восточной Европе и затем в период бронзового века распространиться [ источник не указан 942 дня ] .
Т. Кивисилд в 2003 году предположил либо Южную, либо Западную Азию , а Мирабал в 2009 году указывал на Южную и Центральную Азии . Другие исследователи предполагают происхождение R1a на территории современной Украины , в Средней Азии и в Западной Азии .
В 2000 году предположено происхождение на территории Украины и постледниковое распространение R1a1 во время последнего ледникового максимума . Распространение генома по этой теории обеспечивалось в том числе благодаря экспансии курганной культуры в Европе и на Восток .
предлагает центрально-азиатское происхождение, предполагая, что распределение и возраст R1a1 указывают на древнюю миграцию, соответствующую распространению Культуры курганных погребений в их экспансии из Евразийской степи .
В 2012 году предположено, что R1a1a выделилась в евразийских степях или на Ближнем Востоке и на Кавказе. Внутренняя и Центральная Азия, является перекрывающейся зоной для линий R1a1-Z280 и R1a1-Z93, что подразумевает раннюю зону дифференциации R1a1a-M198 , предположительно произошла в пределах Евразийских степей или на Ближнем Востоке и на Кавказе, поскольку они лежат между Южной Азией и Центральной и Восточной Европой .
Три генетических исследования, проведённые к 2015 году, подтвердили курганную гипотезу Гимбутас . Согласно этим исследованиям, гаплогруппы R1b и R1a , теперь наиболее распространенные в Европе, были распространены из российских степей наряду с индоевропейскими языками. Исследования также обнаружили аутосомный компонент, присутствующий в современных европейцах, которого не было у неолитических европейцев, которые были бы введены с отцовскими линиями R1b и R1a, а также с индоевропейскими языками .
_and_R1a1a_oldest_expansion_and_highest_frequency_(2014).jpg)





Дэвид Энтони считал людей ямной культуры предками индоевропейцев . Согласно исследованию 2015 года, массовая миграция людей ямной культуры имела место около 2500 лет до нашей эры. Её связывают с тем, что данная культура одной из первых массово научилась использовать лошадей для передвижения людей и перевозки грузов, а также активно применять бронзовое оружие. Генетики из Стэнфордского университета и др. (Poznik et al. 2016) выявили, что с демографической экспансией в Западную Европу в бронзовом веке связана не гаплогруппа R1a, а гаплогруппа R1b1a1a2a1a-L151/L11 (L23 и её субклады), в то время как субклад R1b-Z2103 , представленный в ямных погребениях, в этой экспансии не участвовал .
Ямную культуру сменила катакомбная культура , у представителей которой также определили Y-хромосомную гаплогруппу R1b , а не R1a .
Первое сильное распространение гаплогруппы R1a коррелирует с миграциями именно культуры боевых топоров . С этой миграцией в Германию и Нидерланды попал субклад R1a1a1b1-Z283. В Скандинавию — R1a1a1b1a3-Z284. Люди этой культуры смешались с племенами, населявшими Германию до их прихода и носившими гаплогруппы I1 и I2, таким образом образовав первую индоевропейскую культуру на территориях нынешних Германии и Скандинавии. Однако эта культура не была первично германской, а была первичной индоевропейской общностью и скорее всего несла в себе основы и для германцев и для прото-балто-славян. Языковой анализ подтверждает такой подход — первичный германский язык развивался как смесь прото- балто-славянских языков культуры шнуровой керамики (R1a1a1b1-Z283) и более поздней унетицкой культуры , люди этой культуры были гибридными носителями R1a/ R1b [ источник не указан 942 дня ] .
Эта первая волна миграции большей частью состояла из носителей R1b , но в её массе были также племена, носившие R1a в виде субклады R1a1a1a1-L664. Эти племена смешались затем с носителями R1b, в особенности с её субкладой R1b1a1a2a1a1-U106, от которой спустя много веков произойдут в итоге германцы . Данная миграция поглотила и закончила в итоге культуру колоколовидных кубков и культуру шнуровой керамики в Центральной Европе . Из получившегося процесса вышла унетицкая культура и стала основой для германских, кельтских и итальянских культур [ источник не указан 942 дня ] .
Унетицкая культура расширила ареал для Скандинавии и таким образом дала начало скандинавскому Бронзовому веку . В этот же период люди, носившие R1a1a1a1-L664 и R1b (L11 и U106), достигли Скандинавии. Люди в тот период в Скандинавии говорили на первично германском языке, основанном на собственном лексиконе, лексике поглощённой Культуры шнуровой керамики, являвшейся носителем R1a1a1b1a3-Z284 и говорившей на смеси прото-балто-славянских языков и третий компонент это носители неиндоевропейских языков и гаплогрупп G2a, I1, I2. В итоге современная Скандинавия состоит из 40 % I1, 20 % R1a и 40 % R1b [ источник не указан 942 дня ] .
Миграция из ямной культуры через Северо-Восточную Европу и связанная с этим культура шнуровой керамики . Эти племена заселили наиболее плотно нынешние страны — Польшу , Белоруссию и Украину , Россию образовав ядро протобалто-славян . Высокая концентрация гаплогруппы R1a в балтийских и в славянских странах связана не только с миграцией этой культуры, а также объясняется несколькими последовавшими волнами переселений с территории нынешней России, последняя из которых имела место между V и X веками нашей эры .
В процессе движения культуры шнуровой керамики она поглотила в себя трипольскую культуру [ источник не указан 510 дней ] на Западной Украине и в северо-восточной Румынии , люди которой были носителями Y-хромосомных гаплогрупп G2a и I2a1b-M423. В них также содержались следы от генетики земледельцев с Ближнего Востока — гаплогруппы E-V13 , C1a, I2a2 и I2c. Культура шнуровой керамики в период своего развития соседствовала в степях между Днепром и Уралом со срубной культурой (R1a-Z93) и в Польше с тшинецкой культурой . Последняя славянская миграция, внëсшая заметный вклад, произошла начиная с VI века нашей эры с территорий Украины до Польши, Словакии и Словении . Эта миграция заполнила людьми образовавшиеся свободные пространства, которые получились после миграции и вторжения древних восточных германских племён в Римскую империю .
Балтийская ветвь народов развилась из нескольких культур входивших в культуру шнуровой керамики и боевых топоров, в том числе, по мнению нескольких историков, в балтских племенах заметны схожие с фатьяновской культурой 3200 — 2300 лет до н. э. культурные традиции, включая керамику [ источник не указан 510 дней ] . Фатьяновская культура (R1a1a1b2-Z93) является восточным вариантом культуры боевых топоров — шнуровой керамики , который появился на Северо-Востоке Европы. В раннем бронзовом веке кочевники, носители R1a, смешались с носителями уральских языков N1c1. Этот подход подтверждается наличием гаплогрупп R1a и N1c1 в Южной Финляндии , до Литвы и на Северо-Западе России. Латышские и литовские носители R1a так же несут в себе балто-славянские линии, такие как R1a1a1b1a1-M458, R1a1a1b1a2b-CTS1211 и R1a1a1b1a2a-Z92 [ источник не указан 942 дня ] .
Носители прото индоиранского языка , впоследствии ставшие ариями в « Ригведе » и « Авесте », возникли в синташтинской культуре [ источник не указан 1140 дней ] , 2100—1750 лет до н. э. в долинах рек Тобол и Ишим к востоку от Уральских гор . Синташтинская культура была наследницей более старой культуры — абашевской 2500—2900 лет до н. э., пришедшими с Верхнего Дона и Волги на Урал . Также в образовании синташтинской культуры принимала участие полтавкинская культура (2700—2100 лет до н. э.). Синташтинская культура и её носители R1a1a1b2-Z93 были первыми носителями индоевропейского языка к западу от Урала в период бронзового века. Эти племена в итоге вышли к обширным равнинам и пустыням Средней Азии и дошли до богатых металлами Алтайских гор . Арии быстро распространились по всей Центральной Азии , от берегов Каспийского моря до Южной Сибири и Тянь-Шаня . Этому способствовала активная торговля, сезонные миграции населения вслед за стадами и грабительские походы. Именно синташтинцам приписывают одни из самых ранних образцов боевых колесниц — 2100 лет до н. э. Благодаря колесницам они смогли распространиться на богатый металлами горнодобывающий регион Бактрию — современная граница Туркменистана , Узбекистана , Таджикистана и Афганистана . В итоге они могли добывать большое количество меди на Урале и протоиндоиранцы из синташтинской культуры активно экспортировали её на Ближний Восток [ источник не указан 942 дня ] .
Индоиранцы освоили Зеравшанскую долину , основав в итоге Петровскую медно-рудную колонию в Тугае в 1900 году до н. э. из-за большого содержания в ней медной руды. Олово добывалось в Карнабе и Мушистоне. Олово во времена позднего бронзового века было ценным ресурсом — медно-оловянный сплав прочнее на излом и удар, чем обычная бронза с мышьяком. К 1700 годам до н. э. индоиранцы расширили ареал до нижней долины Амударьи и основали общины, занимавшиеся земледелием с орошением — тазабагьябская культура . К 1600 годам до н. э. старые города Бактрии были заброшены в силу новых волн степных кочевников с севера, которые наступали на поселения. Индоиранской культурой в Центральной Азии являлась андроновская культура , существовавшая до 800 г. до н. э. Индоиранцы продвигались дальше на юг и достигли Гиндукуша [ источник не указан 942 дня ] .
К 1700 г. до н. э. кочевники- скотоводы проникли в Белуджистан — Юго-Западный Пакистан . В долину реки Инд они пришли к 1500 г. до н. э. Северная и центральная части Индийского субконтинета были заселены ими к 500 г. до н. э. Взаимопроникновение культуры степных скотоводов, имевших на вооружении колесницы и носителей индской культуры и языка, привело к взаимному обмену знаниями и языками. В итоге к 1600 г. до н. э. была захвачена Ассирия и инды стали правителями Митанни к 1500 г. до н. э. Мидийцы , парфяне и персы — все носители иранских языков из андроновской культуры — перешли жить на Иранское плато к 800 г. до н. э. Оставшиеся в Центральной Азии племена стали в итоге скифами , таковыми их и зафиксировала историческая наука, зародившаяся в Древней Греции [ источник не указан 942 дня ] .
Оставшиеся после миграций представители ямной культуры , которые обитали в Понтийско — Каспийских степях, стали сарматами и так же были зафиксированы в исторической науке Древней Грецией и Древним Римом. Эти перемещения и миграции населения привели к распространению гаплогруппы R1a в Южной части Центральной Азии и в Индийском субконтиненте. Наиболее высокая концентрация R1a — до 65 % — наблюдается на стыке границ Киргизии , Таджикистана и Северного Афганистана . В Индии и Пакистане [ источник не указан 942 дня ] .
R1a составляет от 15 до 50 % генетического материала у населения в зависимости от региона, этнической группы и кастового сословия. R1a наиболее часто представлена на Северо-Западе Индийского субконтинента, наименее часто — На юге — Тамил , Керала , Карнатака , Андхра-Праде , Бенгалия . Более 70 % браминов — высшей касты в Индии — несут в себе значительную часть R1a1. Однако линии митохондриальной ДНК в Южной Азии в подавляющем большинстве случаев являются доиндоевропейскими. Например, в Индии более 75 % мтДНК — митохондриальные гаплогруппы М и R — это местные линии и только 10 % — восточно-азиатские линии. Оставшиеся 15 % гаплогрупп — это 7 — 8 % ближневосточных линий и только 7 % могут быть отнесены к линиям, попавшим с носителями из Понтийско-Каспийских степей , в основном в форме гаплогрупп U2 и W . Европейские линии мтДНК гораздо чаще представлены в Центральной Азии, в Афганистане и на Севере Пакистана. Первые крупные поселения, в которых постоянно проживали женщины индоарийской языковой группы и генетики, это Северный Пакистан, Западная Индия — Пенджаб и Гуджарата и Северная Индия — Уттар — Прадеш , где митохондриальные гаплогруппы U2 и W являются наиболее распространенными сегодня [ источник не указан 942 дня ] .
В 1934 году шведский археолог на северо-западе Китая , в районе реки Тарим (ныне Синьцзян-Уйгурский автономный район ), обнаружил около 200 мумий европеоидного типа . Наиболее старая из мумий датируется примерно 2 тыс. лет до н. э. При анализе ДНК у 11 из 12 мумий мужчин была найдена гаплогруппа R1a1 по нестабильному снипу M17. Современные жители Таримского бассейна — уйгуры . Они несут в себе до 20 % R1b1a1a1-M73 и 30 % R1a1. У таримский мумий определили митохондриальные гапогруппы U4, U4’9, U5b2c, U5a2, T1a1b, I1b, G3b, H15b1, D4j1b и A17.
Ранее было выдвинуто две теории о происхождении этой культуры. Первая — это группа ранних скотоводов из репинской культуры 3700 — 3300 лет до нашей эры, которые мигрировали из Донских и Поволжских степей до гор Алтая, затем основали афанасьевскую культуру 3600—2400 лет до нашей эры и уже от неё распространились до Таримского бассейна. Вторая — таримские мумии это люди, происходившие из протоиндо-иранского народа, которые мигрировали по всей Центральной Азии примерно 2000 лет до нашей эры из синташтинской культуры .
Исследование 2021 года показывает, что популяция, связанная с мумиями, содержит не индоевропейские субклады R1a, а базальные субклады этой гаплогруппы, восходящие к , и она была значительно изолирована от соседей начиная с 9 тыс. л. н., а к 4,5 тыс. л. н. расселилась в бассейне Тарима. Также у них определяли Y-хромосомную гаплогруппу R1b1c-PH155/PH4796 [ источник не указан 124 дня ] .
При исследовании ДНК установлено, что гунны — это народ, появившийся при смешивании европейских носителей и северо-востока Азии. При этом большая часть генетики в гуннах была от Хунну — древнего кочевого народа, жившего в Китае. Современные жители территорий, где раньше жили Хунну, несут 90 % монгольских родов против 10 % европейских. Самая старая находка мтДНК европейского рода найдена на границах Монголии и рядом с озером Байкал , датируется 4000 лет до нашей эры. Данные выводы свидетельствуют, что носители тюркского языка быстро заменили языки скифов и иранские диалекты по всей Центральной Азии.
Таким образом, многие народы переняли тюркский язык , но оставались почти не затронуты тюрками в плане генетики. В итоге болгарский язык , хоть и назван в честь тюркского племени — булгар , не является тюркским. Это славянский язык с незначительным влиянием тюркского. Венгерский язык , иногда ошибочно принимаемый за язык, оставшийся от гуннов-хуннов, на самом деле является языком уральской семьи — мадьяр . Тюркские языки, несмотря на огромную площадь распространения и большое количество разных народов-носителей, лингвистически не сильно различаются между собой и носители разных тюркских языков понимают друг друга довольно хорошо. Это связано с относительно недавним распространением языка и кочевым характером его носителей [ источник не указан 942 дня ] .
При этом генетическое совпадение между представителями ямной культуры и культуры боевых топоров достигает 75 % [ источник не указан 995 дней ] . Исследователи отмечают, что R1a и R1b могли распространиться в Европу с Востока после 3000 лет до нашей эры . Однако при исследованиях все семь образцов из ямной культуры принадлежали к R1b1a1a2-M269, а R1a1a не была обнаружена в образцах, отнесённых к ямной культуре . Это поставило вопрос — откуда возникла R1a1a в культуре боевых топоров, учитывая что ямная культура не являлась носителем данной субклады.
Семёнов и Булат спорят о происхождении R1a1a в культуре боевых топоров, отмечая, что несколько исследований указывают на присутствие R1a1a в генетике данной культуры .
Исследования от 2015 года обнаружили, что часть предков людей ямной культуры ведёт свою генетику с Ближнего Востока, и что неолитический период в ямной культуре, связанный с освоением соответствующих навыков, начался на Балканах. При этом исследователь Хаак, поддерживающий эту версию, отмечает, что генетически люди ямной культуры не происходят от земледельцев Ближнего Востока, а ведут корни от людей с Ближнего Востока .
Рёссен — культура среднего неолита, существовавшая между 4600—4300 годами до н. э., была распространена в Германии и предшествовала культуре боевых топоров , но не напрямую, а через последовательно сменяющиеся культуры — культуру воронковидных кубков и культуру шаровидных амфор . Рёссен может иметь связь с древним подклассом R1a, а именно R1a1a1a1-L664 — согласно ДНК-исследованиям, она сформировалась к 2700 году до нашей эры.
Некоторые исследователи полагают, что R1a попала на Балканы через Анатолию , а уже оттуда распространялась дальше на северо-запад, до культуры рёссен, затем на восток от трипольской культуры до ямной культуры и афанасьевской культуры . При этом исследователи признают — R1a отсутствует в промежуточных культурах между Ближним Востоком, Анатолией и Балканами. Аско Парпола, исследуя эту проблему, предположил, что трипольская культура соотносится с поздним вариантом праиндоевропейского этноса. Он отмечает, что в трипольской культуре, возможно, изобретено колесо, а также что, вероятно, люди трипольской культуры была захвачены или ассимилированы носителями праиндоевропейского языка в 4-м тысячелетии до н. э., после чего культура распространилась до Понтийской степи к 3400 годам до нашей эры и в результате развития перетекла в ямную культуре .
На данный момент мало информации о прибытии прото греческих носителей языка из степей. Микенская культура возникла около 1650 лет до нашей эры и явно связана с прибытием переселенцев от степных культур. Микенский язык тесно связан с прото индо-иранскими языками и разделение их на отдельные языки произошло относительно поздно, между 2500—2000 годами до нашей эры. Своеобразные микенские колесницы , наконечники копий, кинжалы из бронзы и другие изделия из неё показывают большое сходство с необычным археологическим памятником, называемым в научной литературе Сейминско-турбинский феномен (с 1900 по 1600 лет до нашей эры) — в нём известна Володарская стоянка и в том числе в ней — Сейминский могильник [ источник не указан 942 дня ] .
Изделия и предметы из этого археологического памятника, расположенного рядом с городом Володарск (Нижегородская область), напрямую коррелируют с находками в ареале Микенской цивилизации — это южная часть Балканского полуострова . Также схожие признаки культуры, оставившей Сейминский могильник, найдены в Монголии , что говорит о высокой мобильности кочевых степных народов. Данные факты указывают на возможную миграцию культур из лесостепной зоны центральной части Восточно-Европейской равнины на Балканы , смешивание их с местным населением и затем основание ими Микенской цивилизации, положившей начало Греческой культурной общности [ источник не указан 942 дня ] .

Часть генетической составляющей в жителях Южной Азии происходит от западно-европейского населения, и некоторые исследователи предполагали, что субклад R1a1a1b2-Z93, возможно, прибыл в Индию через Иран и расширился там во время Хараппской цивилизации .
В 2015 году предположено , что корни R1a1a1b2-Z93 лежат в Западной Азии и её распространение вместе с L342.2 проходило в Юго-Восточном направлении от Закавказья до Южной Азии" , мотивируя это археологическими данными о восточной популяции Западной Азии в 4-м тысячелетии до нашей эры, кульминацией которого стали так называемые миграции куро-араксской культуры в период после (период истории Месопотамии между 4000 — 3100 лет до нашей эры) . Тем не менее Лазаридис отметил, что в куро-аракский образец I1635 была привнесена Y-хромосомная гаплогруппа R1b1-M415 (xM269) также называемая R1b1a1b-CTS3187 .
Согласно результатам исследования 2014—2015 годов , диверсификация-отделение R1a1a1b2-Z93 и процессы урбанизации в долине Инда произошли 5600 лет назад, и географическое распределение R1a1a1b2a1~-M780 (нижестоящего к R1a1a1b2a-Z94) может отражать это. Согласно ДНК, прослеживаемого через генеалогическое древо, R1a1a1b2a1~-M780 сформировалась 4700 лет назад. Этот процесс по возрасту совпадает с движением на восток между 2800 и 2600 годами до н. э. ямной культуры в область полтавкинской культуры , предшественницы синташтинской культуры , из которой возникли индоиранцы . Субклад R1a1a1b2a1~-M780 сконцентрирован в долине Ганг, центре классической ведийской цивилизации [ источник не указан 715 дней ] .
В 2016 году отмечено , что «произошло поразительное расширение» субклады R1a1a1b2-Z93 в промежуток между 4500—4000 л. н., что на несколько столетий предшествует угасанию, краху Индской цивилизации .

В 2003 году высказано предположение о происхождении либо в Южной, либо в Западной Азии . Это связано с фактом: гаплогруппа R1a, ранее уже увязываемая как маркер с предполагаемым индоарийским вторжением, была найдена в самой высокой частоте генома в Пенджабе и также в высокой частоте (26 %) — в племени , одно из коренных индийских племён группы — адиваси . Это открытие вместе с более высоким составляющим маркера R1a — тандемное повторное разнообразие в Индии и Иране, по сравнению с Европой и Центральной Азией, предполагает, что источником этой гаплогруппы могут быть Южная и Западная Азии.
В исследовании 2009 года указывается в качестве возможной «прародины» гаплогруппы как Южная, так и Центральная Азия .
Южноазиатские популяции имеют наивысшее разброс микросателлитов в пределах R1a1a , и последующих, более поздних датировок на ближайшего общего предка . А R1a1a присутствует как среди высших каст (брахманов), так и низших каст, хотя присутствие значительно выше среди брахманов . Исходя из этих данных, некоторые исследователи пришли к выводу, что R1a1a возникла в Южной Азии , исключая существенный генетический приток от индоевропейских мигрантов .
Однако это разнообразие и последующие более ранние датировки на выявление ближайшего общего предка также могут быть объяснены исторически большим числом населения, что увеличивает вероятность диверсификации и изменения микросателлитов . R1a1 и R2, возможно, распространялись в Южную Индию из юго-западного региона Азии несколько раз .
В 2017 году отмечено , что R1a в Южной Азии наиболее вероятно распространилась от одного центрального пучка источников в Центральной Азии, по крайней мере — трёх. И, вероятно, эти генетические линии в дальнейшем развивались в границах Индийского субконтинента , что согласуется с несколькими волнами миграций .

Генеалогическое древо гаплогруппы R1a имеет три основных уровня ветвления с наибольшим количеством определённых субкладов в доминирующей и наиболее известной — R1a1a, она же R1a1.
Топология R1a выглядит следующим образом .
Согласно работе Tatiana Karafet et al. (2014), быстрый процесс диверсификации гаплогруппы K-M526, вероятно, произошел в Юго-Восточной Азии с последующими расширениями на западе предков гаплогрупп R и Q .
Самаые древние известные R1a обнаружены у следующих двух (возраст около 10800 лет ):
(субклад R1a1b~-YP1272>YP1301) определена у представителя культуры Веретье из могильника Песчаница 1 PES001 (10785—10626 лет назад) на озере Лача (Каргопольский район Архангельской области) .
У мезолитчика I1819, жившего 10825—10561 лет назад Василевка-на-Днепре , определена .
R1a1- SRY10831.2 определили у мезолитического образца UZOO-74 с Южного Оленьего острова на Онежском озере, жившего ок. 8 тыс. лет назад .
R1a1-M17 была определена у представителя Глазковского некрополя эпохи неолита (8000—7000 лет назад) .
Гаплогруппа R1a обнаружена в останках следующих культур и народов: культура боевых топоров , Культура полей погребальных урн , синташтинская культура , андроновская культура , пазырыкская культура , тагарская культура , таштыкская культура , у жителей древнего Танаиса (город) , в таримских мумиях , у аристократов Хунну , в двух древних останках хазар .
Субклад R1a1a1b1-Z283 обнаружен у представителя культуры боевых топоров , в захоронении (Bergsgraven), в Швеции , номер теста ber1 .
У темнокожих представителей фатьяновской культуры определена Y-хромосомная гаплогруппа R1a1a1-M417, при этом у половины образцов удалось уточнить субклад R1a1a1b2-Z93 .
Линия R1a1a1b2a1~-Y3, предковая для субклада R1a1a1b2a1a-L657, в настоящее время распространëнного особенно в Индии в высшей варне — у браминов , найдена у образца I6561 в Александрии у реки Оскол , в Харьковской области (Alexandria, Ukraine_MBA, Украина). Датировка захоронения 2134—1950 гг. до н. э.
| Номер теста | Захоронение | Датировка | Субклады |
|---|---|---|---|
| kzb005 | Казбурун 1 | 1880—1690 гг. до н. э. | R1a-M459, R1a- Z283, Z280 |
| kzb008 | Казбурун 1 | 1880—1690 гг. до н. э. | R1a-M459, R1a- Z283, Z280, S24902 ? |
| kzb002 | Казбурун 1 | 1875—1665 гг. до н. э. | R1a-M459, R1a- Z93 , YP5585 |
| kzb003 | Казбурун 1 | 1765—1630 гг. до н. э. | R1a-M459, R1a- Z93, Z94 |
| mur002 | Мурадым 8 | — | R1a-M459 |
| mur003 | Мурадым 8 | 1880—1685 гг. до н. э. | R1a-M459, R1a- Z93 |
| kzb007 | Казбурун 1 | 1755—1630 гг. до н. э. | R1a-M459, R1a- Z93 , Z94, Z2124, Z2125, Z2123, Y934, BY30762 ? |
| Номер теста | Захоронение | Датировка | Субклады |
|---|---|---|---|
| cim357 | Глиное Сад | 914—805 гг. до н. э. | R1a-M459, R1a- Z93 |
| Номер теста | Захоронение | Субклады |
|---|---|---|
| scy193* | Глиное | R1a-M459, R1a- Z93 , Z94, Z2124, Z2125, Z2123, Y934, BY30762, BY30764 ? |
| Номер теста | Захоронение | Датировка | Субклады |
|---|---|---|---|
| chy002 | Чёрный Яр | 65 — 220 гг. н. э. | R1a-M459, R1a- Z93 , Z94, Z2124, R1a1a1b2a2-Z2122, Y57, Y52 |
| tem002 | Темясово | 125—240 гг. н. э. | R1a-M459, R1a- Z93 , Z94, Z2124, Z2125, S23592 , YP1558, Y73758, YP5844, FGC48758 |
| tem003 | Темясово | 130—320 гг. н. э. | R1a-M459, R1a- Z93 , Z94, Z2124, Z2125, Z2123 , YP3920 |
Скелеты отца и двух его сыновей с археологического объекта, обнаруженного в 2005 году недалеко от Ойлау ( Саксония-Анхальт , Германия) и датируемых примерно 2600 годом до н. э., показали положительный результат для маркера Y-SNP SRY10831.2. Таким образом предковая клада присутствовала в Европе по меньшей мере 4600 лет назад и была связана с одним из археологических памятников культуры боевых топоров (шнуровой керамики) . Номер Ysearch для останков Eulau — 2C46S. У и у образцов из Волосово-Даниловского могильника (R1a-S224, фатьяновская культура, Ярославская область) значения аллелей по 10 локусам совпадают, а по 4 локусам различаются на единицу .
Также гаплогруппа R1a была выявлена у представителей межовской , хвалынской , срубной , потаповской , полтавкинской , тшинецкой . археологических культур, на стоянке Афонтова гора (бронза) , у скифов , у населения срубной культурно-исторической общности , среди аланских захоронений (субклад Z2124 — R1a1a1b2a2) , у населения салтово-маяцкой культуры (субклад Z2124 — R1a1a1b2a2) , у одного представителя культуры шнуровой керамики (боевых топоров) из Эсперштедта и у двух из саксонской пещеры Лихтенштейн , у одного представителя культуры полей погребальных урн из Хальберштадта и у двух из Эйлау . Гаплогруппа R1a1 прослежена у обитателей верховий Западной Двины с 5120±120 л. н. ( ) до VIII—V веков до н. э. (городище Анашкино) .
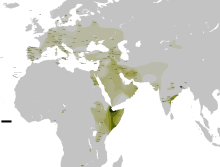
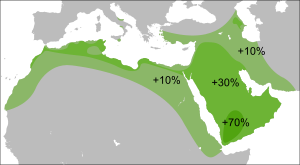
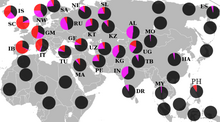
Распространение в странах Ближнего Востока и Передней Азии [ источник не указан 1027 дней ] :
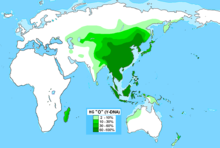
В Центральной и Южной Азии преобладает R1a Z93, являющаяся маркером древних индо-иранцев.
У киргизов R1а1а — 63,5 % . У алтайцев доходит до 58%, шорцев — 56,9 %, телеутов — 53,3% , В Афганистане R1a1a встречается у 51 % пуштунов и 64 % у таджиков , реже — среди хазарейцев (7 %). У хакасов — 28%, У казахов родов — ошакты 26,7 суан — 31,7 % и кожа 35 %. У узбеков —60-65% . и у уйгур —60 %. У тувинцев — 12% (наибольшая концентрация у северо-восточных — 26%), у бурят — 4%.
В Южной Азии R1a1a часто наблюдается с высокой частотой в ряде демографических групп.
В Индии высокие частоты этой гаплогруппы наблюдаются у западных (72 %) на восток, (60 %) на западе, (67 %) на севере и (31 %) на юге. Она также была найдена у нескольких южно-индийских дравидийских племён адиваси , включая (26 %), валмики из Андхра-Прадеша и в Тамилнаде , что говорит о том, что R1a1a широко распространена у племенных южных индийцев.
Кроме того, исследования показывают высокие проценты в региональных группах, таких как Манипури (50 %), на крайнем северо-востока и среди пенджабцев (47 %) на крайнем северо-западе
В Пакистане он встречается у 71 % среди племени Моханны в провинции Синд на юге и 46 % среди балтийцев Гилгит-Балтистан на севере. Среди сингальцев Шри-Ланки 23 % были обнаружены как R1a1a (R-SRY1532). Индусы района Читван в регионе Тераи Непал показывают его на уровне 69 % .
Частота R1a1a сравнительно низкая среди некоторых тюркоязычных групп, включая турок, азербайджанцев, казахов и якутов.
Но выше (от 19 до 28 %) в некоторых тюркских или монгольских группах северо-западного Китая — баоань , дунсян , саларов , уйгуров .
В Восточной Сибири R1a1a встречается среди некоторых коренных этнических групп, включая камчатцев и чукчей , и достигает у ительменов 22 % .
R1a1a был обнаружен в различных формах, в большинстве районов Западной Азии, в самых разных концентрациях. От почти не присутствующих в таких районах, как Иордания, до гораздо более высоких уровней в некоторых частях Кувейта, Турции и Ирана. Племена бедуинов Шаммар в Кувейте показывают самую высокую частоту на Ближнем Востоке — до 43 % .
В 2001 году отмечено , что в западной части страны у иранцев низкие уровни R1a1a, в то время как мужчины в восточной части Ирана имеют до 35 % R1a1a. В 2004 году R1a1a найдена примерно у 20 % иранских мужчин из городов Тегеран и Исфахан . В исследовании Ирана 2006 года отмечены гораздо более высокие частоты на юге, чем на севере.
Новое исследование показало 20,3 % R-M17 * среди курдских образцов, которые были взяты в провинции Курдистан в западном Иране, 9,7 % среди Мазандараниса в Северном Иране в провинции Мазандаран, 9,4 % среди гилаков в провинции Гилан, 12,8 % Персидский и 17,6 % среди зороастрийцев в Йезде, 18,2 % среди персов в Исфахане, 20,3 % среди персов в Хорасане, 16,7 % афро-иранцев, 18,4 % Кешми «Гешми», 21,4 % среди персидских говорящих людей Бандари в Хормозгане и 25 % среди Жители Балоха в Систане и провинции Балухстан.
Гаплогруппа R1a1a была найдена в повышенных уровнях среди выборки израильского населения, которые сами себя обозначили как левиты и ашкеназские евреи (левиты составляют около 4 % евреев). В исследовании Бехара установлено, что R1a1a является доминирующей гаплогруппой у ашкеназских левитов (52 %), хотя она редко встречается у ашкеназов-коэнов (1,3 %).
| Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||||||||||||||||||||||||||
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||||
| A0-T | |||||||||||||||||||||||||||||||||
| A00 | A0 | A1 | |||||||||||||||||||||||||||||||
| A1a | |||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||
| F1 F2 F3 | |||||||||||||||||||||||||||||||||
| G | |||||||||||||||||||||||||||||||||
| H | IJK | ||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||
| I | J | K2 | |||||||||||||||||||||||||||||||
| L (K1a) | T (K1b) | K2a/K2a1/ NO /NO1 | |||||||||||||||||||||||||||||||
| N | O | P (K2b2) /P1 | |||||||||||||||||||||||||||||||
| S (K2b1a) | M (K2b1b) | Q | R | ||||||||||||||||||||||||||||||
| Ключевые понятия |

|
|
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
|
Формирование славянских народов — генофонд, история
|
|||||||
|---|---|---|---|---|---|---|---|
| Основное | |||||||
| Генофонд славян |
|
||||||
|
Древнеславянские
археологические культуры |
|
||||||
| Гаплогруппа R1a | |
|---|---|
| Тип | Y-ДНК |
| Место появления | Южная Сибирь |
| Предковая группа | R1 |
| Сестринские группы | R1b |
| Субклады | Гаплогруппа R1a1 (Z282 (Европа) и Z93 (Азия)), R1a2 |
| Мутации-маркеры |
R1a — L62, L63, L120, M420, M449, M511, M513 R1a1a — M17, M198, M512, M514, M515, L168, L449, L457, L566 |