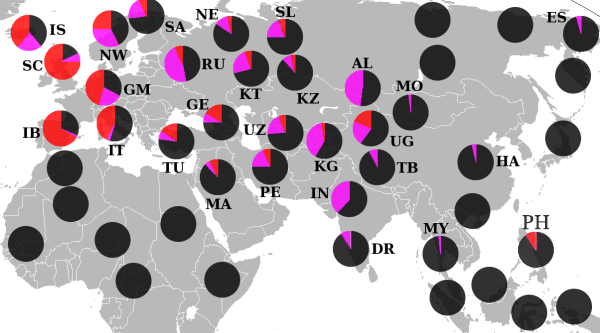
Гаплогруппа R (Y-ДНК)
- 1 year ago
- 0
- 0

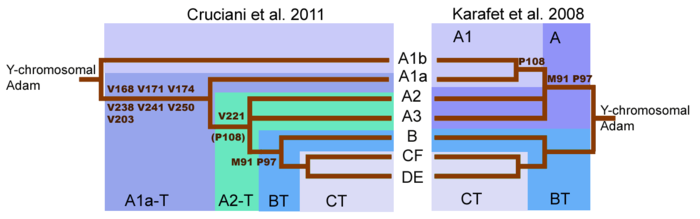
Гаплогруппа L (M20) — Y-хромосомная гаплогруппа . Определяется SNP (снипами) M11, M20, M61 и M185. Является вторичным потомком гаплогруппы K и первичной ветвью гаплогруппы LT(K1) , гаплогруппа L в настоящее время имеет альтернативное филогенетическое имя K1a и является параллельной и наиболее родственной Y-хромосомной гаплогруппе T (K1b) .
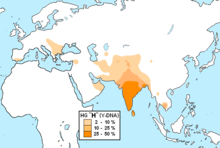
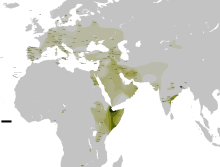
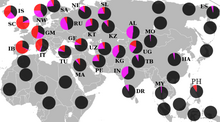
Мировая концентрация гаплогруппы приходится на Южную Азию , но составляет лишь 11,2 % от всего населения региона (281 из 2504 исследованных людей). В южной Азии отсутствует максимальное разнообразие субкладов (ветвей) L, оно приходится на Западную Азию и Кавказ , достигая пика в полосе, которая охватывает западный край Ирана , северный Ирак и северную Сирию .
Частота L не превышает 10-20 % практически ни у одного народа мира с населением более 1 млн человек, что делает это мужское потомство одним из самых редких на земле . Исключения: 85,2 миллионный народ джат из северо-западной Индии (36.8 %) и двухмиллионная языковая община с юга Индии (26.1 %) . В последнем случае данные могут быть искажены, так как исследовалось всего 46 человек.
На относительную редкость L указывают мужские генетические древа компаний Yfull и FTDNA (лидер по количеству Y-ДНК тестов). Количество выявленного разнообразия ветвей L составляет 339 вариантов у FTDNA и 349 вариантов у Yfull, это около 1 % от количества ветвей, выявленных у всех остальных гаплогрупп (данные на 30 июня 2021 ).
Одна из самых «горных». Вне южной Индии и южного Пакистана пики концентраций L приходятся на популяции совершенно разных горных регионов планеты:
Часть этносов, проживающих в данных регионах, сближаются гипотетической дене/сино-кавказской языковой семьёй , предложенной A. Старостиным в 1980-х годах, ещё до начала генетических исследований человечества.
Наибольшая частота L-M20 приходится на Южную Азию, охватывая соседние Памирские горы Центральной Азии. Пики концентраций L обнаруживаются у популяций Пакистана, Индии и Таджикистана, проживающих в высокогорных долинах в близости от Гималаев и Тибета:
Гаплогруппа L и три её субклада были обнаружены в 2006 году учёным S. Sengupta
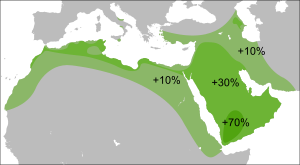
В 2014 году был обнаружен редкий субклад — L2 (L-L595) который встречается только в Европе и западной Азии и отстоит от субкладов L1 генетической дистанцией в 23000 лет до общего предка . Это наводит на мысли о Ближневосточной зоне зарождения субклада L.
В 2020 году у этнического курда из северной Сирии был выявлен редкий промежуточный субклад L1a3 (L-M2533*), входящий в ветвь L1a (L-M2481), но не имеющий родство с ветвями L1a1 (L-M27) и L1a2 (L-M357, L-L1307) ближе, чем на 17000 лет до общего предка.
За основу взяты древа ISOGG (2018) и Yfull (v7.06.00). Приведена распространенная в научных работах номенклатура геномутаций.
L M20 , M11, M61, M185 (гаплогруппа зародилась 42600 л. назад и распалась 23000 л. наз. на два субклада L1 и L2)
• L1 M22 (зародился 24000 лет наз. и распался 18200 лет наз. на два субклада — L1a и L1b)
• • L1a M2481 (зародился 18200 лет наз. и распался 17000 лет наз. на L1a1, L1a2 и L1a3)
• • • L1a1 (L1, L1a до 2015) M27, M76
Юг Индии в абсолютном большинстве связан с дравидоязычными народами. Исключение — 82.5 миллионый народ джат северной Индии, 36,8 % мужчин этого народа являются носителями L, однако точный процент L1a1 неизвестен.
• • • L1a2 (L3, L1c до 2015) M357, L1307 (Возраст до общего предка -
Пакистан: пики на юге и в высокогорных долинах на крайнем севере.
Памир Таджикистана.
Индия: Очень высокая концентрация 62,7 % на крайнем севере в Ладакхе , у этнической группы брокпа\минаро, которая составляет около половины от народа шина. В целом L1a2 составляет около 1 % на всю страну, рассеян по всей южной Индии крайне низкими частотами.
Чечня (Северный Кавказ, Россия). Уже более 1000 человек числятся в таблице чеченского национального ДНК проекта , созданного на базе американской лаборатории FTDNA. Его данные показывают, что от 7 до 14 % чеченских мужчин являются носителями L1a2, однако никто это число официально не уточнял. ДНК-исследования кавказцев, проведённые О. Балановским в 1998—2009, годах происходили в Чечне во времена боевых действий и не охватили все регионы республики.
• • • L1a3 (M2533*)
Северная Сирия, Турция. Крайне редок, на 2021 год известно лишь два его носителя. Выявлен у курда из северной Сирии в 2020-ом году.
• • L1b (L2 до 2015) M317, L655
Северная Турция и восточная часть Альп.
• L2-L595 обнаружен в 2014-ом году
Европа и западная Азия. Крайне редок. Не имеет родства с другими ветвями ближе 23000 лет до общего предка.
L-M2645*/M20/PF5570* сформировалась около 42,6 тыс. лет назад, последний общий предок современных носителей гаплогруппы L жил 24,2 тыс. лет назад . L-M20 происходит от Y-хромосомной гаплогруппы LT , которая происходит от гаплогруппы K-M9 . По словам доктора гаплогруппа L зародилась в сложной системе Памирских гор на территории нынешнего Таджикистана, и мигрировала на территорию Пакистана и Индии 30 000 лет назад . Тем не менее, большинство других иcследований указывают на западноазиатское происхождение гаплогруппы L и связывают её экспансию в долину Инда с неолитическими фермерами . На сей день абсолютное большинство исследованных останков неолитических фермеров показывают субклад гаплогруппы G (в прошлом G2a3), только очень малая часть относится к потомству гаплогруппы J2 . Палео-ДНК центральной и южной Азии эпохи неолита всё ещё не исследована, однако научная работа Singh 2016 , посвящённая изучению современного распространения J2 в южной Азии, отмечает, что география её распространения в мире сходится c расположением агрокультурных центров .
Учёные McElreavy и Quintana-Murci, описывая Индскую (Хараппскую) цивилизацию , утверждают, что «[т]олько Y-хромосомная гаплогруппа L-M20 имеет высокую среднюю частоту — 14 % в Пакистане и поэтому отличается от всех других гаплогрупп по своему частотному распределению. L-M20 также найдена, хотя и более низкими частотами, в соседних странах, таких как Индия, Таджикистан, Узбекистан и Россия. Как распределение частот, так и предполагаемое время экспансии (~ 7000 лет назад) этой генетической линии позволяют предположить, что её распространение в долине Инда может быть связано с экспансией местных фермерских групп в период неолита».
Обширное исследование Sengupta 2006 , в котором была изучена Y-ДНК 728 индийцев и 176 пакистанцев, выявило три субклада гаплогруппы L: L1-M76,M27 (ныне L1a1), L2-M317 (ныне L1b) и L3-M357 (ныне L1a2, L-1307 по yfull.com) . Почти все индийские носители гаплогруппы L показали субклад L1a1, причем L1a2, отстоящий от L1a1 генетической дистанцией в 17000 лет , оказался очень редким (0,4 % 3/728). В Пакистане же субклад L1a2 составил 86 % от всей L и достиг средней частоты 6,8 % в целом. L1a1 выявлена с частотой 7,5 % в Индии и 5,1 % в Пакистане, пик её разнообразия приходится на Махараштру — прибрежную западную часть Индии. Субклад L1b был найден только в Пакистане — 1,14 % (2 из 176).
Имеющиеся результаты исследований Y-ДНК мировых популяций, множество из которых приведены в данной статье, показывают редкость распространения субклада L1b-M317 в южной и центральной Азии , в то же время данный субклад имеет большинство носителей L-M20 в западной Азии и Европе . У субкладов L1a1 и L1a2 всё предстоит наоборот, их частота резко падает вне территорий южной Азии и исторической Бактрии . Из ненаучных данных это подтверждает и ДНК-проект «Haplogroup L» на базе коммерческой лаборатории FTDNA (около 700000 Y-хромосомных ДНК-тестов ), состоящий из 795 носителей L-M20 со всего мира. (7.06.2019)
Высокая частота субклада L1a2-M357 в Европе и на Кавказе обнаруживается только у нахоязычных народов Кавказа, в большей части у чеченцев — более 10 % (110 из 922 человек на 26.06.2019 г.) в проекте «Chechen-Noahcho DNA project», созданном на базе американской лаборатории FTDNA. Исследование Balanovsky 2011 , в котором участвовало 1525 мужчин из 14 популяций Кавказа, выявило L1a2 только у чеченцев и ингушей.
Обширное исследование (Mahal 2018) по южной Азии в целом, состоящее из 2504 образцов мужской ДНК из Индии, Бангладеш, и Пакистана — выявило 11,2 % L-M20 (281 из 2504) .
Индия
Встречается чаще среди
дравидийских
каст (около 17-19 %), и реже среди
индоарийских
(около 5-6 %).
Может достигать 68 % в некоторых племенах и кастах
Карнатаки
,
38 % в некоторых кастах
Гуджарата
,
48 % в некоторых кастах
Тамилнада
, и 12 % по общей частоте в
Пенджабе
.
Более ранние исследования (например, Wells 2001) сообщают об очень высокой частоте (приближающейся к 80 %) гаплогруппы L-M20 в
Тамилнаде
, по-видимому, из-за экстраполяции данных полученных из выборки 84 калларов — тамило-говорящей высокой касты правителей Тамилнада, среди которых 40 (приблизительно 48 %) показали мутацию M20 определяющую гаплогруппу L. Присутствие гаплогруппы L-M20 редко встречается среди племенных групп (около 5,6-7 %).
68 % L-M20 было обнаружено у племени Корова из Карнатаки , 38 % в касте из округа Джунагадх в Гуджарате , 21 % в касте из округа Джунагадх в Гуджарате, 17 % в племени Каре Воккал (Kare Vokkal) из Уттара Каннада в Карнатаке. Также встречается с низкой частотой в других популяциях областей Джунагадх и Уттара Каннада. L-M20 имеет большая часть мужчин (36,8 %) среди Джатов в Северной Индии , и встречается у 16,33 % Гуджаров в Джамму и Кашмире . Обнаружена у 18,6 % и 15 % маратхов Махараштры . L-M20 также обнаружена у 32,35 % и у 17,82 % Лингаятов Карнатаки. L-M20 обнаружена на уровне 20,7 % среди , 16,7 % среди , и 17,2 % среди Тамилнада. L-M11 встречается частотой 8-16 % среди индийских евреев . 2 % L-M11 обнаружено у Сидди — народа Африканского происхождения. В целом гаплогруппа L-M20 в настоящее время присутствует в индийской популяции с общей частотой около 7-15 %.
Пакистан
Наибольшая концентрация L-M20 в Пакистане тянется вдоль реки
Инд
, где 3300-1300 лет до н. э. процветала
Индская (Хараппская) цивилизация
. Наибольшая частота и разнообразие L1a2-M357 обнаружены в
Белуджистане
— 28 %,
другая точка концентрации L1a2-M357 находится в горных изолированных долинах на севере страны, там он обнаружен у 16,5 %
Буришей
и 25 %
Калашей
.
У Пакистанских
Пуштунов
численность которых составляет около 32 млн чел.
процент M357 составил около 7 % (Firasat 2007),
исследование
Lee 2014
(270 человек) показало результат — 5,9 %
В среднем M357 имеет распространение 11,6 % по всему Пакистану.
Афганистан
Пуштуны
являются самым многочисленным народом Афганистана (около 14 млн человек)
. Исследование
(Lacau 2012)
ДНК 190 мужчин показало, что гаплогруппа L-M20 с общей частотой 9,5 % является второй по численности мужской линией среди них, субклад L1a2-M357 составил 7,5 % из 9,5 %.
Исследование демонстрирует существенную разницу распределения L-M20 по обеим сторонам хребта
Гиндукуш
— L выявлена у 25 % северных афганских пуштунов и 4,8 % южных. В частности, L1a2-M357 составляет большинство как на севере (20,5 %), так и на юге (4,1 %). В более раннем исследовании, в котором участвовало меньшее количество образцов, сообщалось, что L1a2-M357 составляет 12,24 % от общего количества афганских пуштунов.
Таджики — второй по численности народ в Афганистане (ок. 9-11 млн человек), сконцентрированный в северных провинциях страны. Выявлено 6,34 % L1a (9 из 142). Из них 5 L1a2 и 4 L1a1. Провинции — Балх (6 чел.), Бадахшан (2 чел.), Саманган (1 чел.)
У узбеков (ок. 4 млн чел ) выявлено 9,52 % L (12/126). Из них L1a1 — 6 чел, L1a2 — 5 чел. L1b — 1 чел. — северныe провинции Дзаузджан , Сари-Пуль , Балх (1 чел.)
Хазарейцы 2,97 % L-M20 (3/101) — 2 человека в пров. Балх и 1 человек в пров. Бамиан .
| Популяции | Распространение | Источники |
|---|---|---|
| Турция | 57 % — деревня Афшар, 12 % (10/83) — Черноморский регион, 6,6 % (7/106) — Турки из Турции. Также — 4,2 % (1/523 L-M349 и 21/523 L-M11(xM27, M349)) | , |
| Иран |
Самое обширное исследование
Grugni 2012
(938 мужчин,15 этносов, 14 провинций) показало результат — 5 % L-M20 (L1a1 — 1,8 % L1a2 — 1,5 % L1b — 1,5 % L* — 0,2 %)
Результаты других исследований, часть которых вошла в Grugni 2012 : 54,9 % (42/71) — у зороастрийских священников среди Парсов 22,2 % L1b и L1c в южном Иране (2/9) 8 % — 16 % L2-L595, L1a, L1b и L1c у Курдов Курдистана (2-4/25) 9,1 % L-M20 (7/77) у персов восточного Ирана 3,4 % L-M76 (4/117) и 2,6 % L-M317 (3/117), в целом 6,0 % (7/117) L-M20 в Южном Иране 3,0 % (1/33) L-M357 в северном Иране 4,2 % L1c-M357 Азербайджанцев в провинции восточный Азербайджан (1/21) 4,8 % L1a и L1b у Персов из Исфахана (2/42) |
, , Cristofaro 2013 , , |
| Сирия | 51.0 % (33/65) Сирийцев в Ракке , 31,0 % у восточных Сирийцев. В абсолютном большинстве L1b-M317 | |
| Саудовская Аравия | 15.6 % (4/32 из L-M76 и 1/32 of L-317) 1,91 % (2/157=1.27 % L-M76 и 1/157=0.64 % L-M357) | and |
| Курды |
Турция
3,2 % в юго-восточной Турции (Flores 2005) |
|
| Ирак |
3.1 % (2/64) L-M22 (Sanchez 2005)
Северный Ирак (Dogan 2017) Язиды — 10,09 % (11/109 все L1b-m317) Арабы — 3,70 % (4/108 L1a-m27 — 3 чел. L1b-m317 −1 чел) Курды — 2,73 % (3/110 L1a-M27 — 2 чел. L1c — 1 чел.) Туркмены 2,73 % (3/110 L1b-m317 — 2 L1a-m27 — 1) Ассирийцы — 1,09 % (1/92 L1a-m27) |
Dogan 2017 |
| Оман | 1 % L-M11 | |
| Катар | 2.8 % (2/72 L-M76) | |
| Арабы ОАЭ | 3.0 % (4/164 L-M76 и 1/164 L-M357) |
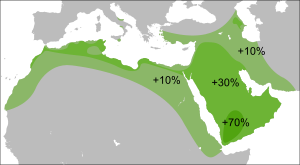
В небольшой выборке, взятой у друзов Израиля, гаплогруппа L была обнаружена у 7 человек из 20 (35 %). С другой стороны, исследования, проведенные на более широких выборках, показали, что мутация L-M20 в среднем составляет 5 % среди друзов Израиля , 4 % среди друзов Ливана . Кроме этого, она не была обнаружена вообще в выборке из 59 друзов Сирии. |
| Популяции | Распространение | Источник |
|---|---|---|
| Казахи | 1 % (3/300) L1c-m357 — 2 чел. в Жамбыльской и Восточно-Казахстанской областях. L1b-m317 — 1 чел. в Акмолинской обл. | |
| Таджики |
Таджикистан
7,74 % (13/168) Шугнан — 7\44 Ишкашим — 3\25 Душанбе — 2\16 Худжанд — 1\22 (Wells 2001). 22,5 % 9\40 — из них L1a2 — 4 (Malyarchuk 2013) Северный Афганистан 6,34 % L1a (9 из 142). Из них 5 L1a2 и 4 L1a1. Провинции — Балх (6 чел.), Бадахшан (2 чел.), Саманган (1 чел.) |
, Wells 2001 , Cristofaro 2013 |
| Узбеки |
Узбекистан
L-M20 3,0 % 11/366 (Wells 2001). L-M20 5,12 % (11/215) 5-Хорезм 4-Фергана 2-Ташкент, из них L1a2-M357 3 — 2-Фергана 1-Xорезм (Balanovska 2017) Северный Афганистан 9,52 % L (12/126). Из них L1a1 — 6 чел, L1a2 — 5 чел. L1b — 1 чел. — северныe провинции Балх , Дзаузджан , Сари-Пуль . |
Wells 2001, Cristofaro 2013, Zhabagin-Balanovska 2017 |
| Уйгуры | 16.7 % (1/6) L1c-M357 в Киргизстане | Cristofaro 2013 |
| Памирцы | 16 % (7/44) Шугнанцы , 12 % 3/25 Ишкашимцы, 0/30 Бартанги | Wells 2001 |
| Хазарейцы | 2.97 % L-M20 (3/101) — 2 человека в пров. Балх и 1 человек в пров. Бамиан | Cristofaro 2013 |
| Ягнобцы | 9.7 % (3/31) | Wells 2001 |
| Бухарские Арабы | 9.5 % (4/42) | Wells 2001 |
| Дунгане | 8.3 % (1/12) L1c в Киргизстане | Cristofaro 2013 |
| Уйгуры (Lopliks) | 7.8 % (5/64) L-M357 деревня Qarchugha, округ Lopnur, Синцзян | Liu 2018 |
| Каракалпаки | 4.5 % (2/44) | Wells 2001 |
| Уйгуры | 4.4 % (3/68) | Karafet 2001 and Hammer 2005 [ |
| Tуркмены |
Афганистан
4,1 % (3/74) L1a-M27 в Джаузджан |
Cristofaro 2013 |
| Челканы | 4.0 % (1/25) | Dulik 2012 and Dulik 2012 |
| Киргизы | 2.7 % (1/37) L1c сев. запад и 2,5 % (1/40) L1a в центре Киргизстана | Cristofaro 2013 |
| Казанские Татары | 2.6 % (1/38) | Wells 2001 |
| Хуэй | 1.9 % (1/54) | Karafet 2001 |
| Башкиры | 0.64 % (3/471) | Lobov 2009 |
| Популяции | Распространение | Источник |
|---|---|---|
| Армяне | от 1,63 % (12/734) до 4,3 % (2/47) | and |
| Лазы | 41.7 % (15/36) L1b-M317 | |
| Грузины | 20 % (2/10) в Гали , 14,3 % (2/14) в Чохатаури , 12,5 % (2/16) в Мартвили , 11,8 % (2/17) в Абаше , 11,1 % (2/18) в Багдати , 10 % (1/10) в Гардабани , 9,1 % (1/11) в Адигени , 6,9 % (2/29) в Омало, 5,9 % (1/17) в Гурджаани , 5,9 % (1/17) в Лентехи . Также — 1,5 % (1/66) L-M357(xPK3) , 1,6 % (1/63) L-M11 | , and |
| Дагестан | 9.5 % L1b-M317 (4/42) у Аварцев, 8,3 % (2/24) L1a2-M357 у Татов, 11,76 % (2/17) L1b-M317 у горских Евреев 3,7 % (1/27) Чамалинцев, | Yunusbaev 2006, Caciagli 2009, Karafet 2016 |
| Чеченцы и Ингуши |
Чеченцы
Из ненаучных данных: Более 10 % L1a2-M357 и около 1 % L1b-M317 (110 из 922 и 9 из 922 человек на 26.06. 2019 г.) в ДНК-проекте «Chechen-noahcho dna project» созданном на базе американской лаборатории FTDNA. 14 % (14/100) L1a2-M357 у чеченцев из Ауховского района Дагестана (Balanovsky 2011) Ингуши Из ненаучных данных: 7 % L1a2-M357 (20/277) в ДНК-проекте «Ingush DNA project» Все чеченцы и ингуши носители L1a2-M357 происходят от ветви возрастом 3300 лет, которая является западно-Азиатской и отстоит от средне и южно Азиатских ветвей генетической дистанцией в 6000-7000 лет. |
Chechen-Noahcho DNA project
Ingush DNA project Balanovsky 2011 |
| Популяция | Распространение | Источник |
|---|---|---|
| (Fascia). Италия | 19.2 % L-M20 | Valentina Coia 2013 |
| (Nonstal). Италия | 10 % L-M20 | F. di Giacomo 2003 |
| Южный Тироль, Италия | 8.9 % Ладино-язычных из 8,3 % Валь Бадия, 2,9 % долина Пустерталь , 2,2 % германоязычных из Валь Бадия, 2 % германоязычных в верхнем Финшгау , 1,9 % германоязычных в нижнем Финшгау и 1,7 % Итальяноязычных в Больцано | Pichler 2006 and Thomas 2008 |
| Самний , Италия | 10 % Аквиланцев L-M20 | Alessio Boattini 2013 |
| Виченца, Италия | 10 % Венецианцев L-M20 | Alessio Boattini 2013 |
| Восточный Тироль, Австрия | L-M20 1,9 % у Тирольцев в регионе Б (Isel, Lower Drau, Defereggen, Virgen, Kals valley) | H.Niederstätter 2012 |
| Северный Тироль, Австрия | L-M20 0,8 % Ройтте | D.Erhart 2012 |
| Архангельская область, Россия | 5.9 % Русских L1c-M357 ( Нет результатов исследования в открытом виде, требуется уточнение информации ) | Hongyang Xu 2014 |
| Португалия | 5.0 % в Коимбра | Beleza 2006 |
| Болгария | 3.9 % у Болгар | Karafet 2016 |
| Фландрия, Бельгия | L1a*: 3,17 % в Мехелен 2,4 % в Тюрнхаут and 1,3 % в Де-Кемпен . L1b*: 0,74 % на западе и востоке | Larmuseau 2010 and Larmuseau 2011 |
| Гипускоа , Испания | L1b 1,7 % | Young 2011 |
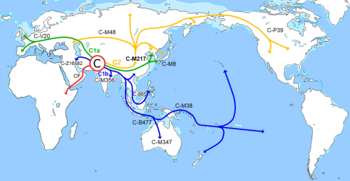
| Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||||||||||||||||||||||||||
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||||
| A0-T | |||||||||||||||||||||||||||||||||
| A00 | A0 | A1 | |||||||||||||||||||||||||||||||
| A1a | |||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||
| F1 F2 F3 | |||||||||||||||||||||||||||||||||
| G | |||||||||||||||||||||||||||||||||
| H | IJK | ||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||
| I | J | K2 | |||||||||||||||||||||||||||||||
| T (K1b) | K2a/K2a1/ NO /NO1 | ||||||||||||||||||||||||||||||||
| N | O | P (K2b2) /P1 | |||||||||||||||||||||||||||||||
| S (K2b1a) | M (K2b1b) | Q | R | ||||||||||||||||||||||||||||||
| Главные темы | |
|---|---|
| Города и поселения | |
| Соседи | |
| Сельское хозяйство | |
| Культура | |
| Народы, лингвистика | |
| Ключевые понятия |

|
|
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
| Гаплогруппа L-M20 | |
|---|---|

|
|
| Тип | Y-ДНК |
| Время появления |
25-30 тыс. лет назад
45500-39700 по yfull.com |
| Место появления | Южная Азия , Западная Азия , Памир |
| Время до БОП | 24000 лет назад |
| Предковая группа | K > LT |
| Сестринские группы | T |
| Субклады |
L1a1-M27,M76 (L1,L1a до 2015 года)
L1a2-M357,L1307 (L3, L1c до 2015 года), Гаплогруппа L3 (Y-ДНК) L1a3-M2533* носитель ветви выявлен в 2020-ом году L1b-M317 (L2 до 2015 года) L2-L595 (открыт в 2014 году) |