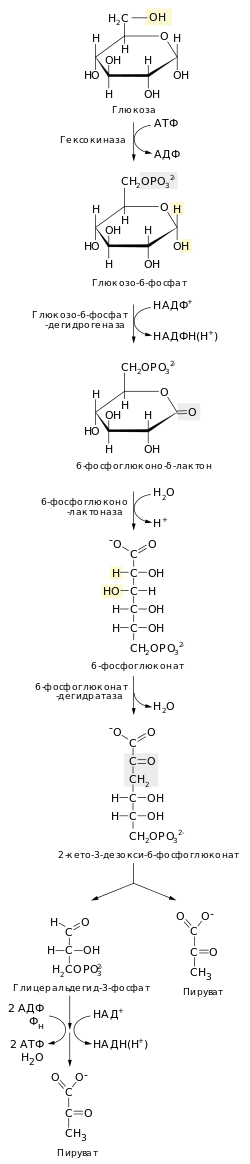
Путь Энтнера — Дудорова
- 1 year ago
- 0
- 0

Глико́лиз , или путь Эмбдена — Мейергофа — Парнаса (от греч. γλυκός — сладкий и греч. λύσης — расщепление) — процесс окисления глюкозы , при котором из одной молекулы глюкозы образуются две молекулы пировиноградной кислоты . Гликолиз состоит из цепи последовательных ферментативных реакций и сопровождается запасанием энергии в форме АТФ и НАДH . Гликолиз является универсальным путём катаболизма глюкозы и одним из трёх (наряду с пентозофосфатным путём и путём Энтнера — Дудорова ) путей окисления глюкозы, встречающихся в живых клетках . Реакция гликолиза в суммарном виде выглядит следующим образом:
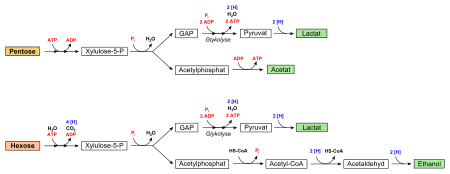
Кислород не требуется для протекания гликолиза. В аэробных условиях пировиноградная кислота далее декарбоксилируется , соединяется с коферментом А и вовлекается в цикл Кребса . В анаэробных условиях (при гипоксии ) пируват восстанавливается до молочной кислоты либо претерпевает дальнейшие превращения в ходе брожения .

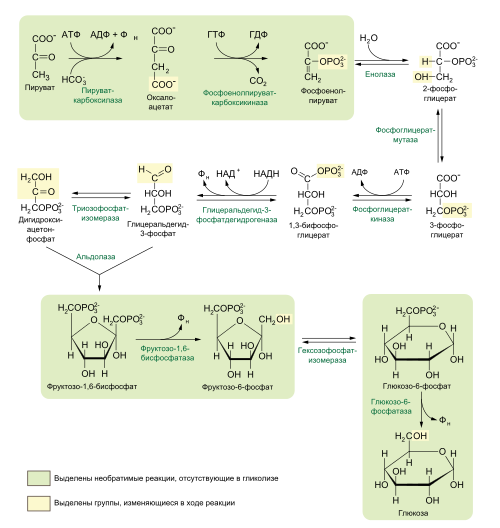
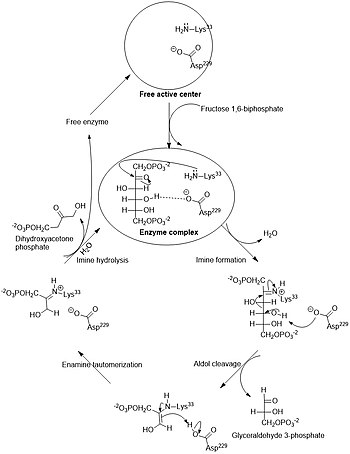
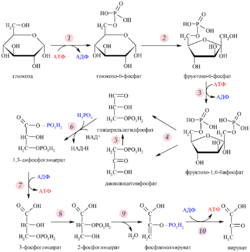
Распад шести углеродного сахара глюкозы на две молекулы трёхуглеродного пирувата осуществляется в 10 стадий, первые 5 которых составляют подготовительный этап с затратой АТФ, а 5 последующих — этап, сопряжённый с образованием АТФ . Все сахара и их производные, образующиеся при гликолизе, являются D-изомерами . В ходе реакций гликолиза глюкоза сначала фосфорилируется по гидроксильной группе при шестом атоме углерода (C-6), давая глюкозо-6-фосфат ( стадия 1 ). Глюкозо-6-фосфат затем изомеризуется в фруктозо-6-фосфат ( стадия 2 ), который вновь фосфорилируется, на этот раз по гидроксильной группе при первом атоме углерода, при этом образуется фруктозо-1,6-бисфосфат ( стадия 3 ). В ходе обеих этих реакций фосфорилирования донором фосфорильной группы является АТФ. Далее фруктозо-1,6-бисфосфат расщепляется на две трёхуглеродные молекулы — дигидроксиацетонфосфат и глицеральдегид-3-фосфат ( стадия 4 ), эта стадия и дала название всему пути. Дигидроксиацетонфосфат изомеризуется в глицеральдегид-3-фосфат ( стадия 5 ), так что к концу подготовительного этапа из глюкозы образуется 2 молекулы глицеральдегид-3-фосфата, которые в дальнейшем претерпевают одинаковые превращения. Изомеризация на стадии 2 необходима для дальнейшего фосфорилирования, а также разрыва связи С—С на стадии 4, как будет подробнее показано в дальнейшем. При этом в подготовительной стадии гликолиза расходуется 2 молекулы АТФ, что увеличивает свободную энергию промежуточных соединений пути .
Энергетическую выгоду даёт второй этап гликолиза, сопряжёный с образованием АТФ. Каждая из двух молекул глицеральдегид-3-фосфата окисляется и фосфорилируется фосфорной кислотой ( а не АТФ ), образуя 1,3-бисфосфоглицериновую кислоту ( стадия 6 ). Выделение энергии происходит при превращении двух молекул 1,3-бисфосфоглицерата в две молекулы пирувата ( стадии 7—10 ), и большая часть этой энергии запасается при присоединении фосфатной группы к четырём молекулам АДФ с образованием четырёх молекул АТФ. Суммарный выход составляет 2 молекулы АТФ на молекулу глюкозы, поскольку 2 молекулы АТФ расходуются в подготовительном этапе. Кроме того, во втором этапе гликолиза часть энергии запасается при образовании двух молекул восстановленного НАДH на одну молекулу глюкозы .
Таким образом, гликолиз включает в себя химические перестройки следующего типа:
Итак, суммарное уравнение гликолиза:
Каждое из 9 промежуточных соединений на пути от глюкозы к пирувату содержат остатки ортофосфорной кислоты . По-видимому, фосфатные группы в этом случае выполняют следующие 3 функции:
На подготовительном этапе гликолиза шестиуглеродная молекула глюкозы расщепляется на два триозофосфата. При этом затрачиваются две молекулы АТФ . Подготовительный этап гликолиза включает 5 реакций, которые подробно описаны ниже.
В первой реакции гликолиза происходит активация молекулы глюкозы путём её фосфорилирования по шестому атому углерода (С-6) с образованием глюкозо-6-фосфата , при этом донором фосфорильной группы выступает АТФ :

| Фермент | Кофактор |
Изменение свободной энергии
(ΔG′ о , кДж/моль) |
| Гексокиназа | Mg 2+ | −16,7 |
Эта реакция катализируется ферментом гексокиназой (в клеточных условиях гексокиназа неспособна осуществлять обратную реакцию). Он относится к группе киназ — ферментов, катализирующих перенос терминальной фосфорильной группы с АТФ на акцептор — нуклеофил . В случае гексокиназы акцептором является гексоза, как правило, D-глюкоза, хотя в некоторых тканях гексокиназа также может катализировать фосфорилирование и других распространённых гексоз, например, D- фруктозы и D- маннозы (подробнее см. ниже).

Как и многим другим киназам, гексокиназе для активности необходимо присутствие ионов Mg 2+ , поскольку собственно субстратом для этого фермента является не АТФ 4- , а комплекс MgАТФ 2- . Ион магния «закрывает собой» часть отрицательного заряда фосфатных групп АТФ, делая терминальный атом фосфора более доступным для нуклеофильной атаки гидроксильной группой глюкозы. При связывании с глюкозой гексокиназа значительно изменяет конфигурацию, два её домена при связывании с АТФ сближаются друг с другом на 8 Å . Такое сближение подводит связанный с ферментом АТФ ближе к молекуле глюкозы, также с ним связанной, а также блокирует вход в активный центр воды из раствора, которая в противном случае гидролизовала бы фосфоангидридные связи в молекуле АТФ. Как и другие 9 ферментов гликолиза, гексокиназа является растворимым цитозольным белком .
Человеческий геном кодирует 4 различные гексокиназы (I—IV), которые катализируют одну и ту же реакцию (два и более фермента, катализирующие одну и ту же реакцию, но кодируемые разными генами , называются Изоферментами ). Гексокиназа IV, также называемая глюкокиназой , присутствует в гепатоцитах и отличается от других гексокиназ некоторыми кинетическими и регуляторными свойствами и играет важную физиологическую роль .
Фермент , или фосфоглюкозоизомераза катализирует обратимую изомеризацию глюкозо-6-фосфата ( альдозы ) во фруктозо-6-фосфат ( кетозу ) :

| Фермент | Кофактор |
Изменение свободной энергии
(ΔG′ о , кДж/моль) |
|
Фосфогексозоизомераза,
или глюкозоизомераза |
Mg 2+ | 1,7 |
Механизм этой реакции включает в себя образование промежуточного енодиольного соединения. Эта реакция одинаково хорошо идёт в обоих направлениях, как и можно было предположить из её небольшого ΔG′ о . Эта изомеризация играет ключевую роль для всех последующих превращений гликолиза, поскольку перестройка карбонильной и гидроксильной групп при С-1 и С-2 необходима для следующих двух стадий. Для фосфорилирования, происходящего в следующей стадии, необходимо, чтобы при С-1 карбонильная группа была перестроена в гидроксильную, а для четвёртой стадии — разрыва связи между С-3 и С-4 — необходимо наличие карбонильной группы при С-2 .
В третьей реакции гликолиза, протекающей с затратой АТФ, фермент фосфофруктокиназа-1 катализирует перенос фосфорильной группы от АТФ на фруктозо-6-фосфат с образованием фруктозо-1,6-бисфосфата :

| Фермент | Кофактор |
Изменение свободной энергии
(ΔG′ о , кДж/моль) |
| Фосфофруктокиназа-1 | Mg 2+ | −14,2 |
В клеточных условиях фосфофруктокиназа не может осуществлять эту реакцию в обратном направлении, и эта реакция является первой реакцией, продукт которой (фруктозо-1,6-бисфосфат) участвует только в дальнейших реакциях гликолиза, потому что глюкозо-6-фосфат и фруктозо-6-фосфат могут участвовать и в других [ каких? ] процессах .
У некоторых, как правило анаэробных, бактерий и протистов фосфофруктокиназа в качестве донора фосфорильной группы для образования фруктозо-1,6-бисфосфата использует пирофосфорную кислоту (PP i ), а не АТФ:
В растительных клетках имеется как АТФ-зависимая фосфофруктокиназа, так и (реакция, катализируемая последней, обратима) . Пирофосфат-зависимая фосфофруктокиназа локализована в цитозоле и активируется в условиях стресса, при дефиците АТФ (например, при аноксии ) и .
Фосфофруктокиназа-1 регулируется аллостерически . Её активность увеличивается, когда клеточные запасы АТФ истощаются, а продукты распада АТФ (АДФ и АМФ ) накапливаются. Напротив, при наличии достаточного количества АТФ и других [ каких? ] ресурсов её активность подавляется. У некоторых организмов фруктозо-2,6-бисфосфат является потенциальным аллостерическим регулятором фосфофруктокиназы 1. Косвенным образом активность этого фермента увеличивает также (промежуточное соединение пентозофосфатного пути , другого пути окисления глюкозы) (подробнее о регуляции ферментов гликолиза см. ниже).
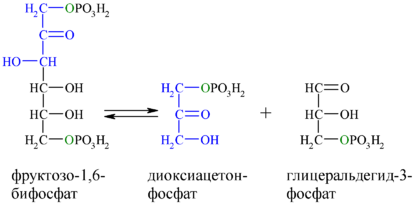
Фермент фруктозо-1,6-бисфосфатальдолаза , или просто альдолаза , катализирует обратимую альдольную конденсацию . Фруктозо-1,6-бисфосфат расщепляется на два различных триозофосфата: глицеральдегид-3-фосфат (альдозу) и дигидроксиацетонфосфат (кетозу) :

| Фермент | Кофактор |
Изменение свободной энергии
(ΔG′ о , кДж/моль) |
| Альдолаза | 23,8 |
Существуют 2 класса альдолаз. Альдолазы класса I имеются у животных и растений , их работа сопровождается образованием промежуточного основания Шиффа . Альдолазы класса II имеются у грибов и бактерий , при их работе промежуточных оснований Шиффа не образуется. Вместо этого ион цинка в активном сайте фермента связывается с атомом кислорода карбонильной группы при С-2. Ион Zn 2+ поляризует карбонильную группу и стабилизирует енольное промежуточное соединение, образующееся при разрыве связи С—С .
Хотя реакция, катализируемая альдолазой , имеет положительную ΔG′ о в направлении расщепления фруктозо-1,6-бисфосфата, при низких концентрациях реагентов, имеющихся в клетке, реальное изменение свободной энергии мало и альдолазная реакция обратима. В обратном направлении альдолазная реакция идёт при глюконеогенезе .
Лишь один из двух продуктов альдолазной реакции — глицеральдегид-3-фосфат — может участвовать в дальнейших превращениях гликолиза. Другой продукт — дигидроксиацетонфосфат — быстро и обратимо переводится в глицеральдегид-3-фосфат ферментом :

| Фермент | Кофактор |
Изменение свободной энергии
(ΔG′ о , кДж/моль) |
| Триозофосфатизомераза | 7,5 |
Механизм этой реакции схож с механизмом реакции, катализируемой фосфогексозоизомеразой на стадии 2. Таким образом, обе «половины» глюкозы превращаются в глицеральдегид-3-фосфат . Эта реакция завершает подготовительный этап гликолиза. К его концу молекула глюкозы, фосфорилированная по С-1 и С-6, расщепляется на две молекулы глицеральдегид-3-фосфата .
Второй этап гликолиза содержит стадии, в которых часть химической энергии молекулы глюкозы запасается в виде АТФ за счёт субстратного фосфорилирования АДФ, а также образования НАДH. Две молекулы глицеральдегид-3-фосфата, образовавшиеся в ходе подготовительного этапа гликолиза, во втором этапе подвергаются одинаковым превращениям. В конечном счёте каждая из них переводится в пируват, при этом образуется 4 молекулы АТФ. Однако суммарный выход АТФ в гликолизе составляет 2 молекулы, так как 2 молекулы АТФ расходуются в подготовительном этапе .
В первой реакции второго этапа гликолиза молекула глицеральдегид-3-фосфата окисляется и фосфорилируется в 1,3-бисфосфоглицерат , эта реакция катализируется :

| Фермент | Кофактор |
Изменение свободной энергии
(ΔG′ о , кДж/моль) |
| Глицеральдегид-3-фосфатдегидрогеназа | 6,3 |
Это первая из двух энергозапасающих реакций, продукты которых в дальнейшем участвуют в образовании АТФ. Альдегидная группа глицеральдегид-3-фосфата окисляется, но не до свободной карбоксильной группы , а до ангидрида карбоновой кислоты с фосфорной кислотой. Ангидрид такого типа — ацилфосфат — имеет очень высокую стандартную энергию гидролиза (ΔG′ о = −49,3 кДж/моль). Большая часть свободной энергии окисления альдегидной группы глицеральдегид-3-фосфата запасается при образовании ацилфосфатной группы при С-1 1,3-бисфосфоглицерата .
В ходе этой реакции глицеральдегид-3-фосфат ковалентно связан с дегидрогеназой. Альдегидная группа глицеральдегид-3-фосфата взаимодействует с группой —SH остатка цистеина в активном центре фермента. Когда глицеральдегид-3-фосфат находится в связанном состоянии, NAD + , также находящийся в активном центре фермента, забирает протон от С-1, в результате чего там образуется кетогруппа . К первому же атому на место связи с атомом серы цистеина присоединяется неорганический фосфат HOPO 3- , и протон с фосфата высвобождается во внешнюю среду. Таким образом, после этой реакции образуется 1,3-бисфосфоглицерат и NADH + Н + .
Количество NAD + в клетке (< 10 −5 М) гораздо меньше, чем количество глюкозы, расщеплямой за несколько минут. Если NADH, образующийся на этой стадии гликолиза, не будет постоянно расходоваться (то есть окисляться), то гликолиз останавливается .
Фермент фосфоглицераткиназа переносит высокоэнергетическую фосфорильную группу с карбоксильной группы 1,3-бисфосфоглицерата на АДФ , в результате чего образуются АТФ и 3-фосфоглицерат :

| Фермент | Кофактор |
Изменение свободной энергии
(ΔG′ о , кДж/моль) |
| Фосфоглицераткиназа | Mg 2+ | −18,5 |
Этот фермент получил своё название за обратную реакцию, при которой происходит перенос фосфатной группы с АТФ на 3-фосфоглицерат . Он катализирует оба направления реакции. Реакцию фосфорилирования 3-фосфоглицерата он катализирует при глюконеогенезе и при фотосинтетическом поглощении СО 2 .
Стадии 6 и 7 гликолиза с энергетической точки зрения рассматриваются вместе [ кем? ] и образуют единый процесс, при котором 1,3-бисфосфоглицерат является промежуточным продуктом. Он образуется при первой из этих реакций (которая сама по себе эндергоническая ), а его фосфатная группа переносится на АДФ при второй, строго экзергонической , реакции. Суммарное уравнение процесса, объединяющего стадии 6 и 7, выглядит следующим образом:
Поэтому стадии 6 и 7 вместе составляют экзергонический процесс. Обе эти реакции обратимы при клеточных условиях, и составляемый ими процесс обеспечивает запасание энергии, образовавшейся при окислении альдегидной группы до карбоксильной, в форме АТФ при его образовании из АДФ и фосфорной кислоты. Образование АТФ при переносе фосфорильной группы с субстрата (в данном случае — 1,3-бисфосфоглицерата ) на АДФ получило название субстратного фосфорилирования , в отличие от окислительного фосфорилирования, происходящего в дыхательной цепи. При субстратном фосфорилировании участвуют растворимые ферменты и химические промежуточные соединения (в данном случае — 1,3-бисфосфоглицерат ), а в окислительном фосфорилировании задействованы мембранно-связанные белки , и АТФ образуется за счёт трансмембранного протонного градиента .
Фермент фосфоглицератмутаза катализирует обратимый перенос фосфорильной группы с С 3 на С 2 глицерата, приводящий к образованию 2-фосфоглицериновой кислоты . Для этой реакции необходим Mg 2+ :

| Фермент | Кофактор |
Изменение свободной энергии
(ΔG′ о , кДж/моль) |
| Фосфоглицератмутаза | Mg 2+ | 4,4 |
Реакция осуществляется в два этапа. Вначале фосфорильная группа, связанная с остатком гистидина в активном центре фосфоглицератмутазы, замещает собой атом водорода в гидроксильной группе при С 2 углероде 3-фосфоглицерата , образуя 2,3-бисфосфоглицерат , который связывается другим гистидиновым остатком. Фосфорильная группа при С 3 углероде 2,3-бисфосфоглицерата после этого перемещается на остаток гистидина, с которым был связан фосфат, перенесённый на С 2 , а её место замещается протоном, связанным со вторым остатком гистидина. Таким образом, к концу такого цикла образуется 2-фосфоглицерат, а фермент фосфорилируется .
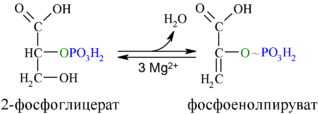
Во второй реакции гликолиза, в которой образуется соединение с более высоким потенциалом для перенесения фосфата (первой была стадия 6), фермент енолаза катализирует обратимое элиминирование воды ( дегидратацию ) из молекулы 2-фосфоглицерата, в результате которого образуется фосфоенолпировиноградная кислота (ФЕП) :
| Фермент | Кофактор |
Изменение свободной энергии
(ΔG′ о , кДж/моль) |
| Енолаза | Mg 2+ | 7,5 |
Механизм реакции, катализируемой енолазой, включает стабилизацию промежуточного продукта ионами Mg 2+ . В ходе этой реакции соединение с относительно низким потенциалом для транспорта фосфата (ΔG′ о при гликолизе 2-фосфоглицерата составляет −17,6 кДж/моль) в соединение с высоким потенциалом для транспорта фосфата (ΔG′ о при гликолизе ФЕП составляет −61,9 кДж/моль) .
В последней реакции гликолиза происходит перенос фосфорильной группы с фосфоенолпирувата на АДФ, катализируемый пируваткиназой , для работы которой необходимы ионы К + и Mg 2+ или Mn 2+ :

| Фермент | Кофактор |
Изменение свободной энергии
(ΔG′ о , кДж/моль) |
| Пируваткиназа |
К
+
,
Mg 2+ /Mn 2+ |
−31,4 |
Таким образом, эта реакция представляет собой субстратное фосфорилирование . Её продукт — пируват — вначале образуется в енольной форме, которая затем быстро таутомеризуется в кето-форму, которая преобладает при pH = 7 (то есть в клеточных условиях) .
Суммарно эта реакция имеет большое отрицательное изменение свободной энергии из-за спонтанной конверсии енольной формы пирувата в кето-форму. Около половины энергии, выделившейся при гидролизе ФЕП (ΔG′ о = −30,5 кДж/моль), запасается при образовании фосфоангидридной связи в АТФ (ΔG′ о = −30,5 кДж/моль), а остальная энергия (-31,4 кДж/моль) составляет мощную движущую силу для направления этой реакции в сторону образования АТФ .
С энергетической точки зрения в гликолизе можно выделить 2 процесса:
1) Превращение глюкозы в пируват — энергетически выгодный процесс:
2) Образование ATP из ADP и 2P i — энергетически невыгодный процесс:
Общее изменение энергии Гиббса при гликолизе ΔG′ s составляет:
Поэтому при нормальных условиях, а также клеточных условиях (отличных от нормальных) гликолиз является в значительной мере необратимым процессом благодаря значительному уменьшению свободной энергии системы .
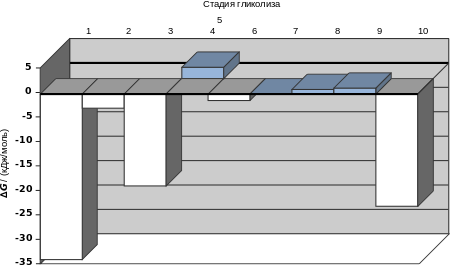
Из представленной выше диаграммы видно, что только три реакции (1, 3 и 10) протекают с высоким изменением свободной энергии, причём равновесие сильно смещено в сторону образования конечных продуктов, а другие реакции легко обратимы. При глюконеогенезе они могут идти в противоположном направлении, причём их будут катализировать те же ферменты, что и при гликолизе. Для необратимых реакций 1, 3 и 10 в глюконеогенезе используются обходные пути .
Многие углеводы , отличные от глюкозы, также разрушаются по пути гликолиза, но после того, как они будут переведены в одно из промежуточных соединений гликолиза .

Полимеры глюкозы гликоген , запасаемый в тканях животных, и крахмал , запасаемый растениями, могут быть использованы клеткой для получения энергии при помощи гликогенолиза — фосфоролитической реакции, осуществляемой гликогенфосфорилазой (или у растений). Эти ферменты катализируют атаку гликозидной связи (α1→4), соединяющей два крайних остатка глюкозы на неветвящемся конце, фосфат-ионом, в результате чего образуется глюкозо-1-фосфат и полимер глюкозы, содержащий на фрагмент глюкозы меньше, чем исходный. Часть энергии гликозидной связи при этом запасается в виде эфирной связи, соединяющей фосфат с глюкозой в глюкозо-1-фосфате. Фосфорилаза продолжает отщеплять по одному остатку глюкозы до тех пор, пока не дойдёт до точки ветвления полисахарида (гликозидной связи (α1→6)), где она останавливается. Глюкозо-1-фосфат переводится в глюкозо-6-фосфат ферментом фосфоглюкомутазой , катализирующим обратимую реакцию:
Механизм действия этого фермента такой же, как у фосфоглицератмутазы. Образующийся в ходе этой реакции глюкозо-6-фосфат может далее быть задействован в гликолизе или пентозофосфатном пути .
Описанная выше ситуация характерна лишь для гликогена и крахмала, запасённых внутри клетки. Фосфоролиз гликогена и крахмала, поступающих в организм с пищей, в пищеварительном тракте не имеет никаких преимуществ перед обычным гидролизом : так как клеточные мембраны непроницаемы для фосфатов сахаров, образующийся при глюкозо-6-фосфат необходимо сначала превратить в обычный сахар . При гидролизе, осуществляемом, например, пищеварительным ферментом α-амилазой , частицей, атакующей гликозидную связь, является вода, а не фосфат-ион .
Дисахариды до их проникновения в клетку предварительно гидролизуются до соответствующих моносахаридов . В пищеварительном тракте такой гидролиз осуществляют ферменты, прикреплённые к поверхности клеток пищеварительного эпителия (в скобках указан фермент, катализирующий соответствующую реакцию):
Образующиеся моносахариды активно транспортируются в эпителиальные клетки, затем попадают в кровь и разносятся к различных тканям , где фосфорилируются и вовлекаются в гликолиз .
У большинства организмов гексозы, отличные от глюкозы, вовлекаются в гликолиз после преобразования в фосфорилированное производное. Гликолитическое расщепление фруктозы называется . D-Фруктоза, в свободном виде присутствующая во многих фруктах и образующаяся при гидролизе сахарозы в тонкой кишке у позвоночных , фосфорилируется гексокиназой:
Такой путь является основным механизмом вовлечения фруктозы в гликолиз в мышцах и почках . В печени она вовлекается в гликолиз иначе. Фермент печени фруктокиназа катализирует фосфорилирование фруктозы по С-1, а не С-6:
Далее фруктозо-1-фосфат расщепляется на глицеральдегид и дигидроксиацетонфосфат ферментом фруктозо-1-фосфатальдолазой . Далее дигидроксиацетонфосфат превращается в глицеральдегид-3-фосфат гликолитическим ферментом триозофосфатизомеразой, а глицеральдегид фосфорилируется ATP и ферментом до глицеральдегид-3-фосфата:
Получившиеся 2 молекулы глицеральдегид-3-фосфата вовлекаются в гликолиз . Ниже представлена схема вышеописанных процессов:

D-Галактоза, продукт гидролиза лактозы, из кишечника всасывается в кровь , откуда попадает в печень , где фосфорилируется галактокиназой по С-1 с затратой АТФ:
Галактозо-1-фосфат далее по С-4 в глюкозо-1-фосфат в серии реакций, в которых (UDP) функционирует как коферментоподобный переносчик гексоз. Эпимеризация включает сначала окисление гидроксильной группы при С-4 до кетогруппы, а затем обратное восстановление кетогруппы до гидроксильной с обращённой конфигурацией. В этих двух реакциях окисления и восстановления кофактором выступает NAD . Ниже представлена схема описанного процесса:


D- Манноза , образующаяся при пищеварительном расщеплении многих полисахаридов и гликопротеинов , может быть фосфорилирована по С-6 гексокиназой:
далее изомеризуется ферментом до фруктозо-6-фосфата — промежуточного соединения гликолиза .
Регуляция гликолиза обычно осуществляется совместно с регуляцией обратного процесса — глюконеогенеза . У млекопитающих глюконеогенез протекает в основном в печени, где его функция заключается в синтезе глюкозы для перенесения к другим тканям в ситуациях, когда запасы гликогена истощены и с пищей в организм не поступает достаточного количества глюкозы. Как упоминалось выше, благодаря обратимости семи из десяти реакций гликолиза в ходе глюконеогенеза эти реакции протекают в обратном направлении и при катализе теми же ферментами, а для необратимых реакций (1, 3 и 10) используются обходные пути. Эти обходные реакции также необратимы. Так, при глюконеогенезе пируват переходит в фосфоенолпируват через промежуточную стадию образования оксалоацетата при катализе , превращающей пируват в оксалоацетат, и , переводящей оксалоацетат в фосфоенолпируват (обходной путь для десятой стадии). Обходная реакция для третьей стадии — превращение фруктозо-1,6-бисфосфата во фруктозо-6-фосфат ферментом фруктозо-1,6-бисфосфатазой , а для первой стадии — превращение глюкозо-6-фосфата в глюкозу глюкозо-6-фосфатазой .
Гексокиназа, катализирующая фосфорилирование глюкозы на стадии 1, в организме человека представлена четырьмя изоформами (I—IV). Они кодируются разными генами [ какими? ] , но катализируют одну и ту же реакцию. В миоцитах преобладает гексокиназа II , имеющая высокое сродство к глюкозе, — её полунасыщение наступает при 0,1 мМ глюкозы. Поскольку в миоцит глюкоза попадает из крови, где концентрация глюкозы составляет 4—5 мМ, то внутри клетки поддерживается концентрация глюкозы, достаточная для насыщения гексокиназы II, и этот фермент работает в полную силу. Мышечные гексокиназы I и II аллостерически ингибируются их продуктом, глюкозо-6-фосфатом, так что при повышении внутриклеточной концентрации глюкозо-6-фосфата выше нормального уровня происходит временное обратимое подавление активности этих ферментов. Таким образом скорость образования глюкозо-6-фосфата находится в балансе со скоростью его расщепления .
Изоформы гексокиназы играют различные роли в углеводном метаболизме печени и мышц: в мышцах глюкоза потребляется для получения энергии, а печень поддерживает концентрацию глюкозы в крови, потребляя глюкозу или образуя её путём глюконеогенеза в зависимости от концентрации. В печени преобладает гексокиназа IV (глюкокиназа), которая отличается от мышечных гексокиназ I—III в трёх важных аспектах. Во-первых, концентрация глюкозы, при которой происходит полунасыщение гексокиназы IV, составляет около 10 мМ, что выше обычной концентрации глюкозы в крови. Быстрое выравнивание концентраций глюкозы в цитозоле гепатоцитов и крови обеспечивает наличие в мембранах гепатоцитов белков-транспортёров глюкозы — GLUT2 . Когда уровень глюкозы в крови повышен, например, после потребления пищи, богатой углеводами, излишек глюкозы переносится в гепатоциты, где гексокиназа IV превращает её в глюкозо-6-фосфат. Так как гексокиназа IV не насыщена при 10 мМ глюкозы, её активность продолжает расти при повышении концентрации глюкозы до 10 мМ и более. При низкой концентрации глюкозы в крови её концентрации в гепатоцитах недостаточно для работы гексокиназы IV, и глюкоза, образовавшаяся в ходе глюконеогенеза, покидает клетку и не фосфорилируется. Во-вторых, гексокиназа IV не подавляется глюкозо-6-фосфатом и поэтому может продолжать работать даже тогда, когда накопление глюкозо-6-фосфата полностью подавляет гексокиназы I—III. Наконец, в-третьих, гексокиназа IV подавляется при обратимом связывании с ней регуляторного белка [ какого? ] , имеющегося только в печени. Этот белок наиболее эффективно связывается с гексокиназой IV в присутствии аллостерического регулятора — фруктозо-6-фосфата . Однако за связывание фруктозо-6-фосфата с этим белком конкурирует глюкоза, которая при связывании с ним вызывает диссоциацию комплекса этого белка и фермента и ослабляет подавление его активности. Сразу после приёма насыщенной углеводами пищи, когда уровень глюкозы в крови высок, глюкоза входит в гепатоциты с помощью GLUT2 и активирует гексокиназу IV по вышеописанному механизму. При голодании, когда уровень глюкозы в крови становится ниже 5 мМ, фруктозо-6-фосфат активирует подавление гексокиназы IV с помощью этого регуляторного белка, в результате чего печень не конкурирует с другими органами за потребление глюкозы. Этот механизм ингибирования при помощи регуляторного белка интересен ещё и тем, что этот белок фиксирует гексокиназу IV внутри ядра клетки, так что она оказывается отделённой от других ферментов гликолиза, локализованных в цитозоле. При повышении концентрации глюкозы в цитозоле она выравнивается с концентрацией глюкозы в ядре путём транспорта через ядерные поры . Глюкоза вызывает диссоциацию регуляторного белка, гексокиназа IV выходит в цитозоль и начинает фосфорилировать глюкозу .
Гексокиназа IV и глюкозо-6-фосфатаза также регулируются на уровне транскрипции (подробнее о транскрипционной регуляции см. ниже) .

Как уже отмечалось, глюкозо-6-фосфат может вовлекаться как в гликолиз, так и в другие процессы, в том числе в синтез гликогена и пентозофосфатный путь. Метаболически необратимая реакция, катализируемая фосфофруктокиназой-1 (PFK-1) является этапом, строго закрепляющим участие данной молекулы глюкозы только в гликолизе. Кроме субстрат-связывающих сайтов, этот сложно устроенный фермент имеет несколько регуляторных сайтов, с которыми связываются аллостерические активаторы или ингибиторы .
ATP является не только субстратом для PFK-1, но и конечным продуктом гликолиза. Когда высокий уровень ATP в клетке сигнализирует о том, что образование ATP превосходит его потребление, ATP связывается с PFK-1 в особом аллостерическом сайте и снижает сродство этого фермента к субстрату — фруктозо-6-фосфату . АДФ и АМФ , концентрация которых возрастает, когда потребление ATP опережает его образование, аллостерически связываются с PFK-1 и уменьшают ингибиторное действие связанного с этим ферментом ATP. Эти механизмы способствуют увеличению активности фермента при накоплении ADP или AMP и понижению — при накоплении ATP .
Цитрат , ключевое промежуточное соединение при аэробном окислении пирувата, жирных кислот и аминокислот, также является аллостерическим регулятором PFK-1. Высокая концентрация цитрата увеличивает ингибиторный эффект ATP, дополнительно уменьшая расщепление глюкозы в ходе гликолиза. В этом случае, как и в некоторых других, описанных ниже, цитрат выступает в роли внутриклеточного сигнала, свидетельствующего об удовлетворении клеткой своих энергетических потребностей при окислении жиров и белков .
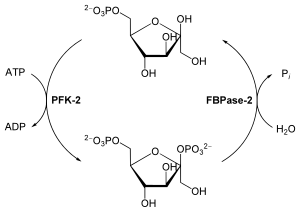
Реакции, катализируемой PFK-1, в гликолизе соответствует реакция глюконеогенеза, при которой фруктозо-1,6-бисфосфат переводится во фруктозо-6-фосфат. Эта реакция катализируется ферментом фруктозобисфосфатазой-1 (FBPаза-1). FBPаза-1 строго подавляется аллостерическим связыванием АМФ, так что когда клеточные запасы АТФ невелики, а уровень АМФ высок, АТФ-зависимый синтез глюкозы приостанавливается .
Таким образом, противоположные этапы гликолиза и глюконеогенеза, катализируемые PFK-1 и FBPазой-1 соответственно, регулируются координированно и реципрокно (то есть обратно). Вообще, при достаточных концентрациях ацетил-CoA или цитрата (продукт конценсации ацетил-CoA с оксалоацетатом в цикле Кребса ) или когда большая доля клеточного аденилата находится в форме ATP, предпочтительным процессом является глюконеогенез. При повышении уровня AMP стимулируется гликолиз путём стимулирования PFK-1 .
Особая роль печени в поддержании постоянного уровня глюкозы в крови требует дополнительных регуляторных механизмов, координирующих образование и потребление глюкозы. Когда уровень глюкозы в крови понижается, гормон глюкагон сигнализирует печени, чтобы она образовывала и выделяла в кровь больше глюкозы и приостановила использование глюкозы для собственных нужд. Одним из источников глюкозы является гликоген, запасённый в печени. Другим источником является глюконеогенез, использующий в качестве исходных реагентов пируват, лактат , глицерол и некоторые аминокислоты. Когда уровень глюкозы в крови высок, другой гормон, инсулин , сигнализирует печени, чтобы она использовала глюкозу как «топливо» и предшественник для синтеза и запасания гликогена и триацилглицерола .
Быстрая гормональная регуляция гликолиза и глюконеогенеза опосредована фруктозо-2,6-бисфосфатом — аллостерическим регулятором ферментов PFK-1 и FBPазы-1. Когда фруктозо-2,6-бисфосфат связывается с особым аллостерическим сайтом на PFK-1, он увеличивает сродство фермента к его субстрату — фруктозо-6-фосфату и уменьшает его сродство к аллостерическим ингибиторам — ATP и цитрату. При физиологических концентрациях субстратов PFK-1 — АТФ и фруктозо-6-фосфата , а также других положительных или отрицательных эффекторов этого фермента (ADP, AMP, цитрат) PFK-1 находится в инактивированной форме в отсутствие фруктозо-2,6-бисфосфата . На FBPазу-1 фруктозо-2,6-бисфосфат имеет противоположный эффект: он снижает его сродство к субстратам и тем самым замедляет глюконеогенез .
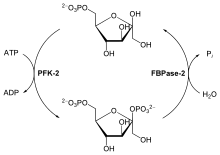
Клеточная концентрация аллостерического регулятора фруктозо-2,6-бисфосфата складывается из относительных скоростей его образования и разрушения. Он образуется при фосфорилировании фруктозо-6-фосфата ферментом фосфофруктокиназой-2 (PFK-2), а разрушается фруктозо-2,6-бисфосфатазой (FBPазой-2). PFK-2 и FBPаза-2 представляют собой две различные ферментные активности одного и того же бифункционального белка. Баланс этих активностей в печени, определяющих клеточный уровень фруктозо-2,6-бисфосфата, регулируется глюкагоном и инсулином .
Глюкагон стимулирует аденилатциклазу печени к синтезу 3',5'- cAMP из ATP. cAMP далее активирует cAMP-зависимую протеинкиназу, которая переносит фосфорильную группу с ATP на бифункциональный белок PFK-2/FBPазу-2. Фосфорилирование этого белка усиливает активность FBPазы-2 и подавляет активность PFK-2. Поэтому глюкагон снижает уровень фруктозо-2,6-бисфосфата в клетке, тем самым подавляя гликолиз и стимулируя глюконеогенез. Этим обусловлен эффект глюкагона на повышение уровня глюкозы в крови. Инсулин имеет противоположный эффект, так как он стимулирует активность фосфопротеинфосфатазы , катализирующей перенос фосфорильной группы с PFK-2/FBPазы-2, что активирует PFK-2, в результате чего уровень фруктозо-2,6-бисфосфата увеличивается, стимулируется гликолиз и подавляется глюконеогенез .

Действие другого регуляторного механизма также основано на регуляции уровня фруктозо-2,6-бисфосфата. В печени млекопитающих ксилулозо-5-фосфат, продукт пентозофосфатного пути, также задействован в увеличении темпов гликолиза после потребления богатой углеводами пищи. Уровень ксилулозо-5-фосфата в клетке увеличивается, когда глюкоза, поступающая в печень, превращается в глюкозо-6-фосфат и далее участвует как в гликолизе, так и пентозофосфатном пути. Ксилулозо-5-фосфат активирует фосфопротеинфосфатазу 2А ( PP2A ), который дефосфорилирует бифункциональный фермент PFK-2/FBPазу-2. Дефосфорилирование активирует PFK-2 и подавляет FBPазу-2, из-за чего повышается концентрация фруктозо-2,6-бисфосфата, что стимулирует гликолиз и подавляет глюконеогенез. Увеличение темпов гликолиза запускает образование ацетил-CoA, а увеличение темпов пентозофосфатного пути приводит к образованию NADPH. Ацетил-CoA и NADPH служат стартовыми реагентами для синтеза жирных кислот, так что при потреблении богатой углеводами пищи идёт интенсивный синтез жирных кислот. Ксилулозо-5-фосфат также увеличивает образование всех ферментов, необходимых для синтеза жирных кислот .

У позвоночных имеется по меньшей мере 3 изозима пируваткиназы, которые различаются местонахождением в тканях и ответом на воздействия модуляторов. Высокая концентрация ATP, ацетил-CoA, длинных жирных кислот (признаки достаточной энергетической обеспеченности) аллостерически подавляют все изозимы пируваткиназы. Пируваткиназа печени (L-форма), но не мышц (M-форма), также регулируется фосфорилированием. Когда низкий уровень глюкозы в крови приводит к выбросу глюкагона, cAMP-зависимая протеинкиназа фосфорилирует L-изозим пируваткиназы, инактивируя его. Это замедляет использование глюкозы в печени как источника энергии и направляет её на экспорт в мозг и другие органы. В мышцах эффект повышения концентрации cAMP строго противоположный. В ответ на воздействие адреналина cAMP активирует распад гликогена и гликолиз .
Большая часть вышеописанных регуляторных путей опосредована быстрыми, легко обратимыми процессами: аллостерическим эффектом, фосфорилированием фермента или связыванием с регуляторным белком. Однако существуют и механизмы регуляции, основанные на изменении количества молекул фермента в клетке за счёт изменений в балансе синтеза и разрушения фермента. Эти механизмы регулируются на уровне транскрипции соответствующего фермента .
Под влиянием инсулина находится транскрипция более чем 150 человеческих генов. В их числе гены, участвующие в гликолизе и его регуляции, а именно кодирующие гексокиназы II и IV, PFK-1, пируваткиназу, PFK-2/FBPазу-2 .
Одним из транскрипционных факторов , важных для метаболизма углеводов , является ChREBP ( англ. carbohydrate response element binding protein), экспрессируемый, главным образом, в печени, жировой ткани и почках. Он служит для координирования синтеза ферментов, необходимых для синтеза углеводов и жиров. В неактивной форме ChREBP фосфорилирован двумя фосфатами по и находится в цитозоле, будучи неспособным пройти в ядро. Когда фосфопротеинфосфатаза РР2А удаляет с него один фосфат, ChREBP проникает в ядро, где РР2А убирает с него второй фосфат. Активированный таким образом ChREBP связывается с белком-партнёром, . Комплекс ChREBP-Mlx теперь связывается с элементом ChoRE ( англ. carbohydrate response element) на ДНК в области его промотора и стимулирует его транскрипцию. РР2А аллостерически активируется ксилулозо-5-фосфатом. С помощью ChREBP регулируется синтез таких ферментов, как пируваткиназа, синтаза жирных кислот и . Другой транскрипционный фактор, функционирующий в печени — — регулирует образование пируваткиназы, гексокиназы IV, липопротеинлипазы, ацетил-CoA-карбоксилазы и синтазы жирных кислот. Синтез SREBP-1c стимулируется инсулином и подавляется глюкагоном .
В гликолизе может протекать дополнительная реакция, превращающая 1,3-бисфосфоглицерат в 2,3-бисфосфоглицерат ; эта реакция катализируется ферментом бисфосфоглицератмутазой . 2,3-Бисфосфоглицерат может возвращаться в гликолиз под воздействием фермента 2,3-бисфосфоглицератфосфатазы , которая превращает его в 3-фосфоглицерат . В большинстве тканей количество 2,3-бисфосфоглицерата невелико, но в эритроцитах его содержание значительно, поскольку там он функционирует как аллостерический регулятор гемоглобина . Он связывается с гемоглобином и понижает его сродство к кислороду, способствуя диссоциации последнего и его переходу в ткани .
Некоторые модификации гликолиза обнаружены у бактерий. В частности, когда окисление глицеральдегид-3-фосфата на стадии 6 глицеральдегид-3-фосфатдегидрогеназой ограничено низким содержанием фосфата в среде, у E. coli и некоторых других бактерий дигидроксиацетонфосфат окисляется до пирувата через три реакции, составляющие метилглиоксалевый шунт . В реакции 1 лиаза отщепляет фосфат с образованием метилглиоксаля . В реакции 2 метилглиоксаль присоединяет воду, превращаясь в лактат, вод воздействием . В реакции 3 лактат окисляется мембраносвязанной флавин -содержащей D-лактатооксидазой до пирувата. Если содержание фосфата в среде высоко, то метилглиоксалевый шунт не функционирует, так как лиаза ингибируется фосфатом .
Наконец, у анаэробных бактерий существуют дополнительные пути разложения углеводов. В частности, бактерии, предпочитающие пентозы в качестве субстрата, превращают пентозы и гексозы в ксилулозо-5-фосфат, который далее расщепляется .
Кроме того, у некоторых термофильных архей имеются только 2 из гликолитических ферментов — енолаза и пируваткиназа .
Гликолиз является универсальным, хотя и не единственным, путём катаболизма глюкозы и активно используется как про- , так и эукариотическими организмами . Все десять ферментов гликолиза водорастворимы и находятся в цитозоле . Некоторые ткани и клетки животных способны катаболизировать глюкозу исключительно с помощью гликолиза (например, нейроны мозга или клетки почечных канальцев ). В печени и жировой ткани физиологическая роль гликолиза несколько отличается от таковой в других тканях. Во время пищеварения в печени и жировой ткани гликолиз функционирует в основном как источник субстратов для синтеза жиров . Некоторые ткани растений специальным образом модифицированы для запасания крахмала (например, в клубне картофеля), а некоторые водные растения (например, жеруха ) получают большую часть энергии именно через гликолиз .
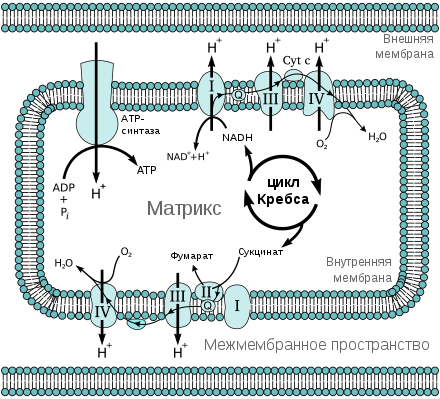
Как отмечалось выше, в аэробных условиях пируват после гликолиза образует ацетил-CoA и вовлекается в цикл Кребса. Две молекулы НАДH, образующиеся при гликолизе в цитозоле, в этих условиях вновь окисляются до НАД + , отдавая свои электроны в электроно-транспортную цепь (ЭТЦ), которая у эукариот находится в митохондриях . По ЭТЦ эти электроны переходят от одного переносчика к другому, пока не дойдут до конечного акцептора электронов — кислорода:
Перенос электронов от НАДH к O 2 в митохондриях обеспечивает энергию для синтеза АТФ путём окислительного фосфорилирования .
В анаэробных условиях пируват подвергается дальнейшим превращениям, обеспечивая регенерацию НАД + и других предшественников биосинтеза ( брожение ). При этом образуются продукты брожения, такие, как, например, лактат или этанол . В этих условиях гликолиз является единственным способом получения энергии для синтеза ATP из ADP и P i . У некоторых анаэробов, как и у аэробов, функционирует ЭТЦ, однако конечным акцептором электронов служит не кислород, а отличное от него окисленное органическое или неорганическое вещество .
Также на анаэробный тип метаболизма переходят некоторые органы и ткани в условиях гипоксии (нехватки кислорода), например, скелетные мышцы при активной работе. В анаэробных условиях в них пируват преобразуется в лактат, который транспортируется в другие ткани (например, печень, сердечную мышцу ) и там вновь превращается в пируват ( цикл Кори ) . Помимо этого, анаэробный распад глюкозы имеет место в эритроцитах, так как в них отсутствуют митохондрии .

Особое физиологическое значение гликолиз имеет в адипоцитах , где он поставляет метаболиты для липогенеза и направляет жирные кислоты вместо ненужного окисления на синтез триглицеридов , тем самым снижая окислительный стресс . В нейронах гипоталамуса гликолиз является важным регуляторным звеном контроля .

При накоплении лактата, образующегося при анаэробных условиях, в крови (например, при интенсивной и длительной работе) развивается лактатацидоз — обусловленное накоплением лактата понижение pH крови , что вызывает резкие нарушения в клеточном метаболизме. Так происходит при некоторых патологических состояниях, когда нарушается снабжение тканей кислородом: инфаркт миокарда , лёгочная эмболия , кровотечение . Лактатацидоз может быть обусловлен сахарным диабетом , когда аэробный гликолиз сменяется анаэробным . Поскольку инсулин ускоряет гликолиз, при диабете I типа (когда вырабатывается слишком малое количество инсулина) происходит замедление гликолиза . По этой причине препараты, стимулирующие гликолитические ферменты и ферменты, осуществляющие регуляцию гликолиза, могут стать важным средством лечения диабета .
При многих типах рака у животных и человека в опухолевых клетках потребление глюкозы и гликолиз ускоряются почти в 10 раз по сравнению с нормальной клеткой. Дело в том, что большинство опухолевых клеток живут в условиях гипоксии, так как на первых порах нет капиллярной сети, которая в необходимой мере снабжала бы их кислородом. По этой причине в энергетическом плане опухолевые клетки становятся целиком зависимыми от гликолиза, который энергетически гораздо менее эффективен, чем полное окисление глюкозы до углекислого газа и воды, и опухолевой клетке приходится потреблять гораздо больше глюкозы, чем нормальной. По-видимому, на ранних этапах трансформации нормальной клетки в опухолевую происходит переход на исключительно гликолитическое энергообеспечение и развивается устойчивость к низкому рН внеклеточной среды (снижение рН обусловлено накоплением лактата) .
Увеличение темпов гликолиза у опухолевых клеток достигается увеличением синтеза гликолитических ферментов и инсулиннезависимых мембранных переносчиков глюкозы ( GLUT1 и GLUT3 ). Индуцируемый гипоксией транскрипционный фактор ( HIF-1 ) на уровне мРНК усиливает образование по крайней мере восьми ферментов гликолиза, а также переносчиков глюкозы в условиях нехватки кислорода. Он также увеличивает образование пептидного гормона VEGF ( фактор роста эндотелия сосудов ), который стимулирует разрастание капиллярной сети навстречу опухоли .
Большая, по сравнению с нормальными тканями, зависимость опухолей от гликолиза даёт возможности для разработки противораковой терапии : ингибиторы гликолиза поражали и убивали бы опухолевые клетки, снижая их обеспечение АТР. В качестве химиотерапевтических агентов в будущем могут применяться три ингибитора гексокиназы: , лонидамин и . Препятствуя образованию глюкозо-6-фосфата, они блокируют не только гликолиз, но и пентозофосфатный путь , который также начинается с этого соединения. Без пентозофосфатов, образующихся в этом пути, клетка не может синтезировать ДНК - и РНК - нуклеотиды , а значит, расти и делиться. Другим препаратом, уже применяющимся на практике, стал иматиниб — блокатор тирозинкиназы , который предотвращает повышение образования гексокиназы, активируемое этой киназой .
Высокая скорость гликолиза в опухолевых клетках также имеет значение для диагностики раковых заболеваний. Относительная скорость потребления глюкозы тканью в некоторых случаях может помочь установить нахождение опухоли. При позитронно-эмиссионной томографии пациенту вводят безвредную, меченную изотопом фтора глюкозу ( фтордезоксиглюкозу ), которая превращается гексокиназой в 6-фосфодезоксиглюкозу и далее не подвергается метаболическим превращениям, накапливаясь в клетках. Метка обнаруживается специальными детекторами, располагающимися по всему телу, и таким образом определяется локализация опухоли .
Роль гликолиза и в брожении, и в дыхании имеет эволюционные основы. Предполагается, что древние прокариоты использовали гликолиз для получения АТФ задолго до того, как кислород накопился в земной атмосфере . Древнейшие известные ископаемые остатки бактерий имеют возраст 3,5 миллиардов лет, однако значительные количества кислорода в атмосфере стали накапливаться 2,7 миллиардов лет назад. Цианобактерии образовывали О 2 как побочный продукт при фотосинтезе. По этой причине, возможно, гликолиз был единственным источником ATP для древних прокариот. Тот факт, что в настоящее время гликолиз является наиболее широко распространённым метаболическим путём на Земле, подтверждает то, что он появился на очень ранних этапах истории жизни. О древности гликолиза свидетельствует и то, что все его ферменты локализованы в цитозоле и для протекания этого пути не требуются особые мембранные органеллы, которые появились приблизительно миллиард лет спустя после возникновения прокариотических клеток. Выше говорилось, что у некоторых термофильных архей из всех 10 гликолитических ферментов имеются только енолаза и пируваткиназа, поэтому может быть, что система ферментов гликолиза развилась из такой двухкомпонентной системы . Таким образом, гликолиз можно рассматривать как метаболическое «наследие» от ранних клеток, который и сейчас используется при брожении и как первый этап разрушения органических соединений при дыхании .

Гликолиз стал первым тщательно описанным метаболическим путём, и по сей день, возможно, остаётся наиболее изученным. После открытия спиртового брожения в экстрактах клеток дрожжей в 1897 году Эдуардом Бухнером и описания всего процесса гликолиза у дрожжей ( Отто Варбург и Ханс Эйлер-Хельпин ) и в мышечной ткани ( Густав Эмбден , Отто Мейергоф , Яков Парнас , считающиеся первооткрывателями гликолиза; в честь них гликолиз получил своё второе название) конкретный механизм реакций гликолиза находился в центре биохимических исследований. В ходе изучения гликолиза развивались методы выделения ферментов, были открыты коферменты, в частности, NAD, и была установлена их глобальная роль, была установлена важнейшая метаболическая роль ATP и других фосфорилированных соединений .

Понимание того, что именно фосфорилированные гексозы являются промежуточными соединениями гликолиза, пришло не сразу и по счастливой случайности. В 1906 году Артур Гарден и проверяли свою гипотезу о том, что ингибиторы протеолитических ферментов могут стабилизировать ферменты, сбраживающие глюкозу. Они добавили сыворотку крови , которая содержит ингибиторы протеолитических ферментов , в экстракт дрожжей и наблюдали ожидаемое ускорение метаболизма глюкозы. Однако в контрольном эксперименте, который должен был показать, что прокипячённая сыворотка не оказывала стимулирующего действия, выяснилось, что прокипячённая сыворотка стимулировала гликолиз. Тщательная проверка компонентов сыворотки показала, что стимулирование было обусловлено наличием в сыворотке неорганического фосфата . В дальнейшем Гарден и Янг установили, что глюкоза, добавленная в экстракт дрожжей, превращалась в гексозобисфосфат («эфир Гардена — Янга», известный как фруктозо-1,6-бисфосфат). Это было началом длинной череды открытий, показавших роль органических эфиров и фосфатных ангидридов в биохимии .