.jpg)







Ирбис
- 1 year ago
- 0
- 0

Млекопита́ющие ( лат. Mammalia ) — класс позвоночных животных , основной отличительной особенностью которых является вскармливание детёнышей молоком . Класс входит в кладу синапсид надкласса четвероногих и представляет собой самую прогрессивную группу позвоночных . По подсчётам биологов, известно более 6500 современных видов млекопитающих , среди которых и вид Человек разумный ( лат. Homo sapiens ); общее же число видов млекопитающих, включая вымершие, превышает 20 000 .
Млекопитающие распространены почти повсеместно . Произошли от высокоразвитых синапсид в конце триаса . К млекопитающим относят ныне живущих однопроходных , сумчатых и плацентарных , а также большое число вымерших групп. Современные виды, входящие в класс млекопитающих, объединяют в 26—29 отрядов .
Внешний вид млекопитающих весьма разнообразен, но в целом соответствует характерному и для других четвероногих плану строения . Анатомия и физиология млекопитающих характеризуется наличием тех же функциональных систем, что и у прочих четвероногих. Однако многие из этих систем достигли высокого уровня развития, так что данный класс среди позвоночных считают наиболее высокоорганизованным . Приспособления млекопитающих к жизни в различных средах обитания весьма разнообразны, их поведение отличается сложностью и многообразием . Они занимают доминирующие позиции среди наземной фауны (в водной среде они уступают лишь лучепёрым рыбам ). Млекопитающие играют большую роль в жизни и хозяйственной деятельности человека: они выступают как важные источники продуктов питания и производственного сырья , выполняют транспортные функции, служат тягловой силой, используются как лабораторные животные и домашние питомцы .
Изучением млекопитающих занимается наука териология ( маммалиология ) .
Латинское наименование Mammalia представляет собой производное от лат. mamma — « грудь , вымя » (дано в связи с наличием молочных желёз у всех млекопитающих) . Такой же смысл имеют фр. mammifères , исп. mamíferos , итал. mammiferi , порт. mamíferos , эспер. mamuloj ( англ. mammals — заимствование из латыни). Значение «вскармливающие молоком» имеют названия нем. Säugetiere , нидерл. zoogdieren , рус. млекопитающие ; значение «сосущие молоко» — названия польск. ssaki , укр. .
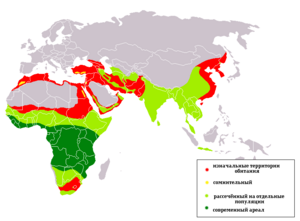
Млекопитающие распространены по всему миру и встречаются на всех континентах , во всех океанах и на большинстве существующих островов . Современный ареал однопроходных ограничивается Австралией , Тасманией и Новой Гвинеей , сумчатых — территорией Австралии, Океании и обоих американских континентов . Самое широкое распространение — у плацентарных , которые уже с конца мезозоя стали доминирующими наземными позвоночными на всех континентах, кроме Австралии и Антарктиды , преобладая по численности особей, биомассе и положению в пищевых цепях . В Австралии же до начала европейской колонизации обитали, не считая человека , лишь немногие виды плацентарных, а именно летучие мыши и мышиные . На отдалённых от других материков островах до прибытия человека существовала лишь бедная видами фауна млекопитающих: на многих из них, в том числе и в Новой Зеландии , её представляли только несколько видов летучих мышей .
Млекопитающие населяют почти все биотопы Земли и встречаются как в пустынях и тропических лесах , так и в высокогорье и в полярных регионах. К немногим регионам и биотопам, в которых, за исключением редкого пребывания людей, отсутствуют млекопитающие, относятся глубоководный океан и Антарктический континент , хотя у его побережья встречаются тюлени и киты . Однако в прошлом, когда Антарктида ещё не была сплошь накрыта ледяным щитом , млекопитающие населяли и этот материк: об этом свидетельствуют относящиеся к эоцену ископаемые остатки гондванатериев , сумчатых ( , микробиотерии ) и плацентарных ( неполнозубые , литоптерны , астрапотерии ) . Используя технические средства , в XX веке человек проник в места, недоступные для других млекопитающих: достиг Южного полюса , поднялся на вершины Джомолунгмы и других высочайших гор , спустился на дно Марианской впадины , приступил к освоению космического пространства . Впрочем, в некоторых из этих предприятий человека принимали участие собаки : Р. Амундсен добрался до Южного полюса на собачьих упряжках, а Лайка совершила свой за 3,5 года до Ю. А. Гагарина .
У многих млекопитающих ареал невелик, поскольку широкому их расселению препятствуют привязанность к условиям среды (температурный режим, пищевые ресурсы, почвенно-грунтовые и орографические условия) и наличие естественных препятствий — например, морских пространств . Так, гребнепалый тушканчик , обитающий в Кызылкуме и Восточных Каракумах , живёт только в сыпучих песках, поросших кустами ; ареал лемурообразных ограничен тропическими лесами Мадагаскара и соседних островов . Учёт специфики видового состава млекопитающих играет важную роль при зоогеографическом районировании Земли . Наибольшее количество эндемичных видов отмечено в следующих странах: Индонезия (201 эндемик из 436 видов местной териофауны), Австралия (198 из 252), Мексика (140 из 450), США (101 из 428), Филиппины (97 из 153), Бразилия (96 из 394) .
Кроме вскармливания потомства молоком , для большинства млекопитающих характерен целый ряд признаков, отличающих их, как правило, от других позвоночных :


Согласно современным представлениям, млекопитающие произошли от более примитивных синапсид из группы цинодонтов , выделившись в конце триасового периода . Наиболее продвинутые немаммальные цинодонты уже сильно напоминали млекопитающих — как, например, изображённый на рисунке Oligokyphus из семейства Tritylodontidae с его развитым шёрстным покровом, живший в позднем триасе и ранней юре . Тогда же имела место и начальная дивергенция млекопитающих: в отложениях позднего триаса найдены ископаемые остатки аделобазилевса , морганукодонтов , и харамиид . Последних обычно рассматривают как ранних представителей подкласса (или инфракласса) аллотериев , куда включают также многобугорчатых — наиболее разнообразный и многочисленный из мезозойских отрядов млекопитающих, просуществовавший свыше 100 млн лет ; что касается морганукодонтов, то они по своему облику и строению чрезвычайно близки к предполагаемому предку всех более поздних млекопитающих .
В верхнем триасе выделились и другие основные линии млекопитающих, известные остатки которых относятся к более позднему времени: линия, включающая однопроходных ; линия триконодонтов (юра — мел ) ; наконец, та линия, к которой принадлежат сумчатые и плацентарные , отделившиеся друг от друга в юрском периоде .

Современные виды млекопитающих образуют хорошо очерченную группу. Эта группа и была описана в 1758 году как класс Mammalia К. Линнеем в 10-м издании его « Системы природы » . В данном издании Линней упоминал всего 184 вида млекопитающих ; в настоящее же время современные виды млекопитающих объединяют в 26—29 отрядов , 153 семейства и 1229 родов .
Позднее в состав класса млекопитающих были включены многочисленные вымершие группы; правомерность включения некоторых из них (например, , морганукодона , синоконодона и некоторых других открытых в XIX—XX веках мезозойских животных) в данный класс вызывает сомнения у ряда палеонтологов .
Традиционная классификация на уровне инфраклассов и выше, оформившаяся к концу XIX века, выглядит следующим образом :
Класс Mammalia Linnaeus, 1758

Названия, заключённые в скобки, употребляют либо как альтернативные наименования соответствующих инфраклассов , либо как названия их тотальных групп .
Трёхчастное деление класса млекопитающих на прото- , мета- и эутериев было предложено ещё французским зоологом А. Блэнвилем (1839), который пользовался другими наименованиями (соответственно: «птицеутробные», «двуутробные», «одноутробные») .
Неоднократно предлагались более современные классификации млекопитающих, позволяющие, в частности, адекватно отразить взаимоотношения между многочисленными вымершими группами, но общепринятыми ни одна из них не стала .

На русский язык название таксона Theria , крупнейшего подразделения млекопитающих, обычно переводится как « звери » (в данном значении оно употребляется параллельно со словом «терии»; см. Звери ). Иногда же слово «звери» используют как синоним слова «млекопитающие» . Ранг данного таксона в различных классификациях различен: либо это подкласс млекопитающих, противопоставляемый подклассу Prototheria ( первозвери ) , либо — после исключения из него ряда вымерших групп — таксон существенно низшего ранга (в системе Маккенны — Белл он имеет ранг «надкогорты» ).
Филогения млекопитающих по молекулярному анализу:
|
Современные отряды
(Mammalia)
|
|||||||||
|---|---|---|---|---|---|---|---|---|---|
|
|||||||||
|
Prototheria
(Первозвери) |
|||||||||
|
Metatheria
(Метатерии) |
|
||||||||
|
Eutheria
(Эутерии) Placentalia (Плацентарные) |
|
||||||||
По размерам и внешнему облику млекопитающие очень разнообразны. Хотя у их подавляющего большинства тело, как и у других амниот , чётко расчленено на голову , шею , туловище , две пары конечностей и хвост , форма и соотношение этих частей тела у разных видов варьируют, отражая приспособления к среде обитания и преобладающему характеру движений . Хорошо известны как непропорционально длинная шея жирафа , позволяющая ему срывать листья и молодые побеги с крон деревьев , так и полное отсутствие шейного перехвата у китообразных . Сильно варьирует и относительная длина хвоста: если у длиннохвостого ящера ( Manis tetradactyla ) и длинноухого тушканчика ( Euchoreutes naso ) он в два раза длиннее туловища, то у даманов , тонкого лори и всех гоминоидов хвост полностью отсутствует . Форма головы, в частности соотношение его лицевой и мозговой частей, зависит от принимаемой пищи. Так, более коротка лицевая часть у хищных и грызунов , которым требуется большая сила сжатия челюстей и, соответственно, меньшая длина рычага сопротивления. Напротив, челюсти относительно удлиняются у роющих ( свиньи ), растительноядных ( жвачные ), требующих увеличения поверхности перетирания пищи, и захватывающих большие объёмы зверей ( усатые киты ) .
Наиболее сильно по внешнему облику от других млекопитающих уклонились рукокрылые , у которых передние конечности видоизменились в , а также сирены и китообразные , передние конечности которых после перехода к водному образу жизни превратились в плавники , а задние исчезли . Поэтому в народной систематике и ранних научных зоологических классификациях рукокрылые и киты не включались в состав группы «звери» . Только в 1693 г. Дж. Рэй со всей определённостью включил рукокрылых и китов в состав той группировки, которую ныне называют «млекопитающими» .
Сильно различаются млекопитающие также по массе и размерам тела. Абсолютные рекордсмены:

|

|

|

|
_crop.jpg)
|


Кожа млекопитающих отличается значительной толщиной и сильным развитием кожных желёз и роговых образований. Как и у других позвоночных , она состоит из двух слоёв: верхнего эктодермального ( эпидермис ) и нижнего мезодермального ( дерма ) .
Для млекопитающих весьма характерны роговые образования эпидермального происхождения: волосы, когти , ногти , копыта , рога и чешуйки . При этом волосы — образование, присущее исключительно млекопитающим и, по-видимому, наиболее продвинутым терапсидам ; оно не гомологично каким-либо производным кожи других амниот. Главная функция волос — теплоизоляция, хотя толчком для появления волосяного покрова послужила, скорее всего, осязательная функция . Помимо термоизолирующей и осязательной функций, волосы защищают кожу от повреждений и паразитов , улучшают аэро- и гидродинамические свойства тела, обеспечивают видоспецифичность окраски. Значительная редукция или полное отсутствие волос у ряда млекопитающих ( слоны , сирены , некоторые носороги , гиппопотамы , китообразные , люди ) носят вторичный характер .
У большинства млекопитающих концевые фаланги пальцев защищены когтями , видоизменениями которых являются плоские ногти , имеющиеся у многих приматов , и копыта , характерные для копытных (кроме уже упомянутых китообразных ). Многие млекопитающие имеют на хвосте или конечностях роговые чешуйки , а у панголинов и броненосцев крупная роговая чешуя , вполне гомологичная имеющейся у пресмыкающихся , покрывает всё тело . Для ряда млекопитающих ( арсинойтерий , носороги , вилороговые , полорогие ) характерно наличие такого «оборудования», как рога ; обычно это — массивные разрастания ороговевшего эпителия , используемые для защиты и нападения . Рога представителей вымершего отряда эмбритопод были образованы не кератином , а костной тканью

Экзокринную систему образуют железы внешней секреции , выделяющие вырабатываемый ими секрет через выводные протоки на поверхность тела или слизистых оболочек внутренних органов (например, функции таких желёз выполняют слюнные железы , печень и поджелудочная железа , секреты которых, попадая в пищеварительный тракт, играют важную роль в ходе пищеварения ) .
Характерной особенностью млекопитающих является сильное развитие кожных желёз , которую они унаследовали через своих синапсидных предков от земноводных . К их числу относятся сальные и потовые железы , развитие которых было связано с появлением волосяного покрова . Видоизменениями потовых желёз по своему происхождению являются млечные железы , которые в качестве секрета выделяют молоко — основную пищу детёнышей млекопитающих. У представителей сумчатых и плацентарных протоки млечных желёз открываются на сосках (млечные железы однопроходных сосков не имеют). Число сосков может варьировать у разных видов от 2 до 24, причём у многих копытных соски сливаются в единое вымя .

Подобно другим позвоночным , млекопитающие имеют внутренний скелет (у броненосцев есть и внешний скелет в виде панциря — как дополнение к внутреннему). Скелет включает череп , позвоночник , грудную клетку , пояса верхних и нижних конечностей и сами конечности .
Череп млекопитающих имеет относительно крупную мозговую коробку, уже у позднетриасового морганукодона она в 3—4 раза крупнее, чем у терапсид того же размера . Из-за срастания некоторых костей между собой число отдельных костей в черепе меньше, чем у других позвоночных . У млекопитающих отсутствуют , и теменное отверстие . На поверхности лобных и теменных костей от вершин заглазничных отростков тянутся два черепных гребня , которые иногда сливатся в единый стреловидный гребень, особенно развитый у хищных . Для черепной коробки млекопитающих характерно наличие скуловой дуги , образованной скуловой костью и скуловым отростком чешуйчатой кости (для высших приматов правильнее говорить о чешуйчатой части височной кости : последняя формируется при взрослении особей путём срастания чешуйчатой, барабанной и костей ), и причленение черепа к первому шейному позвонку ( атланту ) при помощи двух мыщелков , образованных , реже дополнительно . Верхние челюсти образованы верхнечелюстными и межчелюстными костями; только на них расположены верхние зубы. Нёбные отростки верхнечелюстных костей вместе с нёбными костями образуют также характерное для млекопитающих вторичное костное нёбо , а нижняя челюсть у них представлена лишь парными , образующими сустав напрямую с чешуйчатой костью скуловой дуги. Переднее носовое отверстие непарное . Только млекопитающим свойственна расположенная в дне черепа парная барабанная кость; имеющиеся в черепе рептилий и немаммальных синапсид сочленовная и квадратная кости становятся у млекопитающих элементами среднего уха — молоточком и наковальней .
Характерные черты в строении позвоночного столба млекопитающих: плоские поверхности позвонков ( платицельные позвонки ), между которыми располагаются хрящевые диски; условное расчленение позвоночника на пять отделов — шейный, грудной, поясничный, крестцовый и хвостовой ( крестца нет лишь у современных китообразных ); постоянное число шейных позвонков — 7 (как исключение, оно равно 6 у ламантина , а у некоторых ленивцев число шейных позвонков варьирует: у ленивца Гоффмана — от 5 до 6, у трёхпалых ленивцев — от 8 до 10 ) . Грудной отдел позвоночника у млекопитающих чаще всего состоит из 12—15, поясничный — из 2—9, крестцовый — из 1—9 позвонков; сильнее всего варьирует число хвостовых позвонков: от 3 у гиббоновых до 49 у длиннохвостого ящера . К передним грудным позвонкам причленяются рёбра , соединённые с грудиной и образующие вместе с ней грудную клетку .

Пояс передних конечностей млекопитающих обычно состоит из двух лопаток и двух ключиц ; последние, впрочем, нередко редуцированы или вовсе отсутствуют. У однопроходных имеется ещё коракоид , который у остальных млекопитающих рудиментарен и прирастает к лопатке . В типичном случае каждая из причленённых к этому поясу передних конечностей состоит из плеча , предплечья и кисти ; последняя включают запястье, пясть и пальцы, которые обычно состоят из двух-трёх фаланг (однако у китообразных число фаланг увеличено и достигает 13—14). Пояс задних конечностей у млекопитающих, как правило, состоит из парных лобковых , седалищных и подвздошных костей; в состав каждой из задних конечностей входят бедро , голень и стопа (включающая предплюсну , плюсну и кости пальцев ). В зависимости от образа жизни конечности млекопитающих претерпевают различные преобразования: у наземных форм значительно удлиняются бедро и голень, а у водных форм они укорочены, но удлинены фаланги пальцев .

Высокодифференцированная мышечная система млекопитающих обеспечивает выполнение разнообразных движений и имеет в разных отрядах и семействах существенные особенности в зависимости от характерных способов передвижения. Среди скелетных мышц , наибольшее развитие получили мышцы конечностей ; перемещение конечностей под тело (у рептилий и ранних синапсид они располагались по бокам тела) увеличило величину шага и эффективность работы мышц. Характерным анатомическим признаком млекопитающих является также наличие куполообразной мышцы — диафрагмы , которая разделяет грудную и брюшную полости; её возникновение позволило резко интенсифицировать вентиляцию лёгких .
У млекопитающих хорошо развита жевательная мускулатура , обеспечивающая захват и механическую обработку пищи, и подкожная мускулатура , участвующая в терморегуляции (управляет движением волосяного покрова) и общении животных (отвечает за мимику , особенно развитую у хищных и приматов ). Все перечисленные виды мышц относятся к группе поперечно-полосатых ; мышцы же внутренних органов являются, как правило, гладкими . Особая сердечная мышечная ткань имеется в сердце (подробнее см. раздел « »).
Нервная ткань млекопитающих, как и у других позвоночных , эктодермального происхождения и состоит из нервных клеток ( нейронов ) и вспомогательных клеток — нейроглии . Нейроны образуют многочисленные контакты ( синапсы ) друг с другом и осуществляют генерирование и распространение нервных импульсов; таким образом, нейрон — структурная и функциональная единица нервной системы . Саму же нервную ткань подразделяют на серое вещество , образованное телами нейронов и немиелинизированными нервными волокнами , и белое вещество , образованное миелинизированными нервными волокнами . Нервный импульс распространяется по нервным волокнам и передаётся от нейрона к нейрону посредством особого рода межклеточных контактов — синапсов . Почти у всех млекопитающих синаптическая передача осуществляется с помощью химических соединений самой разной природы — нейромедиаторов , а не электрических сигналов .
Как и у других позвоночных, нервную систему млекопитающих по морфологическим признакам делят на центральную , включающую головной и спинной мозг , и периферическую , которая слагается отходящими от головного и спинного мозга нервами .
В структурно-функциональном плане нервную систему подразделяют на соматическую , иннервирующую скелетную мускулатуру и органы чувств , и вегетативную , иннервирующую внутренние органы , кровеносные и лимфатические сосуды ; резкого разграничения между данными системами не наблюдается .

Относительные размеры головного мозга у млекопитающих очень велики: по массе он в 3—15 раз превосходит спинной мозг (у рептилий их масса примерно одинакова) . Головной мозг млекопитающих включает пять отделов: передний (или конечный ), промежуточный , средний , продолговатый мозг и мозжечок , из них особенно развиты первый и последний . Разросшаяся кора полушарий образовала вторичный мозговой свод ( неопаллиум ), накрывший собой весь мозг .
Для спинного мозга млекопитающих характерен своеобразный характер залегания серого вещества , при котором оно образует в поперечном сечении фигуру «бабочки» с боковыми выступами — «рогами». У млекопитающих имеется особый пирамидный путь , идущий от двигательной зоны коры больших полушарий прямо к эфферентным нейронам продолговатого и спинного мозга .
Периферическую нервную систему млекопитающих образуют черепные и спинномозговые нервы , отходящие соответственно от головного и спинного мозга, а также их ответвления (всего в организме млекопитающего насчитывают свыше 200 нервов вместе с их ответвлениями). У млекопитающих, как и у других амниот , имеется 12 пар черепных нервов; число спинномозговых нервов значительно больше (у человека — 31 пара) .
Большинство (11 пар из 12) черепных и часть спинномозговых нервов функционально относят к соматической нервной системе , эти нервы иннервируют кожу , суставы , скелетные мышцы и органы чувств . Другую часть спинномозговых нервов, а также в значительной степени блуждающий нерв функционально относят к вегетативной нервной системе . Морфологически и функционально разделяют симпатическую и парасимпатическую вегетативную нервную систему. В случае первой из них центры локализованы только в спинном мозге . Для парасимпатической системы ситуация обратная: ганглии лежат недалеко от иннервируемых органов или даже в их стенках; центры расположены в крестцовом отделе спинного мозга и в головном мозге . При этом если у рыб симпатическая и парасимпатическая системы делят органы между собой, то у четвероногих (и, в частности, у млекопитающих) их области иннервации накладываются друг на друга вплоть до полного взаимного перекрывания .

Органы зрения у млекопитающих развиты, как правило, достаточно хорошо, хотя и имеют в их жизни меньшее значение, чем у птиц . Внутрь глаза отражённый от объектов свет проникает через зрачок . Хрусталик фокусирует лучи света на сетчатке глаза, а окружающие хрусталик мышцы заставляют его изменять форму, обеспечивая аккомодацию глаза . В отличие от пресмыкающихся и птиц, у млекопитающих при аккомодации расстояние между хрусталиком и сетчаткой не изменяется, а у мелких грызунов ( полёвки , мыши ) из-за незначительности обзора способность к аккомодации практически утрачена . Восприятие света обеспечивают находящиеся на сетчатке фоторецепторы : палочки отвечают только за восприятие света, а колбочки обеспечивают цветоразличение . Впрочем, цветовое зрение у млекопитающих развито слабее, чем у птиц, причём трёхкомпонентный (трихроматический) характер оно носит только у высших приматов , а у подавляющего большинства млекопитающих зрение — дихроматическое .

Органы слуха у подавляющего большинства видов млекопитающих хорошо развиты и при этом содержат вестибулярный аппарат . В состав каждого органа слуха входят внутреннее , среднее и наружное ухо (включающее наружный слуховой проход и ушную раковину ). Если внутреннее и среднее ухо имеются и у других четвероногих , то наружное ухо — новоприобретение млекопитающих, облегчающее улавливание звуковых волн . Ушные раковины обычно подвижны и способствуют достаточной точной (до 6—8°) локализации источников звука (впрочем, у многих водных и роющих видов ушная раковина вторично исчезает), а наружный слуховой проход защищает от внешних воздействий барабанную перепонку , отделяющую полость среднего уха от слухового прохода и колеблющуюся под воздействием прошедшей к ней по слуховому проходу звуковой волны. Специфично для млекопитающих и наличие в полости среднего уха трёх слуховых косточек ( молоточка , наковальни и стремечка ) . Назначение этих косточек — усилить колебания перепонки и передать их мембране овального окна внутреннего уха. Последнее образовано тремя полукружными каналами , улиткой и двумя отолитовыми органами — и ; при этом в улитке располагаются рецепторы , чувствительные к звуку, а полукружные каналы, мешочек и маточка входят в состав вестибулярного аппарата . Основу работы и слуховой, и вестибулярной сенсорных систем составляют механорецепторы , которые реагируют на перетекания и колебания заполняющей внутреннее ухо жидкости, вызванные звуковыми волнами или изменением положения тела .
Органы обоняния у млекопитающих развиты сильнее, чем у других наземных позвоночных. Способность к различению запахов млекопитающие используют для ориентирования в пространстве, при поисках пищи, в рамках межвидовых и внутривидовых контактов. По степени развития обонятельной функции млекопитающих подразделяют на две группы: макросматов с исключительно острым обонянием (большинство млекопитающих) и микросматов с умеренно развитым обонянием ( приматы , китообразные , ластоногие ) . Ко второй группе относится человек , в носу которого имеется около 5 млн обонятельных клеток, к первой— собака , у которой их примерно в 50 раз больше . В целом степень развитости обоняния коррелирует с числом генов , кодирующих различные типы обонятельных белков-рецепторов: у макросматов их обычно более 1000, у многих приматов — около 500, у человека — всего 387, у утконоса — 262 . По-видимому, хуже всего развито обоняние у китообразных, имеющих наиболее высокий процент псевдогенов обонятельных рецепторов . Основу функционирования обонятельной системы составляют обонятельные клетки — биполярные нейроны , генерирующие потенциал действия в ответ на связывание с особыми рецепторами на мембране молекул пахучего вещества .
Органы вкуса млекопитающих представлены вкусовыми почками , которые расположены на слизистых оболочках языка и твёрдого нёба и содержат рецепторы вкуса (последние, как и рецепторы запахов, относятся к группе хеморецепторов ). Традиционно считалось, что система восприятия вкусовых ощущений у млекопитающих — четырёхкомпонентная, причём первичными вкусами являются сладкий , солёный , кислый и горький . Сравнительно недавно выявлен пятый тип рецепторов вкуса, отвечающий за восприятие « мясного » вкуса .
У млекопитающих хорошо развита соматовисцеральная кожная чувствительность , к которой относят все типы сенсорной информации, получаемые от рецепторов , локализованных в теле (а не голове) животного. Среди таких рецепторов выделяют проприорецепторы (рецепторы растяжения мышц и сухожилий), ноцицепторы (болевые рецепторы) и поверхностные кожные рецепторы ( механорецепторы , терморецепторы ) . Определённую осязательную функцию выполняют также волоски , а также вибриссы .

Дыхательная система служит для газообмена организма с окружающей средой, обеспечивая поступление кислорода и выведение углекислого газа . У млекопитающих она представлена органами дыхания ( лёгкими ) и дыхательными путями : верхними и нижними. В систему верхних дыхательных путей входят полость носа , носоглотка и ротоглотка , а частично — и ротовая полость , поскольку она тоже может участвовать в процессе дыхания. Система нижних дыхательных путей состоит из гортани (иногда её относят к верхним дыхательным путям), трахеи и бронхов .
Строение гортани у млекопитающих усложнено: для неё характерно наличие щитовидного хряща и надгортанника , не свойственных другим позвоночным, а также голосовых связок — парных складок слизистой оболочки, вибрация которых позволяет издавать различные звуки .
У млекопитающих хорошо развиты трахея и бронхи (внелёгочные и внутрилёгочные). В ходе эмбрионального развития бронхи млекопитающих ветвятся, образуя сложное бронхиальное дерево . В процессе многократного разветвления бронхи переходят в терминальные бронхиолы , а те — в ещё более мелкие дыхательные бронхиолы . Последние лежат уже непосредственно в лёгких и переходят в , заканчивающиеся . На стенках альвеолярных ходов и альвеолярных мешочков располагаются альвеолы — тонкостенные пузырьки, густо оплетённые капиллярами ; через них и происходит газообмен. Число альвеол огромно: у хищных с их активным образом жизни — 300—350 млн, у малоподвижных ленивцев — около 6 млн.
Попеременное изменение объёма лёгких обеспечивают не только движения грудной клетки, но и движения диафрагмы , наличие которой — характерный анатомический признак млекопитающих . Дыханием управляет расположенный в продолговатом мозге дыхательный центр . Повышение в крови (а значит, и в спинномозговой жидкости) концентрации углекислого газа , сопровождающееся образованием угольной кислоты H 2 CO 3 , влечёт возрастание концентрации ионов H + ; они раздражают нервные клетки, которые и дают сигнал межрёберным мышцам усилить дыхательные движения .

Кровеносная система , призванная обеспечивать снабжение тканей тела кислородом и питательными веществами и освобождение их от продуктов распада, достигает у млекопитающих большого совершенства. Центральный орган кровообращения — четырёхкамерное сердце , ритмичные сокращения мускулатуры которого заставляют кровь циркулировать по кровеносным сосудам . Оно состоит из правого и левого желудочков , а также правого и левого предсердий . Предсердия собирают поступающую к сердцу по венам кровь и направляют её в желудочки, а те, сокращаясь, выбрасывают кровь в артерии . Камеры сердца сообщаются между собой и с магистральными сосудами — аортой , лёгочным стволом — при помощи отверстий, снабжённых клапанами . Сердце заключено в околосердечную сумку — перикард . Мышечная ткань сердца ( миокард ) имеет особое строение и представляет собой функциональный синцитий . В ней, помимо кардиомиоцитов , способных к сокращению , имеются клетки, не способные сокращаться, но способные к генерации электрических импульсов и проведению их к сократительным клеткам. Они образуют проводящую систему сердца .
Правая сторона сердца обеспечивает малый круг кровообращения ( лёгочный ): из правого желудочка венозная кровь поступает по лёгочным артериям в лёгкие, откуда по лёгочным венам обогащённая кислородом кровь следует в левое предсердие. Левая сторона сердца обеспечивает работу большого круга кровообращения : от левого желудочка отходит единственная дуга аорты — левая (этим млекопитающие отличаются от крокодилов с двумя дугами аорты и от птиц — с единственной, но правой ), через ответвления которой артериальная кровь разносится по всему телу. Затем кровь, отдав тканям организма кислород и питательные вещества и забрав углекислоту и продукты обмена, через венозную систему возвращается к сердцу — к правому предсердию, в которое впадают передняя (собирающая кровь от головы и передних конечностей) и задняя (от задней части тела) полые вены .
Кровь и связанная с ней лимфа — жидкие соединительные ткани организма млекопитающего . Кровь состоит из плазмы и форменных элементов : тромбоцитов, лейкоцитов и эритроцитов (все эти форменные элементы образуются у млекопитающих в костном мозге). Тромбоциты (у млекопитающих они, в отличие от других позвоночных, всегда представляют собой безъядерные кровяные пластинки ) играют определяющую роль в процессе свёртывания крови , а также участвуют в метаболизме серотонина . Лейкоциты ( белые кровяные тельца ) — основа иммунной системы организма. Эритроциты ( красные кровяные тельца ) содержат железосодержащий белок гемоглобин , обеспечивающий перенос кислорода от лёгких к тканям организма .
Особенности эритроцитов млекопитающих — их малый размер, двояковогнутая форма (у мозоленогих — овальная ) и отсутствие ядер . Двояковогнутая форма эритроцитов увеличивает площадь поверхности для диффузии кислорода внутрь эритроцита, а благодаря отсутствию ядра в эритроците помещается больше гемоглобина, связывающего кислород. У эритроцитов также отсутствуют митохондрии , и они синтезируют АТФ исключительно за счёт анаэробного дыхания . Если бы эритроциты потребляли кислород в ходе аэробного дыхания , они бы не были такими эффективными его переносчиками .
Лимфатическая система выступает как дополнение к кровеносной системе и служит посредницей в обмене веществ между кровью и тканями. Лимфа по происхождению — межтканевая жидкость, образованная просочившейся через стенки капилляров плазмой крови и протекающая по лимфатическим сосудам . Форменными элементами лимфы являются лимфоциты , причём имеет место рециркуляция лимфоцитов из крови в лимфу и из лимфы в кровь . В лимфатической системе млекопитающих отсутствуют « лимфатические сердца » — пульсирующие участки сосудов, имеющиеся у земноводных и пресмыкающихся, поскольку при характерном для млекопитающих подвижном образе жизни движение лимфы по сосудам обеспечивается сокращениями скелетных мышц .
Иммунная система позволяет организму противостоять бактериям , вирусам , грибам , паразитам , чужеродным макромолекулам , а также избавляет его от собственных модифицированных клеток (например, опухолевых ). Каждый организм имеет неспецифический (врождённый) иммунитет , который дополняется специфическим (приобретённым) иммунитетом . И неспецифический, и специфический иммунитет имеет несколько уровней защиты, однако неспецифические иммунные реакции протекают быстрее специфических .
Неспецифические защитные механизмы являются эволюционно более древними, чем специфические. Их активация и действие не зависят от патогена . Их также называют неклональными механизмами, потому что при неспецифических иммунных реакциях, в отличие от специфических, не происходит увеличения численности особой группы клеток, ориентированных на защиту от конкретного антигена. К неспецифическим механизмам можно отнести кислую среду кожи, препятствующую развитию микроорганизмов , непроницаемость эпидермиса , систему комплемента плазмы крови , антимикробные ферментные системы, а также неспецифические медиаторы, например, интерфероны и интерлейкины . На клеточном уровне неспецифическую защиту осуществляют гранулоциты , система моноцитов и макрофагов , а также натуральные киллеры . Работа последних основывается как на специфических, так и неспецифических механизмах. Важным неспецифическим защитным механизмом является также воспалительный процесс .
Специфический иммунный ответ направлен на защиту от конкретного антигена . Он включает в себя клеточные и гуморальные механизмы. Системный специфический иммунный ответ активируется миграцией антигенпрезентирующих клеток в лимфоидные органы, которые запускают размножение специфических групп защитных клеток. Среди них выделяют T- и B-лимфоциты . Эти клеточные системы обеспечивают крайне специфичные реакции на конкретный антиген и, как упоминалось выше, подвергаются клональной экспансии. Кроме того, специфические иммунные механизмы включают также механизмы , в которых запечатлевается информация о данном антигене, и при новом столкновении с этим антигеном она обеспечивает более быстрый и эффективный иммунный ответ .
Все клетки крови образуются из плюрипотентных стволовых клеток костного мозга . Однако в определённый момент эмбрионального развития часть из них мигрирует в тимус (вилочковую железу) или остаются в костном мозге и начинают созревание в T- и B-лимфоциты соответственно. В связи с этим тимус и костный мозг называют первичными лимфоидными органами . В них продолжается клеточная пролиферация и дифференциация . В процессе созревания T-лимфоциты контактируют со специализированными эпителиальными клетками, дендритными клетками и макрофагами, что даёт возможности для отбора T-клеток, нужных иммунной системе именно в этот момент . Зрелые T- и B-лимфоциты покидают места своего созревания и мигрируют во вторичные лимфоидные органы : селезёнку , лимфатические узлы , а также ( англ. mucosa-associated lymphoid tissue, MALT ). Она представляет собой скопления лимфоидных клеток, расположенные под слизистой желудочно-кишечного тракта, дыхательных путей, мочевыводящих путей, слёзных желёз . Кроме того, по всему телу беспорядочно разбросаны лимфатические клетки, иногда организованные в крупные скопления, например, миндалины или пейеровы бляшки .

Млекопитающие относятся к гомойотермным (то есть теплокровным) животным, обеспечивающим определённый уровень температуры тела в основном за счёт внутренних физиолого-биохимических процессов (исключение составляет пойкилотермный грызун голый землекоп ). При этом у них хорошо развита химическая терморегуляция : интенсивность окислительных процессов (основные из них — переваривание пищи и мышечная работа), обеспечивающих выделение тепла, регулируется рефлекторно — под воздействием сигналов теплового центра промежуточного мозга .
От излишней потери тепла млекопитающих предохраняет шёрстный покров, а у водных форм — подкожный слой жира . Излишки тепла отводятся потоотделением. Значительную роль в терморегуляции играет развитая система кожных кровеносных сосудов, диаметр просветов которых регулируется нервно-рефлекторным путём . Механизм теплообразования за счёт активации клеточного метаболизма называется несократительным термогенезом . К таким механизм относят, в частности, липолиз , особенно бурой жировой ткани . Кроме жира, в качестве внутренних энергетических запасов организма могут выступать и углеводы . Если же тепло образуется в процессе сокращения скелетных мышц, то говорят о сократительном термогенезе .

Для пищеварительной системы млекопитающих, в которую входят пищеварительный тракт (с 4 отделами: рото-глоточной полостью , пищеводом , желудком , кишечником ) и пищеварительные железы , характерны общее удлинение пищеварительного тракта при бо́льшей его дифференцировке по сравнению с другими группами позвоночных , которой сопутствует значительное развитие пищеварительных желёз . Высокой эффективности использования пищи способствует максимальное среди позвоночных развитие симбиотического пищеварения, при котором эндосимбионты ( археи - метаногены , бактерии , грибы , простейшие ) участвуют в процессе расщепления и гидролиза богатых клетчаткой растительных кормов .

Ротовое отверстие у млекопитающих ограничено присущими только им мягкими подвижными губами ; такие губы, впрочем, у однопроходных отсутствуют, а у китообразных губы слабо развиты и лишены подвижности . Зубы есть у большинства млекопитающих, кроме современных однопроходных , усатых китов , панголинов и муравьедов . Они сидят в ячейках (альвеолах) челюстных костей и являются гетеродонтными : дифференцированы на резцы , клыки , предкоренные и истинные коренные . Число зубов, их форма и размеры у разных млекопитающих различаются. Так, для грызунов характерны сильно развитая единственная пара резцов, служащая для срезания или обгрызания растительности, и отсутствие клыков, а у хищных клыки, напротив, сильно развиты и приспособлены для захвата и умерщвления добычи .
После попадания в ротовую полость пища пережёвывается зубами; при этом она смачивается слюной , поступающей по протокам из слюнных желёз, что облегчает её проглатывание и продвижение по пищеводу. Под влиянием слюны сложные углеводы ( крахмал , сахар ), содержащиеся в пище, распадаются на мономеры . Из полости рта пища попадает в глотку , а оттуда — через пищевод в желудок . Последний у большинства млекопитающих является простым (однокамерным); однако у некоторых млекопитающих — особенно у питающихся грубыми и трудно перевариваемыми растительными кормами — развился сложный (многокамерный) желудок, состоящий из нескольких обособленных отделов. Наиболее сложно в морфологическом и физиологическом плане он устроен у жвачных , обладающих четырёхкамерным желудком: в первых трёх отделах которого пища подвергается брожению с участием эндосимбионтов, а пищеварительные железы есть лишь в стенках четвёртого отдела, где пища и подвергается воздействию желудочного сока .
Кишечник млекопитающего подразделяется на тонкий и толстый (к последнему относятся слепая , ободочная и прямая кишка ) . Бо́льшая часть пищи переваривается в тонком кишечнике, через стенки которого основные питательные вещества всасываются в кровь . Остатки непереваренной пищи поступают в толстый кишечник, где они подвергаются бродильным процессам с участием эндосимбионтов — бактерий, грибков и простейших. Для видов, кормящихся грубой растительной пищей, особо важную роль в этих процессах играет слепая кишка, слабо развитая или вообще отсутствующая у плотоядных видов; стенки слепой кишки большинства млекопитающих содержат большое количество лимфатической ткани, из-за чего она играет ещё и важную роль в иммунной системе . В ободочной кишке каловые массы обезвоживаются, скапливаются в прямой кишке и затем удаляются из организма через анальное отверстие .

Почки млекопитающих , как и у других наземных позвоночных, тазовые ( метанефрические ) . Они обычно имеют бобовидную форму и располагаются в поясничной области, по бокам позвоночника. В наружном корковом слое каждой почки располагаются нефроны , в почечных клубочках которых происходит фильтрация плазмы крови и образуется моча , стекающая затем по мочеточникам в мочевой пузырь . Из него моча по мочеиспускательному каналу выходит наружу .
Почки регулируют количество воды в организме и выводят из него токсичные продукты обмена веществ. Конечными продуктами белкового обмена являются мочевина и мочевая кислота . Мочевина образуется в печени из аммиака , а мочевая кислота — продукт распада пуринов . При этом у млекопитающих мочевина по содержанию в моче резко преобладает над мочевой кислотой (у рептилий и птиц соотношение обратное). Помимо почек, определённую роль в выделении играют потовые железы и лёгкие , а также печень .
Почки млекопитающих также являются основным органом, участвующим в осморегуляции . Они могут выделять гипотоническую мочу при избытке воды и осмотически концентрированную — при её дефиците .

Эндокринную систему составляют железы внутренней секреции , регулирующие работу различных органов тела. Эти железы не имеют выводных протоков и выделяют вырабатываемые ими гормоны в кровь (реже — в лимфу) . У млекопитающих эндокринная система принципиально сходна с подобными системами других позвоночных, отличаясь не столько составом или числом гормонов, сколько усилением их активности и специализации .
К числу важнейших эндокринных желёз относят :
Поджелудочная железа и половые железы, кроме эндокринной секреции, осуществляют и экзокринную секрецию: помимо гормонов, поджелудочная железа выделяет также пищеварительные ферменты , а половые железы продуцируют гаметы . В связи с этим поджелудочную железу и половые железы относят к железам смешанной секреции . Множество гормонов образуются у млекопитающих в желудочно-кишечном тракте, почках, печени и других органах, не относящихся к эндокринной системе .

Оплодотворение у млекопитающих, как и у других амниот , внутреннее. Половые железы самок млекопитающих — яичники — всегда лежат в полости тела, в которую своими воронками открываются и яйцеводы . Во время овуляции созревшая яйцеклетка через полость тела попадает в фаллопиеву трубу — верхний извитой отдел яйцевода . Развитие яичников самок млекопитающих происходит циклически ( овариальный цикл ); вместе с ним циклическим изменениям подвергается эндометрий матки , обновляющийся в ходе каждого цикла ( эстральный цикл у большинства млекопитающих и менструальный цикл у человека и некоторых приматов) .
Половые железы самцов — семенники — имеют характерную овальную форму; у ряда видов они в течение всей жизни животного остаются в полости тела, но у большинства млекопитающих со временем опускаются в расположенный снаружи кожистый мешочек — мошонку . Вырабатываемые семенниками сперматозоиды , смешиваясь с семенной жидкостью , образуют сперму , которая во время полового акта через пенис самца попадает во влагалище самки (у однопроходных — в её клоаку ); после этого в верхнем отделе яйцевода — фаллопиевой трубе — и происходит оплодотворение ; образовавшаяся зигота тут же начинает дробиться и передвигается в матку .

Вынашивание плода у млекопитающих происходит в матке — нижнем отделе яйцевода . При этом у сумчатых и плацентарных во время беременности возникает плацента — специфический орган, образующийся путём срастания наружной оболочки зародыша ( хориона ) и разрыхлённого эпителия матки и обеспечивающий снабжение зародыша кислородом и питательными веществами, а также выведение углекислоты и продуктов обмена. Однако у сумчатых плацента достаточно примитивна (из-за чего у них период внутриутробной жизни короток, а детёныш рождается сильно недоношенным); у плацентарных же она намного более совершенна, что позволяет существенно продлить период внутриутробного развития, в течение которого плод надёжно защищён от неблагоприятных условий внешней среды .
Млекопитающие демонстрируют удивительное разнообразие приспособлений к обитанию в различных жизненных средах и к различным формам поведения. Ни один класс позвоночных не дал в процессе своей эволюции такого разнообразия форм, как млекопитающие, причём нередко виды, входящие в один и тот же отряд, относятся к различным адаптивным типам — основным экологическим группам; с другой стороны, представители разных отрядов, живущие в сходных условиях и близкие по своему образу жизни, могут иметь весьма сходный внешний вид ( конвергентное сходство ) .
В соответствии с приспособленностью к жизни в определённой среде среди млекопитающих выделяют следующие адаптивные типы: наземные, подземные, водные, древесные и летающие формы (а также переходные группы) .
Наземные звери — наиболее обширная группа млекопитающих. В пределах данной группы можно выделить две основные ветви: лесных зверей и зверей открытых местообитаний . И для тех, и для других характерными способами передвижения по твёрдому субстрату являются ходьба и бег (с подразделением их на разнообразные походки ) .
Лес служит местообитанием многих млекопитающих. Здесь они находят и пищу, и укрытие от врагов и непогоды. Некоторые из лесных зверей ведут строго наземный образ жизни ( лоси , косули , настоящие олени , росомахи , барсуки ); другие — так называемые полудревесные формы — способны и к лазанью по деревьям, где они могут добывать корм и отдыхать ( соболь , некоторые медведи , рыси , бурундуки ) .

Не менее многочисленную группу образуют обитатели открытых пространств ( лугов , степей , пустынь ). Часть из них приспособилась к быстрому бегу на четырёх конечностях. Это — большинство копытных ( куланы , зебры , верблюды , жирафы , сайгаки , бизоны ), а также некоторые хищные ( львы , гепарды , койоты ). Другая группа освоила зигзагообразный бег прыжками на длинных задних конечностях («рикошетирующий бег»): сумчатые тушканчики , многие кенгуру , прыгунчики , мешотчатые прыгуны , долгоноговые , тушканчики , тушканчиковые мыши . Наконец, третью группу образуют мелкие или средней величины звери с короткими ногами и вальковатым телом, которые живут в густой траве, затрудняющей передвижение ( сурки , суслики , хомяки , некоторые пищухи ). Бегают они медленно, а от опасности прячутся в норах .
Для некоторых видов наземных зверей ( обыкновенная бурозубка , волк , рыжая лисица , барсук ) характерен широкий ареал , охватывающий самые разнообразные ландшафты (виды- ). В этом случае явно выраженные приспособления к жизни в какой-либо определённой среде отсутствуют, и данные виды демонстрируют значительную экологическую гибкость . Так, барсук, ареал которого протянулся от северной тайги до пустынь и субтропиков , на севере ареала с октября по май погружается в спячку , а на юге в спячку не ложится .
Наибольшую скорость бега среди наземных зверей развивает гепард — до 105—115 км/ч. Максимальная скорость бега составляет: у льва и газели Томсона — 75—80, кулана — 65—70, зебры , дзерена и койота — 60—65, джейрана и волка — 55—60, дикого осла — 50—55, жирафа , зайца-русака и гигантского кенгуру — 45—50, бизона — 40—45, индийского носорога — 35—40 км/ч .

Многие млекопитающие фактически занимают по организации и образу жизни промежуточное положение между наземными и подземными формами. К таким полуподземным формам относятся звери, роющие более или менее сложные норы, но добывающие корм на поверхности земли: барсуки , кролики , сурки , суслики , аплодонтовые , гоферы , полёвки , трубкозубы , вомбаты . Приспособления к жизни под землёй у них выражены слабо. Обычно они заключаются в заметно укороченных ушных раковинах и развитой мускулатуре конечностей .
Настоящие подземные звери — это небольшая высокоспециализированная группа млекопитающих, которые проводят всю или бо́льшую часть жизни в толще почвы, находя там свой корм: либо обитающих в почве беспозвоночных, либо подземные части растений. Представители данной группы встречаются в разных отрядах. Сумчатые кроты , златокроты , кроты , цокоры прокладывают подземные ходы сильными передними конечностями с длинными «копательными» когтями, а слепыши и слепушонки не копают почву, а «выгрызают» её выступающими вперёд резцами . Грызуны же из семейства землекоповых действуют одновременно и передними лапами, и огромными резцами .
К числу общих морфологических особенностей подземных зверей относятся цилиндрическая форма тела с укороченным хвостом и полная или частичная редукция глаз и ушных раковин .

Среди млекопитающих большую группу составляют полуводные формы — виды, жизнь которых проходит частично на суше, частично в воде. Некоторые из таких животных ( норка , белый медведь , водяная полёвка , выдровая землеройка ) прекрасно плавают и ныряют, но сохраняют общий наземный облик. Их связь с водной средой проявляется в том, что свой корм они добывают не только на суше, но и в воде . Экологическую нишу полуводных форм млекопитающие начали осваивать ещё в середине мезозоя : специализация к полуводному образу жизни характерна, в частности, для докодонта касторокауды из средней юры .

Диагностическим признаком для следующей (по степени адаптации к жизни в воде) группы полуводных зверей служит наличие более или менее развитой плавательной перепонки между пальцами конечностей. Такая перепонка имеется у утконоса , водяного опоссума , выхухоли , многих грызунов ( нутрия , капибара , бобр , бобровая крыса , ондатра и др.), выдры , гиппопотама . У куторы и водяной бурозубки плавательную перепонку заменяет плавательная оторочка из жёстких волосков. Для указанных видов характерны также укорочение шеи и ушных раковин, развитие приспособлений для замыкания ушей и ноздрей, уплощение хвоста, наличие густого меха, противостоящего намоканию, или толстого подкожного жирового слоя .
Ещё более высокую степень приспособления к жизни в водной среде демонстрируют каланы и ластоногие . Толстая кожистая перепонка между пальцами у них развита настолько, что их конечности (у калана — только задние) превращаются в ласты . Пищу эти животные добывают только в воде, однако спариваются и размножаются (а иногда и отдыхают) они вне водной среды .
Полностью водными зверями , никогда не покидающими водной среды, являются сирены и китообразные . У них полностью отсутствует меховой покров, передние конечности превратились в плавники, а задние утрачены. Основным локомоторным органом служит кожистый хвостовой плавник . Если у утконоса и калана максимальная продолжительность пребывания под водой равна 8—10 мин., у ондатры и выхухоли — 12 мин., бобра — 15 мин., моржа и ламантина — 16 мин., то у дельфинов — 20 мин., кашалота — 75 мин., а у бутылконоса она достигает 2 часов. Засвидетельствованы случаи погружения кашалота на глубину 988 м , а некоторые источники указывают, что он способен погружаться даже на 3 км .
Надо сказать, что характеристика китообразных как полностью водных форм относится к ныне живущим представителям данного инфраотряда. Приспособление китообразных к строго водному образу жизни шло постепенно. Пакицет из раннего эоцена Пакистана передвигался на четырёх конечностях и проводил часть времени в воде, охотясь за рыбой, но для отдыха выходил на сушу .

К древесным зверям , иначе именуемым дендробионтами , относят млекопитающих, проводящих бо́льшую часть своей жизни в кронах деревьев, где они добывают пищу, спасаются от врагов, устраивают гнёзда для размножения и отдыха. Основные способы передвижения у них — лазанье по стволам и ветвям деревьев и прыжки с ветки на ветку .
Приспособления для жизни на деревьях у дендробионтов разнообразны. Многие из них используют для лазанья длинные искривлённые когти; при этом одни из них имеют длинный пушистый хвост, которым пользуются как балансиром и парашютом при прыжках ( сумчатые куницы , щёткохвостые поссумы , древесные кенгуру , белки , сони , некоторые куницы ) , у других хвост служит хватательным органом, позволяющим зафиксировать положение тела на ветке ( поссум-медоед , кускусы , карликовый муравьед , белобрюхий ящер , древесные дикобразы , кинкажу ) , у третьих хвост рудиментарен и в передвижении не участвует ( коала , ленивцы ) .
Ряд древесных зверей использует для лазанья хватательные конечности с сильно развитыми пальцами, причём один (у лемуров , долгопята , многих обезьян ) или два пальца (у коалы , кускусов ) противопоставлены остальным (интересно, что долгопят имеет на концах пальцев присасывательные подушечки, что усиливает хватательную функцию конечностей). Как правило, такие дендробионты хватаются за ветви всеми четырьмя конечностями. Однако гиббоны и коаты используют для передвижения по ветвям и перепрыгивания с дерева на дерево лишь передние конечности, а коаты — также и хватательный хвост .
Если белки и сони вполне сохраняют способность передвигаться по земле и хорошо бегают, то более узкоспециализированные дендробионты передвигаются по земле плохо, а ленивцы такую способность утратили почти полностью .

Ещё одним видом способов передвижений, освоенных млекопитающими, является полёт : как пассивный (планирующий), так и активный (с помощью крыльев). По способности к той или иной из этих разновидностей полёта, отражающей принципиально различный уровень освоения воздушной среды, среди летающих зверей выделяют соответственно полувоздушные и настоящие воздушные формы .
Полувоздушные формы млекопитающих относятся к разным семействам и отрядам, но обладают общим планом строения. Речь идёт о наличии летательной перепонки — кожистой складки, которая тянется по бокам туловища между передними и задними лапами и позволяет не только планировать при спуске по отлогой линии, но и совершать повороты в воздухе. К таким формам принадлежат: триконодонт волатикотерий из поздней юры Китая, некоторые двурезцовые сумчатые ( , полосатые кускусы , гигантский летучий кускус , карликовый летучий кускус ), некоторые грызуны ( летяги , большинство шипохвостых ), шерстокрылы (у последних летательная перепонка достигает наибольшего развития, охватывая и хвост). Если у гигантской летяги дальность полёта достигает 60 м, то у шерстокрыла — 140 м .
К
настоящим воздушным формам
принадлежат многочисленные представители отряда
рукокрылых
, у которых передние конечности превращены в длинные кожистые крылья. Рукокрылые способны длительное время, быстро и с хорошей манёвренностью передвигаться в воздухе, добывать там же себе пищу и даже спариваться
.
Грудина
рукокрылых имеет
киль
, как и у птиц, служащий для прикрепления развитых грудных мышц
. Наиболее развитыми летательными способностями обладают представители семейства
складчатогубых
и, в частности,
бразильский складчатогуб
: его полёт — лёгкий и стремительный, причём средняя скорость полёта составляет 40 км/ч, а максимальная — 100—105 км/ч
.
По роду употребляемой пищи млекопитающих можно подразделить на две условные группы: растительноядных и плотоядных . Условность данного деления состоит в том, что лишь немногие виды млекопитающих питаются исключительно растениями или исключительно животными; у большинства в рационе присутствуют и растительные, и животные корма, так что речь идёт лишь о преобладании тех или других . Если такое преобладание не выражено, говорят о всеядных животных — таковы бурый медведь , енот , барсук , соболь , кабан , серая крыса и др.; в их питании более или менее существенную роль играют беспозвоночные и позвоночные животные, ягоды, фрукты, орехи, зелень. С другой стороны, некоторые виды специализированы на употреблении весьма ограниченного набора кормов .
Среди растительноядных млекопитающих выделяются :

Среди плотоядных млекопитающих выделяются :
С особенностями пищедобывания и питания тесно связан целый комплекс морфолого-физиологических особенностей, характерных для отдельных видов млекопитающих (цедильный аппарат усатых китов , вытянутая морда и длинный клейкий язык у питающихся насекомыми трубкозубов , муравьедов и панголинов , способность насекомоядных рукокрылых преследовать, пользуясь ультразвуковой локацией, летящих насекомых, очень острые обоняние и слух лисицы , втяжные острые когти кошачьих и др.). Эти особенности тесно коррелируют с другими функциями организма, уже не имеющими к питанию прямого отношения, налагая на них свой отпечаток .

Среди млекопитающих широко распространён половой диморфизм , то есть явление, при котором разнополые особи одного вида отличаются друг от друга. У млекопитающих известны примеры полового диморфизма по следующим признакам:
Сроки наступления половозрелости у млекопитающих очень сильно варьируют. Приведём сроки полового созревания для некоторых видов (в скобках дана примерная продолжительность жизни в природе, лет): рыжая полёвка — 1 мес. (1—1,5), степная пеструшка — 1,5 мес. (1—1,5), домовая мышь — 2—3 мес. (около года), горностай — 3 мес. (4—7), заяц — 1 год (7—9), лисица — 1—2 года (10—12), волк и лось — 2—3 года (15—20), бурый медведь — 3—4 года (30—50), коала — 4 года (12—13), синий кит — 4—5 лет (до 20 лет), индийский слон — 10—15 лет (70—80) .

Спаривание млекопитающих обычно приурочено к определённому времени года . В умеренных и высоких широтах сигналом к наступлению брачного периода служит изменение длины светового дня, в низких широтах — изменение интенсивности осадков, созревание основных кормовых растений и т. п. При поиске особи противоположного пола у одних видов млекопитающих бо́льшую активность проявляют самки, у других — самцы. Многие звери отыскивают друг друга по запаху, причём в брачный период усиливается деятельность пахучих желёз. Распространены и звуковые сигналы: у лисиц и кошачьих голос подают и самцы, и самки, у оленей ревут только самцы, а у лосей первой о месте своего нахождения характерным храпом извещает самка .
Для большинства видов млекопитающих характерна полигамия в форме полигинии . У основной массы видов ( насекомоядные , летучие мыши , многие грызуны и хищные ) самцы после спаривания не удерживают самок возле себя. У ушастых тюленей и стадных копытных на время размножения образуются гаремы или косяки с единственным взрослым самцом во главе, а у некоторых хищных и грызунов возникают сложные семьи (парцеллы грызунов, прайды львов) из нескольких самцов и самок с системой рангов и разделением обязанностей. Редкое явление — полиандрия (многомужество), известная у ряда летучих мышей и некоторых обезьян (например, у большинства игрунковых ) . Своеобразно устроена семейная группа у голых землекопов : она состоит из нескольких десятков особей и организована по типу муравьиной семьи: во главе стоит размножающаяся самка, которая в 1,5—2 раза превосходит по размерам остальных членов семьи и почти всё время проводит в гнездовой камере в компании с 2—3 неработающими самцами, призванными её оплодотворять; остальные взрослые члены семьи заняты добыванием пищи и рытьём и обустройством нор .
Значительно меньшую группу образуют моногамные виды, причём пары обычно образуются на короткий срок ( шакалы , лисицы , песцы , бобры , ондатры ), реже — на несколько лет ( волки ). Только немногие виды обезьян могут образовывать пары на всю жизнь .
По мере приближения родов самка становится беспокойной и ищет удобное укромное место. У многих видов она подготавливает гнездо или логово для будущего потомства. У большинства млекопитающих самка рожает в лежачем положении; самки зайцев рожают сидя, а самки слонов и многих парнокопытных — стоя. Большинство китообразных рожает под водой, но самки некоторых видов ( гренландский кит , белуха и др.) принимают при родах вертикальное положение, выставив из воды хвостовую часть тела так, что их детёныш появляется на свет в воздухе. Самки рукокрылых рожают в висячем положении: ушаны — либо в горизонтальном положении (брюхом кверху), либо в вертикальном (головой вверх), а подковоносы и крыланы — вися вниз головой; появившийся детёныш попадает в подогнутую к брюху межбедренную перепонку или же в полость между брюхом и сложенными крыльями. У двупалого ленивца самка при родах висит на ветке, держась за неё передними лапами, а детёныш, цепляясь когтями за шерсть, сам добирается до соска матери .
По частоте деторождения виды млекопитающих делят на моноэстричные , которые рожают детёнышей раз в год (большинство сумчатых , насекомоядных , хищных , копытных , зубатых китов ) или раз в 2—3 года ( бурые медведи , тигры , моржи , зубры , слоны , большинство усатых китов ) , и полиэстричные , рожающие несколько раз в году ( зайцы-беляки и белки приносят по 2—3 помёта, полёвки — по 3—4, домовая мышь — до 6—8) . Различна и величина выводка: если у слонов, китов, моржей, зубров, лошади , гигантского кенгуру рождается по одному детёнышу, то в помёте у рыси , соболя , бурого медведя обычно по 2—3 детёныша, волка, зайца, белки — от 3 до 8, виргинского опоссума — от 8 до 18 . Максимальное для плацентарных число детёнышей в одном помёте бывает у обыкновенного тенрека : у одной самки обнаружили 32 эмбриона , а другая в неволе родила и вскормила 31 детёныша (при этом число сосков у самки тенрека может доходить до 29 — больше, чем у любого другого млекопитающего) .
Для многих видов отмечается зависимость величины выводка от географических факторов. Так, у длиннохвостого суслика средняя величина выводка возрастает в направлении с юга на север, составляя: в Западной Монголии — 4,0; в Прибайкалье — 6,9; в Центральной Якутии — 8,2. У оленьего хомячка , обитающего на юго-западе США , она изменяется с увеличением высоты: на высоте 1,0—1,5 тыс. м она равна 4,6; на высоте 1,7—2,0 тыс. м — 4,4; на высоте 2,5—3,0 тыс. м — 5,4; на высоте 3,2 тыс. м — 5,6 .

Млечные железы самки в период беременности увеличиваются в размерах и к моменту родов достигают максимального развития. В первые дни лактации из млечных желёз выделяется молозиво — густая и вязкая жидкость с повышенным, по сравнению с молоком, содержанием белков и витаминов. Затем оно заменяется молоком , химический состав которого у однопроходных близок к составу пота, а у других групп млекопитающих приобретает свои характерные особенности и варьирует достаточно сильно. В приводимой ниже таблице приведены данные для отдельных видов, представляющих различные отряды сумчатых и плацентарных :
| Состав молока млекопитающих | |||||||
|---|---|---|---|---|---|---|---|
| Виды животных | Состав молока, % | Энергетическая ценность | |||||
| вода | белки | жиры | сахар |
минеральные
вещества |
ккал / 100 г | кДж / 100 г | |
| Гигантский кенгуру | 88,0 | 4,6 | 3,4 | 6,7 | 5,3 | 76 | 318 |
| Индийский слон | 78,1 | 4,9 | 11,6 | 4,7 | 0,7 | 143 | 599 |
| Кролик | 67,2 | 13,9 | 15,3 | 2,1 | 1,5 | 202 | 846 |
| Человек | 87,1 | 0,9 | 4,5 | 7,1 | 0,4 | 72 | 301 |
| Кошка | 82,2 | 9,1 | 3,3 | 4,9 | 0,5 | 86 | 360 |
| Собака | 76,4 | 7,4 | 10,7 | 3,3 | 2,2 | 139 | 582 |
| Морской котик | 34,6 | 8,9 | 53,3 | 0,1 | 3,1 | 516 | 2160 |
| Лошадь | 88,8 | 2,5 | 1,9 | 6,2 | 0,6 | 52 | 218 |
| Свинья | 81,2 | 4,8 | 6,8 | 5,5 | 1,7 | 102 | 427 |
| Афалина | 58,3 | 6,8 | 33,0 | 1,1 | 0,8 | 329 | 1377 |
| Северный олень | 67,7 | 10,1 | 18,0 | 2,8 | 1,4 | 214 | 896 |
| Овца | 82,0 | 4,6 | 7,2 | 4,8 | 1,4 | 102 | 427 |
| Коза | 86,7 | 3,2 | 4,5 | 4,3 | 1,3 | 70 | 293 |
| Корова | 87,3 | 3,2 | 3,9 | 4,6 | 1,0 | 66 | 276 |

В действительности химический состав молока несколько различается и у отдельных представителей одного и того же вида (в таблице приведены усреднённые данные). При этом особенно высока энергетическая ценность , или калорийность, молока у морских млекопитающих — ластоногих и китообразных; среди копытных высоких значений она достигает у северного оленя .
Длительность лактационного периода у разных видов млекопитающих варьирует в больших пределах. Зайчата уже через 7—8 дней начинают есть траву, хотя сосать материнское молоко продолжают. У грызунов и хищных лактация длится сравнительно недолго, но у северного оленя она продолжается 4—5 мес., коровы и лошади — 9—10 мес., моржа — не менее года .
После перехода молодняка на самостоятельное питание его рост заметно замедляется. Темпы роста сильно зависят от образа жизни млекопитающего и внешних условий. Суровые условия, характерные для местообитаний полярных и высокогорных животных, требуют быстрого развития детёнышей. Так, новорождённый северный олень уже в первый день в состоянии следовать за матерью, а новорождённый благородный олень делает это не ранее достижения недельного возраста. Весьма быстро растёт молодняк у рукокрылых и многих грызунов , а вот развитие детёнышей медвежьих происходит медленно .
Забота о подрастающем потомстве млекопитающих в основном лежит на самках, движимых материнским инстинктом, хотя у ряда видов, особенно моногамных, подключаются и самцы. Самки вскармливают своих детёнышей молоком, согревают теплом своего тела, благоустраивают гнёзда и норы, переносят на новое место в случае необходимости, защищают от врагов, наказывают за непослушание . Особенно характерно для млекопитающих систематическое обучение молодняка с передачей потомству накопленного родителями индивидуального опыта .
У млекопитающих, живущих на территориях с выраженной сменой времён года , чётко выражен годовой цикл жизни с приспособлением к сезонным изменениям природной обстановки. Обычно период деторождения оказывается приуроченным к наиболее благоприятному периоду (для зверей северных и умеренных широт это — конец весны и начало лета). В процессе эволюции данный факт обусловил (с учётом длительности беременности) и период спаривания, характерный для того или иного вида .
Появлению детёнышей обычно предшествует выбор места, особенно удобного для деторождения и воспитания молодняка. В это время даже постоянно мигрирующие виды переходят к оседлости, их образ жизни становится более скрытным. После подрастания молодняка звери, не привязанные к постоянному жилищу, вновь перемещаются — в места, наиболее богатые кормом .
Приспособления млекопитающих к переживанию зимы (иногда — других неблагоприятных периодов года) многообразны. Многие звери осенью накапливают жировые резервы . Так, подкожный и внутренний жир у степного сурка в июне составляет 10—15 г, в августе — 750—800 г). Нередко звери претерпевают линьку , при которой более редкий летний мех сменяется более густым, а у многих тёмноокрашенных летом видов — зайца-беляка , белого песца , горностая и др. — зимний мех белый, что делает зверька малозаметным на снегу . Кроме того, некоторые представители совершают значительные сезонные миграции на юг ( северные олени , зайцы-беляки , песцы и др.), другие — запасают корм ( лесные мыши , полёвки , цокоры , слепыши , бобры , белки , хорьки и др.) . Многие копытные , хищные , грызуны объединяются к зиме в более крупные группы (стада, стаи) .
Немало млекопитающих впадает на зиму в спячку , позволяющую снизить расходы энергии в период, когда поиск пищи затруднён. Различают зимний сон (факультативная спячка), при которой уровни метаболизма, температуры тела и дыхательной активности снижены незначительно ( медведи , енотовидные собаки , еноты , барсуки в северных частях своих ареалов), и настоящую спячку с сильным оцепенением и значительным снижением температуры тела и интенсивности метаболизма ( ежи , ряд летучих мышей , сурки , суслики , хомяки ). У некоторых грызунов известна и летняя спячка, приуроченная к засушливому периоду .

Характерный для млекопитающих высокий уровень развития высшей нервной деятельности обусловливает сложные формы их поведения. Во-первых, бо́льшую по сравнению с другими позвоночными сложность и подвижность у млекопитающих обретает инстинктивная деятельность, механизмы которой основаны на простых безусловных рефлексах и их сложных сочетаниях ( инстинктах ). Во-вторых, млекопитающие, особенно высшие плацентарные с присущей им высокой степенью развития коры полушарий переднего мозга, отличаются выраженной способностью к тонкому анализу и восприятию внешних воздействий, к предвидению хода событий, к выработке условных рефлексов и накоплению индивидуального опыта .
Отмеченные особенности поведения млекопитающих проявляются в различных сферах их жизни. Многие из них активно пользуются различного типа убежищами — при отдыхе, для защиты от нападений хищников и от неблагоприятных воздействий среды, запасания кормов и выращивания потомства. Наиболее совершенный тип убежищ — постоянные норы , нередко отличающиеся сложным строением. Так, у барсуков норы — это нередко целые системы ходов, расположенных в несколько этажей и включающих гнездовую камеру, боковые отнорки, используемые как уборные, вентиляционные отнорки. Норы хомяков включают вместительные кладовые для хранения запасов корма (до 5 в одной норе). Подобные кладовые имеются также в норах даурских пищух , серых полёвок , степных пеструшек , больших песчанок и ряда других грызунообразных .

Норы бобров , ондатр , выхухолей — это переплетающиеся системы галерей с подводным расположением входов, причём нижние камеры заполнены водой; имеются гнездовые камеры и вентиляционные отнорки. При этом бобры и ондатры роют норы при наличии у водоёмов крутых берегов, а на низких топких берегах они строят для жилья наземные хатки — куполообразные сооружения из сучьев или стеблей растений. Если уровень воды в водоёме непостоянен, бобры регулируют уровень воды, возводя из срезанных стволов деревьев, веток и хвороста плотины — хорошо известный пример сложной строительной деятельности млекопитающих (длина такой плотины обычно составляет 20—30 м, достигая в исключительных случаях 600 м). Иногда бобры прорывают каналы , по которым сплавляют заготовленную древесину .
Многие млекопитающие метят капельками мочи, экскрементами или выделениями пахучих желёз занимаемые ими участки обитания , создавая тем самым «сигнальные поля». При этом они нередко ожесточённо отстаивают свои участки от вторжения других животных; например, схватки бобров с вторгающимися на их территорию другими бобрами часто завершаются гибелью пришельца от укусов. В то же время мечение является и средством смягчения внутрипопуляционной напряжённости: нарушитель помеченных границ ведёт себя настороженно и готов к отступлению при попытке хозяина защитить свои владения .
Значительной сложностью отличается поведение млекопитающих в брачный период. Нередко самки заигрывают с самцами, принимают специальные позы. Самцы же привлекают внимание самок характерными телодвижениями и звуками (крики, щёлканье зубами, удары ног о землю). Для многих видов характерны драки самцов за обладание самкой. У оленей и многих полорогих такие драки принимают форму «брачных турниров» с применением рогов, причём обычно (но не всегда) такие турниры заканчиваются трагическим исходом .
Для млекопитающих весьма характерно наличие игрового поведения , особенно у детёнышей. Игра помогает приобрести полезные навыки и служит средством консолидации в семейных группах и других объединениях животных . Началу игры у львов и псовых предшествует поза с прижатыми к земле передними лапами, которая не встречается в других ситуациях и сигнализирует, что последующие внешне агрессивные действия — всего лишь игра. Обезьяны в подобных случаях прибегают к своеобразной игровой мимике .
Единственная возможность не стать добычей хищника у мелких и средних млекопитающих — спрятаться или убежать. Однако и здесь возможны вариации тактики. Так, зайцы , направляясь на лёжку, запутывают следы, делая «вздвойки» (попятные проходы по своим следам) и «скидки» (большие прыжки в сторону); получающиеся тупики следа затрудняют выслеживание зайца хищниками .

Усложнение внутривидовых связей у многих видов млекопитающих приводит к образованию временных или стойких группировок особей с объединением усилий по использованию ресурсов среды и защите от врагов .
Те млекопитающие, у которых источники пищи очень рассредоточены, а жилищами служат постоянные убежища, обычно ведут одиночный или семейный образ жизни. Таковы однопроходные , сумчатые , насекомоядные , многие грызуны и хищные ; среди приматов — большинство низших приматов и широконосых обезьян , гиббоны и орангутаны .
Для крупных подвижных животных, особенно копытных , характерно образование кочующих стад, стай или кланов. Для социальной организации таких групп типично наличие сложной структуры, связанной с ранжированием особей, приобретающих разное положение в группе: «доминант» (вожак), «субдоминанты» и «подчиняющиеся»; при этом доминант теряет ранг в случае ошибки, болезни или появления более сильного соперника. В некоторых группах намечается разделение обязанностей. Так, в стаях ( прайдах ) львов самцы в основном заняты охраной участка обитания от вторжения чужаков, а самки добывают пищу и заботятся о молодняке . У некоторых хищных (например, у пятнистой гиены ) в кланах наблюдается матриархат : самки крупнее самцов и доминируют над ними .
Львы — единственные социальные животные среди кошачьих . Среди других хищных стаями живут волки , стадами — большинство ластоногих . Иногда же несколько семей объединяются для совместной охоты ( гиеновидные собаки , красные волки ). При совместной охоте стаи волков за крупной дичью обязанности охотников чётко распределены, а их действия скоординированы: одни гонят будущую добычу, другие нападают на неё из засады или перерезают ей путь . Гиеновидные собаки при совместной охоте на крупных животных также координируют свои действия: во время погони лидеры чередуются, стремясь равномернее разделить тяжесть погони, бегущие сзади собаки срезают углы, направляясь наперерез жертве; часть взрослых собак в охоте не участвует, охраняя молодняк, однако добыча делится на всех. Стадом действуют и косатки , нападающие на крупную добычу (например, кита) .

Антилопы импалы , узконосые обезьяны гелады и некоторые другие виды обладают сходной социальной организацией, образуя стада холостяков из одиноких разновозрастных самцов и гаремные стада из самок под контролем одного самца. Относительно крупными стадами (иногда до 100 и более особей) с чётко выраженной иерархией живут другие узконосые обезьяны — павианы (гамадрилы, бабуины, анубисы). Здесь доминантные самцы обладают преимущественным правом доступа к самкам и пище; при этом у гамадрилов стадо, как правило, возглавляет один самец-вожак, для бабуинов типично наличие группы объединившихся доминантных самцов, а анубисы занимают промежуточное положение .
Гориллы живут небольшими стадами (5—30 особей); эти группы включают самца-вожака, одного или нескольких молодых самцов, нескольких взрослых самок и детёнышей. Иерархические отношения здесь проявляются в порядке следования по тропам, причём вожак идёт во главе цепочки. В остальном же вожак ведёт себя как покровитель, но не деспот, и, в отличие от гамадрилов, не является владыкой гарема: отношения полов у горилл носят добровольный характер. Шимпанзе живут либо малыми семьями, либо небольшими стадами; особой иерархии между индивидами, входящими в стадо, не отмечается .

У копытных стадный образ жизни облегчает поиск пищи, миграции и оборону от хищников. Известны случаи, когда антилопы канны в «сомкнутом строю» прогоняли гепардов . Стадо овцебыков не убегает от хищников ( волков или медведей ), а образует замкнутый круг с телятами внутри и взрослыми животными снаружи; последние успешно обороняются от хищников рогами и копытами. Стадо индийских буйволов , почуяв след тигра , приходит в неистовство и в сомкнутом строю преследует врага, пока не настигнет хищника или же не потеряет его след. Африканские буйволы , действуя дружным стадом, способны обратить в бегство даже прайд львов .
Промежуточное положение между одиночным и стадным образом жизни занимает образ жизни млекопитающих, образующих колонии : даманы , рукокрылые , некоторые грызунообразные ( кролики , пищухи , суслики , сурки , шиншиллы и др.); среди хищных колониями живут сурикаты . Здесь животные, находясь в тесном сожительстве и постоянном общении, имеют свои индивидуальные и семейные участки, но при этом разделённая на отдельные группировки популяция не теряет своей целостности . Колонии могут включать несколько десятков или сотен животных; особенно крупные колонии, насчитывающие многие тысячи особей, характерны для рукокрылых . Рекордсменом в этом отношении является бразильский складчатогуб : в некоторых пещерах на юге США он образует колонии численностью до 20 млн особей — самые крупные на Земле скопления млекопитающих .

Среди всех групп позвоночных млекопитающие выделяются наивысшим уровнем развития интеллекта , что связано прежде всего с прогрессивным развитием коры полушарий переднего мозга, хотя этот уровень существенно различается у млекопитающих разных систематических групп . Одним из показателей умственного развития, хорошо применимым к млекопитающим, является коэффициент цефализации (отношение возведённой в квадрат массы мозга к массе всего тела). По этому показателю лидирующее место занимает человек , а второе — дельфины , у которых значение данного показателя примерно в два раза меньше .
Впрочем, большинство исследователей при оценивании интеллекта различных животных отдают первенство не нейроанатомическому, а функциональному подходу . В рамках последнего изучают процессы принятия млекопитающими решений, требующих прогноза конкретной ситуации (в этом плане наибольшие способности показали медвежьи , дельфиновые и высшие приматы ), анализируют общую организацию их коммуникативных систем (здесь выяснилось, что у большинства млекопитающих сигнальные системы не имеют принципиальных отличий от подобных систем других позвоночных, но существенно более высокий уровень демонстрируют узконосые обезьяны , особенно человекообразные ) .
В ряде случаев исследователям удалось наблюдать возникновение у млекопитающих новых форм поведения и распространение такого поведения среди популяции. Так, при наблюдениях за обитающими на острове японскими макаками выяснилось, что самка по кличке Имо последовательно открыла способы отмывания от песка в воде ручья батата и зёрен злаков , после чего новую технологию постепенно переняли и другие макаки, жившие на острове . У человекообразных обезьян наличие развитых средств коммуникации привело к появлению общественного обучения : исследователи наблюдали, как зрелые шимпанзе обучали молодых ловить термитов с помощью просовываемой в ходы термитника соломинки. Интересно, что находившиеся рядом павианы , которые также любят лакомиться термитами, внимательно наблюдали за действиями шимпанзе, но освоить данный приём были не в состоянии .
Учёные, изучавшие язык млекопитающих , выявили наличие в нём, как в человеческом языке , плана выражения и плана содержания . В то же время у низших плацентарных этот язык весьма беден: так, язык желтобрюхого сурка содержит всего 8 звуковых сигналов, содержание которых сводится к пяти «смыслам» («внимание!», «тревога!», «угроза», «страх», «удовольствие») . У макак выделено уже свыше 30 звуковых сигналов, дополняемых мимикой и жестами (например, выпрямление хвоста — жест доминирования) .
.jpg)
Намного бо́льшими интеллектуальными способностями обладают человекообразные обезьяны. В ходе масштабных так называемых « языковых проектов » этологи обучали шимпанзе , бонобо и горилл знакам человеческого языка; в качестве означающих выступали знаки языка глухонемых , специально подобранные предметы или изображения на экране компьютера. В результате лексикон шимпанзе Уошо составил несколько десятков знаков, причём она могла самостоятельно изобретать новые знаки и их сочетания (комбинацией «вода — птица» Уошо назвала впервые встреченного ею на прогулке лебедя), а также употреблять знаки в переносном значении . Горилла Коко в зрелом возрасте могла выразить более 500 понятий .
Выяснилось, что человекообразные обезьяны способны понимать разницу между субъектом и объектом . Так, Роджер Фоутс , работавший в рамках «языкового проекта» с шимпанзе , однажды вместо привычной фразы «Роджер — щекотать — Люси» просигнализировал: «Люси — щекотать — Роджер». Люси сначала попыталась его поправить, но когда тот повторил фразу, пощекотала экспериментатора . Результатом «языковых проектов» стал вывод о том, что интеллект человекообразных обезьян достаточен для овладения человеческим языком на уровне ребёнка первых лет жизни .
Млекопитающим удалось широко освоить все экосистемы Земли . В биоценозах суши и морей они занимают экологические ниши как первичных потребителей растительных кормов, так и плотоядных животных. Среди позвоночных млекопитающие по численности и биомассе в водоёмах уступают лучепёрым рыбам , а на суше удерживают первое место. Мелкие млекопитающие (особенно полёвки и мыши ) могут в годы подъёмов численности достигать плотности в 100—200 зверьков на 1 га ; численность же крупных копытных и хищных обычно не превышает 1—5 на 1000 га .
Представляя верхние звенья пищевых цепей , млекопитающие оказывают влияние на эволюцию растений . У одних растений вырастают колючки , а в листьях и плодах образуются ядовитые алкалоиды , что препятствует поеданию. Другие виды растений приспособились к распространению семян и плодов при посредстве млекопитающих: речь идёт или о появлении у семян и плодов цепких шипиков и крючков ( репейник , череда ) для прикрепления к шерсти животных, или о привлечении внимания последних цветом, ароматом и вкусом плодов. Многие семена не теряют при прохождении через пищеварительный тракт животного своей всхожести , а у семян калины и рябины последняя даже повышается: пищеварительный сок разрыхляет деревянистую оболочку семени .
Деятельность млекопитающих во многом определяет устойчивость биогеоценозов . Так, в пустынях и степях вымирание или истребление роющих грызунов ( полёвки , песчанки ) влечёт разрастание немногих видов растений- многолетников и сокращение числа видов однолетников-эфемеров , что обедняет пастбища .
Некоторые млекопитающие (в основном грызуны ) имеют существенное эпидемиологическое значение, будучи переносчиками возбудителей болезней, опасных для человека и других млекопитающих ( чума , туляремия и др.) .

Угрозу для существования тех или иных родов, видов и подвидов млекопитающих могут составлять как естественные (природные катаклизмы , глобальные изменения климата ), так и антропогенные (хищнический промысел , загрязнение окружающей среды , разрушение биотопов ) факторы. Ряд характерных для плейстоценовой фауны родов — мамонт и шерстистый носорог в Евразии , саблезубый смилодон и гигантские наземные ленивцы ( мегатерий , мегалоникс , ) в Америке , дипротодон в Австралии — вымерли (не без участия человека) по окончании ледникового периода (10—12 тыс. лет назад) .
В историческую эпоху исчезло свыше 100 видов крупных млекопитающих, в том числе 33 вида — до 1700 г., 36 видов — в XVIII—XIX вв. и около 40 — в 1-й половине XX в. Среди них: тур (последний представитель погиб на Львовщине в 1627 г.), стеллерова корова (последняя из них убита в 1768 г. на острове Беринга ), квагга (последняя квагга умерла в 1883 г. в Амстердамском зоопарке ), тарпан (последняя самка тарпана умерла в Московском зоопарке в конце 1880-х гг.) .
В начале XXI века под угрозой уничтожения из-за бесконтрольной охоты находятся лось , кабан и европейская косуля в средней полосе России , сайга и джейран в Средней Азии ; распашка прерий в Северной Америке привела к почти полному уничтожению луговых собачек и американских хорьков ; загрязнение озера Байкал промышленными стоками поставило на грань исчезновения байкальскую нерпу .


Для сохранения биологического разнообразия млекопитающих нужна целенаправленная законодательная и практическая работа. Первой в мире мерой по охране диких животных стало принятое в 1868 году земским сеймом во Львове и властями Австро-Венгрии решение об охране серн и сурков в Татрах .
Одной из важнейших природоохранных мер стала организация заповедников , в которых ведётся работа по сохранению и восстановлению численности разных животных. На территории Российской империи первый частный заповедник — заповедник «Аскания-Нова» — основал на территории своего имения Ф. Э. Фальц-Фейн в 1889 году .
В 1966 году была издана « Международная Красная книга », а в 1978 году — « Красная книга СССР »; в них были приведены списки исчезающих и редких видов и подвидов с рекомендациями по их охране (в 1983 году последовала «Красная книга РСФСР» , а в 2001 году — « Красная книга России » ). По состоянию на февраль 2014 года в Международной Красной книге имеются данные по численности 5506 видов млекопитающих, из них 77 считаются полностью вымершими (категория EX), 2 вида признаны вымершими в дикой природе (категория EW), и ещё 1143 находятся под угрозой различной степени риска (категории LR, VU, EN, CR) . В Красную книгу России занесено 74 вида млекопитающих .

Охота представляет собой одну из древнейших форм хозяйственной деятельности человека. Как показывает анализ костей животных, найденных на стоянках людей эпохи нижнего палеолита , в это время основными объектами охоты были: в тропической зоне — антилопы , дикие быки , тапиры , гиппопотамы , слоны ; в более северных районах — зубры , газели , олени , кабаны , дикие лошади , мамонты , шерстистые носороги , пещерные медведи , пещерные львы . Эти звери служили источником мяса, жира, костного мозга и шкур для изготовления одежды .
Со времён неолитической революции охота на диких млекопитающих перестаёт в большинстве регионов Земли быть основой хозяйства, но сохраняет определённую значимость до сих пор . В настоящее время наибольшее количество видов, служащих объектами промысловой и спортивной охоты, относится к отрядам хищных , грызунов и китопарнокопытных .
Заслуживает упоминания также морской зверобойный промысел. Охота на морского зверя : моржей , ушастых и настоящих тюленей , калана , китов — издавна являлась основой традиционного хозяйства береговых чукчей , коряков , эскимосов , алеутов . Ныне добыча морского зверя в России ведётся на специально оборудованных судах, причём размеры добычи ограничены международными соглашениями .
В конце XIX века возникла и получила со временем значительное развитие новая отрасль хозяйства — пушное звероводство . Оно предполагает разведение пушных зверей — соболя , норку , песца , лисицу , нутрию , шиншиллу — на зверофермах , где животные размещаются в сетчатых клетках . Первые опыты такого разведения были осуществлены в 1895 году в Канаде с серебристо-чёрными лисицами , очень редкими в природе .
Из других зверей на фермах разводят пятнистых оленей и маралов , а также лосей .
Бо́льшая часть одомашненных видов животных относится к млекопитающим, а из них — к китопарнокопытным , а именно к группе парнокопытных . Среди других отрядов одомашнены представители зайцеобразных ( домашний кролик ), грызунов ( морская свинка ), хищных ( собака , кошка , фретка ), и непарнокопытных ( лошадь , осёл ) . Одомашнивание и сопровождавшая его целенаправленная селекционная работа вели к появлению существенных отличий домашних животных от своих диких предков: так, у рогатого скота и свиней значительно увеличились вес и продуктивность, чрезвычайно сильно различаются по размерам и внешнему виду различные породы собак и кошек . Впрочем, проведение чёткой границы между только что приручённой и вполне одомашненной формой возможно не всегда; поэтому приведённые далее сведения о времени и месте одомашнивания не следует считать окончательными .

Первым домашним животным стала собака , одомашненная ещё в верхнем палеолите — причём в начале его, о чём свидетельствуют опубликованные в 2008—2013 гг. результаты анализа ДНК ископаемых останков из пещеры Гойе в Бельгии , возраст которых составил 31,7 тыс. лет, и из пещеры Разбойничья на Алтае , у которых возраст оценивается в 33 тыс. лет ). Первоначально она служила сторожевым животным, но скоро стала использоваться и на охоте. К настоящему времени число пород собак , выведенных человеком с самыми различными целями, превысило 350 ; при этом Международная кинологическая федерация признаёт 343 из них . Значительно позднее была одомашнена кошка . Обычно считают, что это сделали носители неолитической культуры Бадари ( Верхний Египет , V тысячелетие до н. э.), использовавшие кошек для защиты собранных ими урожаев ячменя и пшеницы от грызунов . Статус относящихся к более раннему времени находок ( Кипр , Палестина ) остаётся спорным ; нередко эти находки трактуют лишь как свидетельства ранних попыток одомашнивания .

Одомашнивание копытных началась в Передней Азии на рубеже мезолита и неолита и привела к возникновению скотоводства — наряду с земледелием , одной из двух древнейших форм производящего хозяйства . Одомашненные копытные стали основным источником мяса, молока, шерсти и кожи, использовались как транспортные и тягловые животные. Последовательно были одомашнены: овца (стоянка Зави-Чеми Шанидар в северном Ираке , рубеж X—IX тысячелетий до н. э.), коза (стоянка Гандж-Даре в западном Иране , IX тысячелетие до н. э.), корова ( в северной Сирии , VIII тысячелетие до н. э. ), свинья (поселение Чайоню на юго-востоке , конец VIII тысячелетия до н. э.; более ранние находки из Халлан-Чеми и других мест мало отличаются от остатков дикой свиньи , не позволяя говорить именно об одомашнивании, а не простом приручении ). К более позднему времени относится одомашнивание лошади (степи и лесостепи Восточной Европы , VI тысячелетие до н. э.), осла ( Сирия , 1-я половина V тысячелетия до н. э.), двугорбого ( Средняя Азия , 2-я половина V тысячелетия до н. э.) и одногорбого верблюда ( Аравия , IV тысячелетие до н. э.) .

Позже возникли новые очаги одомашнивания. В Андах в конце III — начале II тысячелетия до н. э. одомашнили гуанако (к которому восходит лама , используемая как вьючное животное) и викунью (предок альпака , дающей шерсть); в I тысячелетии до н. э. здесь же была одомашнена морская свинка , ставшая в данном регионе основным источником мяса . В Южной и Юго-Восточной Азии появились свои разновидности крупного рогатого скота: индийский буйвол (предположительно — в III тысячелетии до н. э.), гаял (дикий предок — гаур ) и бантенг . На территории Тибетского нагорья в III тысячелетии до н. э. был одомашнен як . В середине II тысячелетия до н. э. в Египте появилась одомашненная форма лесного хорька — фретка , а в I тысячелетии н. э. во Франции был одомашнен кролик . В Сибири был одомашнен северный олень , но время не вполне ясно: одни авторы датируют начало оленеводства поздним неолитом (III тысячелетие до н. э.), другие — началом железного века (I тысячелетие до н. э.) .
Отметим, что полностью одомашненными принято считать те виды, которые послушны человеку и свободно размножаются в неволе. Так, индийский слон , издавна используемый в странах Южной и Юго-Восточной Азии для выполнения разнообразных работ, не относится к домашним животным, поскольку в неволе он практически не размножается .
Целый ряд видов млекопитающих — мыши , крысы , хомячки , морские свинки , кролики , собаки , различные обезьяны — используются в качестве лабораторных животных при проведении физиологических, генетических и медицинских исследований .
В последние годы значительно выросло разнообразие млекопитающих, играющих роль домашних питомцев . Помимо обычных домашних животных ( собаки , кошки , кролики , морские свинки ), люди держат дома декоративных мышей и крыс , золотистых и карликовых хомячков , шиншилл , ежей и др.
История научного изучения млекопитающих восходит ещё к «отцу зоологии» Аристотелю , который в сочинениях « История животных », « », « О частях животных » привёл сведения о многих видах млекопитающих Средиземноморья , их строении и образе жизни, особенностях размножения и развития млекопитающих. Он не объединял их в единый таксон, а рассматривал в составе двух из пяти выделенных им основных групп «животных с кровью» (то есть позвоночных): «Живородящие четвероногие» и «Живородящие безногие» ( киты ) .
В Средние века интерес к познанию животного мира снизился, но резко возрос с началом Нового времени . Особо способствовали развитию зоологии млекопитающих работы Дж. Рэя и К. Линнея , в которых было положено начало современным представлениям о биологическом виде , системе животного мира и самом классе Mammalia . Большое значение имели работы Ж. Кювье и Э. Жоффруа Сент-Илера , разрабатывавших вопросы типологии и сформулировавших понятие « » организма, а в систематике млекопитающих детализировавших структуру данного класса. Кювье положил начало палеонтологии млекопитающих. Вклад в систематику млекопитающих внесли также Б. Ласепед , К. Иллигер , А. Блэнвиль , Л. Окен . Основы эмбриологии млекопитающих заложили К. Ф. Вольф и К. М. Бэр .

Выход в свет труда Ч. Дарвина « Происхождение видов » (1859) положил начало перестройке зоологии млекопитающих (териологии) на основе представлений эволюционизма . Активную роль в этой перестройке сыграли Т. Гексли , У. Флауэр , Э. Геккель , В. О. Ковалевский , а также представители американской териологической школы, сформировавшейся на базе Американского музея естественной истории : Э. Коп , Г. Осборн , У. Грегори , позднее традиции данной школы продолжали Дж. Г. Симпсон и М. Маккенна . В опубликованных в 1910 году системах млекопитающих Осборна и Грегори уже весьма последовательно выдержаны принципы эволюционной систематики . Основываясь на изучении нервной деятельности млекопитающих, И. М. Сеченов , И. П. Павлов , Ч. С. Шеррингтон заложили основы современной нейрофизиологии .
В XX веке происходит расширение спектра териологических исследований. Активно изучаются вопросы генетики млекопитающих, их этологии , . Результаты териологии находят всё большее применение в прикладных областях ( охотоведение , звероводство , животноводство и др.).
Крупными вехами в области систематики млекопитающих стали появления системы Симпсона (1945), построенной на принципах эволюционной таксономии , и системы Маккенны (1975; позже неоднократно перерабатывалась) — первой макросистемы данного класса кладистического толка . С 1960-х гг. в теоретический арсенал систематики млекопитающих входят методы молекулярной филогенетики . Сначала филогенетические деревья строились на основе анализа свойств белков , а после появления в 1977 году эффективных методов секвенирования ДНК — в основном по анализу нуклеотидных последовательностей . В результате на рубеже XX—XXI веков представления о родственных связях отрядов млекопитающих претерпевают существенные изменения .

В 1996 году на свет появилась овца Долли — первое клонированное млекопитающее. Долли была получена методом терапевтического клонирования : из соматической клетки вымени взрослой овцы извлекли ядро, поместили его в ооцит другой овцы и получившуюся гибридную клетку стимулировали электрошоком, после чего клетка начала делиться. На стадии бластулы её имплантировали в матку третьей овцы, которая и родила Долли . Этот метод применяется для клонирования и других млекопитающих . К настоящему моменту были успешно клонированы такие млекопитающие, как мышь и корова (1998), коза (1999), макак-резус , гаур и свинья (2000), кошка и кролик (2002), лошадь , мул и крыса (2003), собака и бантенг (2005), хорёк (2006), буйвол , волк и олень (2007), верблюд (2010) и др.
В начале XXI века развитие геномики привело к секвенированию геномов некоторых видов млекопитающих, число которых на конец 2013 года составило около сорока. Среди них — человек (2001—2006 гг.) , домовая мышь (2002) , собака (2005) , шимпанзе (2005) , кошка (2007) , домовый опоссум (2007) , утконос (2007) и другие.
| Дерево категорий |
|
Классы
хордовых
(Chordata)
|
|||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Бесчерепные (Acrania) | |||||||||||||||||||||||||
| Olfactores |
|
||||||||||||||||||||||||
|
† —
вымерший
таксон, * —
парафилетический
таксон
|
|||||||||||||||||||||||||