Механизмы запирания и отпирания канала ствола
- 1 year ago
- 0
- 0

Ключевые механизмы старения — типы биохимических изменений, происходящих во всех организмах по мере их биологического старения , которые ведут к постепенной прогрессирующей потере физиологической целостности, нарушениям функционирования и, в конечном итоге, смерти . Впервые они были определены в 2013 году в научной статье «The Hallmarks of Aging» с целью понимания сути биологического старения и обособления его глубинных механизмов . Перевод статьи на русский под названием «Ключевые признаки старения» был опубликован на сайте научно-исследовательского учреждения МЧС России . В 2022 году список механизмов старения было предложено расширить .
Были предложены три ключевых момента, исходя из которых механизмы старения определяются :
Эти условия применимы в различной степени ко всем нижеприведённым механизмам. Последний критерий в отношении многих механизмов является самым труднопроверяемым, поскольку наука ещё не нашла осуществимые на практике способы таких вмешательств в живые организмы.

С течением времени почти все живые организмы имеют постепенные и необратимые возрастные изменения и связанную с этим потерю функций систем организма. Возраст является основным риск-фактором для главных болезней человека, включая рак , диабет , сердечно-сосудистые заболевания и нейродегенеративные болезни . Поэтому важно описать и классифицировать типы изменений, происходящих в организме с возрастом.
В январе 2023 года, через десять лет после публикации оригинальной активно цитируемой научной статьи, её авторы расширили список предложенных механизмов . В статье, закрытой платным доступом , к первоначальным девяти механизмам были добавлены ещё три: нарушенная макроаутофагия , и дисбактериоз .
Учёные классифицировали их в несколько групп.
Эти механизмы однозначно негативны и запускают домино дальнейших повреждений .
Эти механизмы являются частью антагонистических (компенсаторных) ответов на повреждения от первичных механизмов и направлены для минимизации нанесённого вреда. Антагонистические механизмы имеют и положительное, и отрицательное влияние на организм. При слабой активности они полезны. Но при активном длительном задействовании разрегулировываются и начинают превносить вред.
АФК служат важными переносчиками сигналов , но они также вызывают клеточные повреждения. И если повреждения от небольшого количества АФК клетка более-менее успешно ремонтирует и компенсирует, то высокие уровни АФК приводят к увеличению количества повреждений .
Интегральные механизмы проявляются, когда повреждения, вызванные основными и антагонистическими группами повреждений, уже не могут быть скомпенсированы механизмами клеточного и тканевого гомеостаза .
Исправное функционирование генома является наиболее важной предпосылкой для исправного функционирования клетки и организма в целом. Изменения в генетическом коде давно рассматривались как одна из главных причин старения . Генетическая нестабильность происходит от точечных мутаций , делеций , транслокаций , укорочения теломер, одноцепочечных и двухцепочечных разрывов ДНК (с последующим неполноценным ремонтом), хромосомных перестроек , возникновения дефектов в , нарушения генов вследствие вмешательств вирусов и транспозонов . Помимо вклада в старение, геномная нестабильность также играет центральную роль в онкогенезе , а также фактор нейродегенеративных болезней , таких как боковой амиотрофический склероз и нервно-мышечное заболевание .
Мутации накапливаются и в ядерной ДНК, и в митохондриальной , а также появляются дефекты в ядерной ламине .
О темпах мутации ядерной ДНК можно судить по следующему примеру. Обычные клетки эпителия пищевода в организме молодых людей уже содержат сотни мутаций. К среднему возрасту количество мутаций в них уже может превысить 2000 на клетку . Сравнительный анализ показывает, что темп накопления мутаций в клетках различных млекопитающих обратно коррелирует с их продолжительностью жизни .
Аномальные химические структуры в ДНК образуются преимущественно путём окислительного стресса и воздействия факторов окружающей среды . Клетка имеет несколько механизмов репарации ДНК . Но результат восстановления часто не идеален и повреждения постепенно накапливаются. В нескольких научных работах было продемонстрировано, что недостаточный уровень репарации ДНК приводит к большему накоплению повреждений ДНК , что в свою очередь приводит к ускоренному старению. А увеличение репарации ДНК приводит к большей продолжительности жизни .
Мутации в митохондриальной ДНК также вносят свой вклад в процесс старения, вследствие высокого уровня их репликации, ограниченной эффективности их механизмов репарации , окислительного микроокружения и отсутствия гистонов , защищающих эти ДНК-молекулы. Ситуация с исследованием роли мтДНК в старении осложняется тем, что в одной клетке могут одновременно присутствовать исправные и повреждённые мтДНК — состояние, называемое гетероплазмия . Но по крайней мере, глубокое секвенирование старых клеток выявило, что количество мутаций митохондриальной ДНК существенно увеличивается по мере прохождения ими через множество делений . Любопытно, что большинство мутаций в митохондриальной ДНК в зрелых или сенесцентных клетках вызваны не окислительными повреждениями, а ошибками репликации в начале жизни, которые затем множатся и вызывают нарушение дыхательной цепи в разных тканях организма . В исследовании 2022 года ультра-чувствительное секвенирование показало, что большинство мутаций mtDNA в пожилых клетках происходят от репликацонных ошибок, вызванных ДНК-полимеразой γ , а не от окислительного стресса .
Одним из факторов, приводящих к геномной нестабильности, являются возникающие в процессе жизни дефекты в ядерной ламине .

Теломеры — это участки повторяющихся нуклеотидных последовательностей на концах хромосом , которые связываются со специализированными защитными протеинами, называемых шелтеринами или телосомами, с образованием защитной структуры, называемой Т-петлёй. Эта структура защищает хромосомную ДНК от прогрессирующей деградации. Также эта структура не позволяет белкам систем репарации ошибочно принять концы ДНК за концы разорванной хромосомы и случайно сшить две хромосомы вместе.
В процессе репликации в эукариотических клетках ДНК-полимераза не может скопировать крайние участки теломер. Вследствие этого после некоторого количества делений теломеры заметно сокращаются, что сказывается на геномной стабильности и в конечном итоге приводит либо к апоптозу , либо к сенесценции .
Сокращение теломер ассоциировано со старением, смертностью и возрастными болезнями . Влияние сокращения теломер на старение часто изучают на мышах. Эксперименты на генно-модифицированных животных дают основания предположить причинную связь между укорачиванием теломер и старением . Ещё в 1960-х годах Леонард Хейфлик продемонстрировал, что популяция соматических клеток плода человека претерпевает 40-60 делений, после чего клетки входят в сенесцентную фазу . Всякий раз, когда клетка проходит через митоз , теломеры на концах хромосом слегка уменьшаются. Клеточное деление прекращается как только теломеры истощаются до критической длины . Такое положение дел полезно, чтобы остановить неконтролируемую клеточную пролиферацию (как то зарождающийся рак ), но в случае нормально функционирующих клеток это не позволяет им продолжить деление, когда это необходимо.
Существует фермент теломераза , который удлиняет теломеры в гаметах и стволовых клетках . В соматических клетках ген, ответственный за выработку теломеразы, деактивирован как защита от обращения данных клеток в раковые. Низкая активность теломеразы в соответствующих клетках человека приводит к ряду возрастных заболеваний, связанных с потерей регенеративных способностей тканей . Было экспериментально показано, что ускоренное старение в теломераза-дефицитных мышах возвращается к норме, когда теломераза реактивируется . Более того, обычное старение в мышах может быть задержано с помощью фармакологическое активации или трансдукции теломеразы , а мышь со сверхдлинными теломерами демонстрирует удлинённую продолжительность жизни и улучшение метаболического здоровья .

Геном состоит из огромного количества генов, но в данном типе клеток и в данное время только часть из них экспрессируется . Результат функционирования генома зависит как от порядка нуклеотидов в нём, так и от того, какие участки ДНК открыты для транскрипции и какие управляющие участки ДНК регулируют процесс.
При необходимости клетка может включать и выключать гены путями изменений в упаковываний ДНК на разных уровнях и/или с помощью внешних молекулярных меток, устанавливаемых поверх ДНК (к примеру, метилирование ). Последовательность самих нукеотидов в ДНК при этом не меняется. Это уровень регуляции работы генома, осуществляемый без изменений в самом геноме, - эпигеном . Эпигенетический профиль - когда, где и в какой степени изменения происходят - изменяется по мере старения: в основном за счёт накапливающихся поломок в эпигеноме, но возможна и программная составляющая, имеющая целью оптимизациию работы потрёпанной клетки в общем случае. Кроме того, может иметь место ускорение возрастных эпигенетических изменений и ускорение эпигенетических часов под воздействием факторов загрязнённой окружающей среды . В результате данных возрастных изменений полезные гены могут прекратить работать, а ненужные гены включиться, что нарушает нормальное функционирование клетки . Исследование на мышах показало, что на протяжении жизни происходит большой сдвиг в паттерне экспрессии генов в разных тканях .
В создание и поддержание эпигенетических паттернов вовлечены многие системы ферментов : ДНК-метилтрансферазы , ацетилазы гистонов , деацетилазы , метилазы , , а также протеиновые комплексы, вовлечённые в ремоделирование хроматина и в создание некодирующих РНК .
Авторы статьи "Hallmarks of aging: An expanding universe" выделяют следующие разновидности эпигенетических альтераций, вносящих вклад в старение :
С возрастом метилирование одних участков ДНК возрастает, а других уменьшается. Это может приводить к ухудшение экспрессии нужных генов и к началу экспрессии генов, которые должны быть заглушены. Но функциональные последствия изменений для большинства случаев остаются неясными, поскольку эти изменения происходят на интронах и на .
На основе отслеживания метилирования ДНК были созданы ряд эпигенетических часов , позволяющих определять возраст организма и риски смертности. Эти эпигенетические часы позволяют дать оценку эффективности предполагаемых антивозрастных вмешательств в организм . К примеру, в исследовании по геренерации щитовидной железы, продолжавшемся 1 год, эпигенетический возраст испытуемых добровольцев вместо того, чтобы увеличиться на 1 год, уменьшился на 1,5 года. Спустя 6 месяцев после завершения вмешательства их эпигенетический возраст был меньше хронологического на 2 года . Подобным образом, в эксперименте, где добровольцам в течение 7-месяцев давали α-кетоглутарат , их эпигенетический возраст уменьшился на 8 лет .
Учёные отмечают, что изменения в метилировании ДНК ассоциированы с возрастом, однако до сих пор нет твёрдых доказательств, что эти изменения вносят непосредственный вклад в развитие старения. Необходимы дальнейшие исследования, что дефектный паттерн метилирования ДНК ускоряет старение, а улучшение паттерна продлевает жизнь .
С возрастом связаны глобальная потеря гистонов и изменения в посттрансляционной модификации гистонов. Увеличение образования гистонов увеличивает продолжительность жизни дрозофил .
Было проведено множество исследований на модельных животных, показывающих, что воздействие на ферменты, модифицирующие гистоны, приводят к изменению продолжительности и качества жизни . Как пример, сиртуины являются типом ферментов с деацетилазной активностью . Они способствуют связыванию ДНК гистонами и, таким образом, выключают ненужные гены . Эти фермены используют NAD как кофермент . По мере старения уровень NAD в клетках понижается, что приводит к снижению способности сиртуинов в определённое время выключать ненужные гены. Снижение активности сиртуинов ассоциировано с ускоренным старением, а увеличение активности предотвращает несколько возрастных болезней .
Помимо изменений эпигенетических меток непостредственно на ДНК и модификации гистонов, существуют ещё несколько белков и хроматин -ремоделирующих факторов, наподобие (HP1α) и белков группы polycomb , которые вовлечены в геномную стабильность и процессы репарации ДНК и могут влиять на старение. Изменения в этих эпигенетических факторах могут оказать сильное влияние на архитектуру хроматина, включая глобальную потерю гетерохроматина и его перерастпределение, что часто и наблюдается в старых клетках .
Уже проведено много исследований на беспозвоночных по причинно-следственным связям между изменениями хроматина и темпом старения. Эти исследования продемонстрировали, что, к примеру, мутации потери функции в белках HP1α уменьшают продолжительность жизни, а повышенный темп их создания увеличивает продолжительность максимальной жизни и здоровой жизни . Подобных исследований на млекопитающих пока мало, но большинство из них показывает, что расслабление гетерохроматина усиливает развитие старения и возрастных патологий, а поддержание гетерохроматина благоприятствует долголетию .
Эксперименты на модельных животных показали, что многие типы ncRNA способны оказывать влияние на темп старения. К примеру, lncRNA (включая ), microRNA , circRNA . Некодирующие РНК влиют на продолжительность максимальной жизни и здоровой жизни посредством компонентов путей, связанных со старением, и изменяя поведение стволовых клеток
Следствием сбоев в эпигеноме является возможность просыпания в клетке ретротранспозонов и эндогенных ретровирусов.
Ретротранспозоны — это мобильные элементы в геномах, которые при некоторых условиях способны перемещаться из одного места генома в другое или копировать себя, иногда прихватывая с собой часть генетического материала или нарушая работу участка хромосомы, куда они встроились. Для перемещения и копировани ретротранспозоны используют механизмы клетки. Просыпание ретротранспозонов характерно для сенесцентных клеток. Проснувшиеся ретротранспозоны могут активировать пути иммунных механизмов, поскольку могут распознаться иммунной системой как чужеродная ДНК .
Ввод в организм пожилых мышей дикого типа нуклеозидных ингибиторов обратной транскриптазы ( NRTI ), которые подавляют или ослабляют высвобождение ретротранспозонов, уменьшает у них уровень маркеров генетических повреждений . У -дефицитных мышей ввод NRTI продлевает жизнь и улучшает здоровье . Ввод в организм прогероидных мышей антисмысловых олигонуклеотидов ( antisense oligonucleotide ) увеличивает их продолжительность жизни . В геномах некоторых людей-долгожителей обнаружен редкий вариант -гена, который является более сильным ингибитором -ретротранспозонов, повышает геномную стабильность и способен более надёжно уничтожать раковые клетки, чем стандартный SIRT6 .
Эндогенные ретровирусы — это остатки ДНК вирусов, которые когда-то поразили предков организмов. Эти остатки ДНК могут сохраниться в геномах в подавленном состоянии и передаваться из поколения в поколение. При некоторых условиях (к примеру, разрушении ингибирующих их маркеров эндогенома или при сбое в управляющих участках ДНК), эти ретровирусы могут проснуться и внести свой вклад в старение. Они быть подавлены .
— это гомеостазный процесс поддержание пула протеинов, необходимых для нормального функционирования клетки, в надлежащем качестве и в полном объёме . Неправильное свёртывание протеинов , их окисление, неправильное расщепление и неправильная посттрансляционная модификация могут создать нефункционирующие и даже вредные протеины и , которые затрудняют нормальное функционирование клетки и могут стать причиной многих болезней пожилого возраста (таких как БАС , болезнь Альцгеймера , болезнь Паркинсона , катаракта ) . Эксперименты на дрожжах , нематодах и дрозофилах , к ходе которых с помощью генетических манипуляций с рибосомным белком была увеличена аккуратность трансляции , показали увеличение продолжительности жизни у подопытных организмов . А эксперименты на мышах, в ходе которых в рибосомный белок были внесены мутации, увеличивающие количество ошибок при трансляции, привели к преждевременному старению животных .
Некоторые вещества, попадающие в организм извне, способны усугубить нарушение протеостаза и ускорить старение. Так, было зафиксировано уменьшение максимальной жизни и здоровой жизни у дрозофил , которых кормили пищей, обогащённой различными конечными продуктами гликирования ( AGEs ) и липофусцином (агрегация ковалетно -сшитых белков , сахаров и липидов ). При этом в клетках организмов было обнаружено повышенное содержание белков, модифицированных конечными продуктами гликирования, и карбонилированных белков. Все эти негативные вещи ещё более усугубились, когда учёные дополнительно вывели из строя в подопытных дрозофилах лизосомную протеазу .
Отдельные дефектные протеины могут быть разрушены специальными ферментами, такими как протеасома . Протеиновые агрегации могут быть удалены в процессе макроаутофагии путём включения их в аутофагосомы с дальнейшей доставкой в лизосомы для дальнейшего расщепления в них . Но хотя повреждённые протеины постепенно утилизируются и перерабатываются, они успевают навредить клетке. С возрастом количество таких повреждённых белков и их агрегаций увеличивается - происходит нарушение протеостаза . Этот процесс может быть замедлен и подавлен путём уменьшения потребляемых калорий или приёмом рапамицина - оба метода ингибируют сигнальный путь mTOR . Интраназальное введение в мышей рекомбинированного человеческого белка увеличило в них активность протеасом , уменьшило в мозгу уровень липофусцинов , усилило их когнитивные способности и увеличило продолжительность жизни .
Аутофагия — это процесс, в ходе которого клетка отправляет в лизосомы и переваривает в них свои собственные компоненты (такие как белки и органеллы ) для получения энергии, для избавления себя от этих компонентов в случае их сильного повреждения или потому что те оказываются не нужны, а также для избавления себя от проникших патогенов . По мере старения аутофагия нарушается. Скомпрометированная аутофагия наблюдается при ряде старческих состояний, включая нейродегенерацию и . Активация аутофагии может увеличить продолжительность жизни мышей и усилить иммунный ответ на вакцинацию в престарелых людях путём преодоления иммуностарения . Ранее компрометация аутофагии рассматривали как часть механизма нарушения , но поддержание динамической стабильности пула белков и обслуживание органелл являются разными вещи, поэтому было предложено выделить нарушение макроаутофагии в отдельный механизм .
Гены и белки, участвующие в аутофагии, также вовлечены в процессы фагоцитоза и выдавливания из клетки внутриклеточного мусора (к примеру, нефункционированных митохондрий) в форме экзосфер для последующей утилизации их макрофагами .
В одном из исследований долгожителей учёные обследовали CD4 + лимфоциты у 16 потомков людей, имеющих исключительную продолжительность жизни, и обнаружили в этих клетках повышенную активность аутофагии по сравнению со сверстниками из контрольной группы .
Уже проведено много экспериментов на модельных организмах , доказывающих, что стимуляция аутофагии увеличивает в них продолжительности максимальной жизни и здоровой жизни. К примеру, в эксперименте на дрозофилах увеличение аутофагии лишь в клетках энтероцита кишечника увеличивает их продолжительность жизни . В эксперименте на мышах трансгенное увеличение экспрессии под контролем постоянно экспрессируемого промотора достаточно для продления жизни, улучшения метаболического здоровья и моторной функции .
Пероральное введение в мышей усиливает аутофагию во многих органах и увеличивает их продолжительность жизни до 25%, а также замедляет старение сердца . Механизм спермидина, усиливающий аутофагию, связан либо с ингибированием ацетилтрансферазы (что приводит к уменьшению нескольких ключевых белков аутофагии) , либо с модификацией белка (который существенен для синтеза , важного для аутофагии) . EP300 также является мишенью для других веществ, в которых обнаружены жизнепродлевающие свойства, - и салицилата (активное производное соединение аспирина ) . Фармакологическое ингибирование EP300 с помощью синтетического вещества C646 имитирует стимулирующее действие спермидина на аутофагию и иммунное наблюдение за раком . Вышеперечисленные исследования проводились на мышах. Исследователи периодически отмечают схожесть механизмов действия веществ, усиливающих аутофагию, с тем, что происходит при уменьшении потребления калорий , когда тоже наблюдается усиление аутофагии.
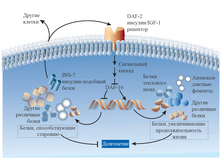
— это способность клетки распознавать изменения в концентрациях поступающих к ней питательных веществ и правильно на это реагировать. Это такие вещества как глюкоза , жирные кислоты , аминокислоты . Во время их изобилия активизируется анаболизм посредством различных сигнальных путей . Наиболее хорошо изученный среди них — сигнальный путь mTOR Когда питательные вещества в дефиците, рецептор AMPK чувствует это и выключает путь mTOR для сбережения ресурсов .
Клеточные системы распознавания веществ мало изменились в процессе эволюции. Они включают внеклеточные лиганды , такие как инсулин и , рецепторные тирозинкиназы , с которыми те взаимодействуют, а также внутриклеточные сигнальные каскады . Эти каскады включают в себя сигнальные пути PI3K-AKT и Ras-MEK-ERK , а также факторы транскрипции , включая и E26, которые гены, вовлечённые в различные клеточные процессы. Комплекс mTORC1 реагирует на питательные вещества, такие как глюкоза и аминокислоты , и стрессорные факторы, такие как гипоксия и нехватка энергии, для регуляции активности множества белков, включая факторы транскрипции наподобие и . Эта сигнальная сеть является центральным регулятором клеточной активности, включающей в себя аутофагию, биогенез mRNA и , синтез белков, метаболизм глюкозы, нуклеотидов и липидов, , активность протеасомы . Активность сигнального пути откликается на наличие питательных веществ и стрессовых факторов, активирую анаболизм , если питательные вещества в изобилии и отсутствует стресс, или активируя клеточные защитные механизмы в ответ на стресс и недостаток питания. Генетически уменьшенная активность компонентов в системах распознавания веществ может увеличить продолжительности максимальной и здоровой жизней в различных модельных организмах . Геномные исследования на людях показали вовлечённость фактора транскрипции FOXO3 и генетических вариантов, кодирующих компоненты систем распознавания веществ, в долголетие людей . Эпигенетический возраст также ассоциирован с внутриклеточным распознаванием веществ . В юности внутриклеточные системы распознавания веществ функционирует таким образом, чтобы способствовать благоприятным анаболитическим процессам, но во взрослом возрасте они приобретают черты, двигающие старение .
В растущем организме важны рост и пролиферация клеток, поэтому путь mTOR высокоактивен. В выросшем организме путь mTOR естественным образом приглушается . Было обнаружено, что принудительная активация этого пути во взрослой мыше ведёт к ускоренному старению и увеличению раковых заболеваний . Методы приглушения пути mTOR, такие как уменьшение потребляемых калорий или приём рапамицина , на сегодняшний день считаются одними из самых надёжных методов увеличения продолжительности жизни червей, мух и мышей.
Если углубиться во влияние диеты на внутриклеточные механизмы распознавание питательных веществ, то переедание
И наоборот, ограничение питания ингибирует mTORC1 и EP300; активирует AMPK, SIRT1 и SIRT3; стимулирует адаптивные клеточные ответы на стрессы. В конечном итоге это удлиняет жизнь у многих модельных организмов вплоть до приматов .
Имеющиеся на данный момент свидетельства указывают на то, что можно добиться увеличения продолжительности жизни подопытных животных путями ингибирования анаболического сигналинга ( ИФР-1 , mTOR , гормон роста ) (к примеру, ограничением в диете или рапамицином), а также активацией катаболического сигналинга ( AMPK , сиртуины ). Различные пути взаимосвязаны: так, активация AMPK ингибирует mTORC1 ( белковый комплекс , каталитическим центром которого является mTOR) . Однако слишком низкий уровень анаболического сигналинга приводит к серьёзным проблемам со здоровьем (нарушение заживления ран, инсулинорезистентность, катаракта, тестикулярная дегенерация) и может привести к преждевременной смерти. Так, прогероидные мыши с очень низким уровнем ИФР-1 могут прожить дольше в случае небольшого дополнительного приёма ИФР-1 .
C возрастом увеличивается активность mTOR в нейронах гипоталамуса мыши, что вносит вклад развитие их возрастного ожирения. Это можно вылечить прямой инфузией рапамицина в гипоталамус .
Парадоксально, но уровни ИФР-1 и гормона роста снижаются естественным образом во время нормального старения, также в мышах с преждевременным старением. Таким образом, снижение активности молекулярного пути ИФР-1 и инсулинового сигналинга является общей характеристикой нормального и ускоренного старения, а постоянно сниженная активность этих процессов увеличивает продолжительность жизни. Эти противоречащие друг другу наблюдения могут быть объяснены тем, что организм снижает активность анаболических путей в ответ на накопившиеся повреждения с целью экономии ресурсов и уменьшения темпа накопления дальнейших повреждений с целью прожить дольше .
Лекарственные вещества, нацеленные на такие болезни как рак и метаболические заболевания , часто изменяют внутриклеточные системы распознавания веществ, поэтому некоторые из этих веществ могут также иметь геропротекторные свойства . Рапамицин и его производные, которые ухудшают работу комплекса mTORC1, продлевают продолжительность жизни различных модельных животных, даже когда ввод их в организм начался в конце взрослой жизни . В мышах рапамицин может улучшить различные аспекты здоровья и защищает организм от некоторых нейродегенеративных и других болезней пожилого возраста , хотя при этом он также может и усугубить отдельные возрастные черты, такие как катаракта . В отношений людей существуют исследования, когда предупреждающий приём ингибиторов mTORC1 пожилыми добровольцами увеличил иммунный ответ их организмов на иммунизацию против гриппа и уменьшил заболеваемость вирусными респитарными заболеваниями последующей зимой .
Нужны дополнительные исследования, чтобы понять, какие пути и в какой степени можно ингибировать и активировать, чтобы получить максимум полезных последствий при минимуме отрицательных .

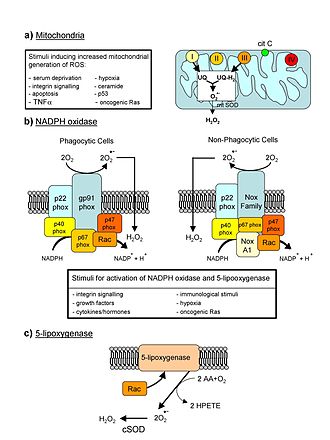
Митохондрия является электростанцией клетки. Различные клетки человеческого тела содержат от нескольких до 2500 митохондрий , каждая из которых конвертирует углерод (в формате Acetyl-CoA ) и кислород в энергию (в формате ATФ ) и диоксид углерода .
С возрастом эффективность митохондрий имеет тенденцию к снижению. Причины этого пока не до конца ясны, но подозреваются несколько механизмов: уменьшение , накопление повреждений и мутаций в митохондриальной ДНК , окисление митохондриальных протеинов, сбои в контроле качества при митофагии .
Неправильно функционирующие митоходрии вносят вклад в старение путём создания помех для внутриклеточных сигналов и запуская воспалительные реакции .
Однако ситуация не всегда однозначна, поскольку митохондриальная дисфункция также связана с гормезисом . Митохондрии являются главным источником образования АФК и дефекты в митохондриях обычно (но не всегда) приводят к повышению образования таких молекул. Но хотя АФК вызывают неконтролируемые повреждения в клетке, они также являются важными сигнальными молекулами. В ряде экспериментов исследователи наблюдали неожиданные вещи, как то
Причина этого в том, что увеличение уровня АФК является триггером для активации механизмов защиты и ремонта, и при некотором уровне АФК положительные последствия от них могут превзойти негативные .
Вещество метформин является слабым митохондриальным ядом, вызывающим низкоэнергетическое состояние, характеризующееся увеличением количества АМФ и активацией AMPK. При соблюдении некоторых условий метформин способен продлевать жизнь различных подопытных животных, в том числе мышей .
Тренировки на выносливость и периодические ограничения в питании могут увеличивать продолжительность здоровой жизни, защищая митохондрии от дегенерации. По-видимому, причина этого в активации аутофагии . Однако в это могут быть вовлечены и другие молекулярные пути, ведущие к долголетию .
Главным внутриклеточным процессом в поддержании качества митохондрий является митофагия , которая предотвращает накопление в клетке дефектных митохондрий. С возрастом митофагия ухудшается, что подстёгивает старение и является серьёзным риск-фактором для многих болезней, включая нейродегенеративные .
При определённых обстоятельствах клетка может прекратить клеточный цикл без умирания, вместо этого перейдя в спящий режим и почти прекратя выполнение своих нормальных функций. Это называется клеточным старением. Данный процесс может быть вызван несколькими факторами, среди которых укорочение теломер , повреждение ДНК и стресс. Иммунная система имеет программу поиска и устранения сенесцентных клеток путём фагоцитоза . В связи с этим есть гипотеза, что переход клеток в сенесцентный режим есть природный механизм для избавления организма от клеток, которые накопили больше повреждений, чем могут исправить системы ремонта. Но по мере ухудшения качества работы иммунной системы та уже перестаёт в полном объёме справлятья с чисткой организма от сенесцентных клеток.
Связи между сенесцентными клетками и старением организма следующие:
Учёные обнаружили, что к увеличению продолжительности жизни могут приводить воздействия на механизм сенесцентности, выглядящие отчасти противоположными. Увеличение продолжительности жизни можно получить и путём лёгкой стимуляцией вызывающих сенесцентрость метаболических путей опухолевой супрессии , и удалением сенесцентных клеток из организма . Были разработаны вещества, способные выборочно уничтожать сенесцентные клетки в организме, - сенолитики . Мыши, которым длительное время давали сенолитики, продемонстрировали более долгую жизнь без увеличения частоты случаев заболевания раком и дефектного ремонта тканей .
Но сенесцентные клетки имеют и положительное влияние на организм, как то способствование заживлению ран . По этой причине учёные предполагают, что чрезмерное удаление сенесцентных клеток из организма хотя и даёт кратковременный положительный эффект, может в дальнейшем привести к ускоренному накоплению сенесцентных клеток и ускорить старение . Необходимы дальнейшие исследования для понимая возможной выгоды и побочных эффектов от терапии сенолитиками .
Стволовые клетки — это недифференцированные или частично дифференцированные клетки, которые имеют способность делиться бесконечно. Эмбрион в возрасте нескольких дней почти полностью состоит из стволовых клеток. По мере роста эмбриона клетки размножаются, дифференцируются и приступают к выполнению своих профильных функций внутри организма. Во взрослом организме стволовые клетки преимущественно сосредоточены в зонах, которые подвергаются интенсивному износу ( желудочно-кишечный тракт , лёгкие , слизистые оболочки , кожа ) или в местах образования клеток, требующих постоянного пополнения ( эритроциты , клетки иммунной системы , сперматозоиды , волосяные фолликулы ).
Потеря способности к регенерации является одним из самых очевидных следствий старения. В основном это происходит из-за того, что со временем количество в организме стволовых клеток и скорость их деления постепенно снижаются . В качестве примера можно привести уменьшение гемопоэза , что, среди прочего, ведёт к уменьшению количества и ухудшению качества иммунных клеток — процессу, называемому , а также к повышенному риску развития анемии и миелоидных злокачественных образований .
Было обнаружено, что омоложение стволовых клеток может обратить вспять некоторые проявления старения на уровне организма . Так, трансплантация стволовых клеток, полученных из молодых мышей, прогероидным мышам увеличивает их продолжительность жизни и уменьшает дегенерацию даже в тех их тканях, в которые донорские клетки не попали. Учёные предполагают, что причиной этого являются секретируемые стволовыми клетками факторы .
Существует исследование на выборке в 16 человек, согласно которому физические упражнения в игровой форме замедляют потерю стволовых клеток в мышцах человека .
Различные ткани и составляющие их клетки нуждаются в тесной координации их работы для функционирования организма как единого целого. Одним из главных способов для этого является выделение сигнальных молекул в межтканевую жидкость и кровь. При достижении других клеток и тканей такие молекулы изменяют поведение тех. Профиль сигнальных молекул изменяется по мере старения.
Появляется всё больше данных, что возраст-зависимые изменения в одной ткани могут вести к нарушениям в других тканях, создавая эффект "заразного старения", также называемый "эффектом свидетеля". Механизмов здесь несколько. К примеру, было обнаружено, что сенесцентные клетки выделяют специфический набор сигнальных молекул, получивший название , который вызывает аналогичное сенесцентное состояние в соседних клетках . Но есть у таких взаимосвязей и обратная, положительная сторона: жизнепродлевающие воздействия, направленные на одну ткань или орган, могут также замедлить старение в других тканях и органах .
Разработаны вещества, способные подавлять действие SASP и, таким образом, замедлять старение. Этот класс веществ получил название сеноморфики .
Существует несколько способов восстановления межклеточных взаимодействий, приводящих к продлению жизни и появлению признаков омоложения. Одним из них является замена в организме некоторых системных факторов крови . Так, одиночное переливание крови старых мышей в молодых мышей приводит к появлению через несколько дней в последних признаков старения , а простое разбавление крови старых мышей буферным изотоническим раствором с 5% содержания альбуминов приводит к появлению признаков омоложения во многих тканях , что свидетельствует о существовании циркулирующих в крови факторов, способствующих процессам старения . Растворимые вещества в крови молодых мышей восстанавливают у старых мышей способность к обновлению и ремонту повреждений .
Старение сопряжено с многочисленными повреждениями в долгоживущих компонентах внеклеточного матрикса , включая образование конечных продуктов гликирования (AGEs), карбонилирование , карбамилирование , фрагментацию эластина и коллагена , что ведёт к фиброзу тканей (фибростарение) . Сковывание внеклеточного матрикса также отражается на сенесцентных клетках, которые в результате секрецируют матриксные металлопротеиназы , увеличивающие повреждения во внутриклеточном матриксе , и посредством протеолиза создают молекулярный паттерн, способствующий возникновению ещё больших повреждений, что активизирует просенесцентные , профиброзные и провоспалительные пути . Важность коллагена для долголетия человека также подчёркнута открытием редкого варианта гена , который кодирует протеин, специфический для коллагена мозга. Этот вид протеина может усиливать защиту от болезни Альцгеймера . Внеклеточный матрикс, приготовленный из фибробластов молодых людей, индуцирует молодое состояние в старых сенесцентных клетках .
Старость коррелирует с высокими уровнями воспалительных активаторов в крови, такими как IL-1 , IL-6 , C-реактивный белок , IFNα и другие . (инфламмэйнджинг) первоначально рассматривалось как часть механизма изменённой межклеточной коммуникации. Однако вследствие его значительного вклада в старение и болезни пожилого возраста , а также ввиду тесных взаимосвязей с клеточным старением и нарушением микробиома , учёные предложили рассматривать старческое воспаление в качестве самостоятельного механизма .
Постепенное изменение профиля межклеточных сигнальных молекул приводит к вялотекущему хроническому воспалению, охватывающему весь организм, получившему название « » . Кратковременное воспаление выполняет в организме положительную роль, в случае возникновения повреждения рекрутируя компоненты иммунной системы и механизмов ремонта в повреждённую часть организма настолько долго, насколько повреждение и угроза сохраняются. Но постоянное присутствие воспалительных маркеров по всему телу изнашивает иммунную систему и травмирует здоровые ткани .
Инфламмэйджинг является результатом многих процессов, главные из которых
Эти изменения приводят к повышенной активации многих провоспалительных путей, что приводит к повышенному образованию IL-1b , фактора некроза опухоли и интерферонов . Инфламмэйджинг также способствует патогенезу ожирения , диабета 2 типа , атеросклероза , остеоартрозу , . Также инфламмэйджинг подавляет функции эпидермальных стволовых клеток (см. " ") и понижает эффективность иммунной системы в распознавании и удалении сенесцетных клеток (см. " "), что является примерами связи различных механизмов старения между собой .
Проведено множество исследований, демонстрирующих увеличение продолжительности максимальной жизни и здоровой жизни в результате использования противовоспалительных терапий. Так,
Физические упражнения и пониженный индекс массы тела ведут к уменьшению количества воспалительных маркеров в плазме крови . Воспалительные маркеры также можно понизить через здоровое питание. В частности, к понижению воспалительных маркеров ведут потребление оливкового масла , орехов . СРБ понижают кофе , витамина D (возможно, вследствие высокого уровня недостатка витамина D в людях современного общества), глюкозамин и хондроитин . Другие потенциальные противовоспалительные вещества, имеющие пользу для здоровья: , кверцетин , кофермент Q (Q 10 ), имбирь , ресвератрол , магний , карнозин , мелатонин , куркумин , , телмисартан , силденафил , пентоксифиллин , а также пробиотики , пребиотики и симбиотики .
С возрастом в микробиоме кишечника происходят заметные изменения, в осносном сдвиг в численности различных микроорганизмов и уменьшение их разнообразия . В связке с потерей структурной целостности кишечника и других барьеров (как то гематоэнцефалический барьер ) эти изменения в популяциях микроорганизмов могут способствовать .
Учитывая влияние кишечного микробиома на организм хозяина, можно сделать предположение о возможности увеличения продолжительности и качества жизни посредством измения этого микробиома .
Исследования последних лет продемонстрировали, что микробиом является важным фактором для многих физиологических процессов, таких как переваривание и усвоение питатальных веществ, защита от патогенов , синтез в организме существенных метаболитов , включая витамины , аминокислоты , вторичные жёлчные кислоты, короткоцепочечные жирные кислоты . Микробиота кишечника также посылает сигналы периферической и центральной нервным системам и другим отдалённым органам, тем самым оказывая сильное влияние на общее здоровье организма хозяина . Нарушения в двухнаправленной бактерии-хозяин коммуникации приводят к дисбактериозу и вносит вклад в возникновение различных патологических состояний, таких как ожирение , диабет 2-го типа , язвенный колит , , сердечно-сосудистые заболевания и рак .
Сообщество микроорганизмов внутри пищеварительного тракта сильно различается от человека к человеку как следствие генетических различий людей (в первую очередь этничности ), диеты, привычек образа жизни (культура), окружащей обстановки (география и экология). Это делает трудным выявление взаимосвязей между микробиотой и возрастными изменениями. И всё же некоторые мета-анализы выявили ассоциации между микробиотой и болезнями, которые были подтверждены для различных патологий и стран .
Бактериальное разнообразие закладывается в детстве и остаётся относительно стабильным в течение взрослого периода. Однако его структура и активность с возрастом всё же постепенно меняется, постепенно приводя к общему уменьшению разнообразия. Так, несколько исследований, проведённых на полуляциях долгожителей, показали уменьшение основных таксонов бактерий, таких как бактероиды и , но при этом увеличение некоторых родов бактерий, таких как бифидобактерии и - что, вероятно, благотворно сказывается на продолжительности жизни .
Другое исследование показало, что здоровое долголетие ассоциировано с уменьшением основных таксонов, таких как бактероиды , присущих большинству людей. Более того, среди индивидуалов, достигших большого возраста, сохранение высокого уровня бактероидов и низкий уровень уникальности микробиоты существенно ассоциированы с уменьшением вероятности выживания . С другой стороны, исследование ELDERMET (в Ирландии ) рапортует об увеличении преобладания бактерий родов бактероиды , и в людях с возрастом более 65 лет по сравнению с молодыми . Это исследование также обнаружило возрастные сдвиги в микробиоте, ассоциированные с хрупкостью, познанием, депрессией и воспалением .
Различные исследования рапортуют различные закономерности, происходящие в микробиоте людей при старении. Гетерогенность данных показывает, что могут существовать различные траектории изменения микробиома с возрастом. Но при этом учёные обнаружили интересную схожесть в производных соединениях аминокислот , которые микробиота создаёт в плазме крови .
Анализ микробиомы столетних жителей показал высокое содержание в них некоторых определённых бактерий, таких как Alistipes putredinis и Odoribacter splanchnicus . Некоторые из этих видов бактерий способны производить уникальные вторичные жёлчные кислоты , которые демонстрируют мощный антимикробный эффект против грамположительных бактерий , имеющих устойчивость ко многим лекарствам (это такие бактерии как Clostridioides difficile и Enterococcus faecium ) .
Разносторонние исследования моделей ускоренного старения показало, что два типа прогероидных мышей имеют кишечный дисбактериоз, характеризующийся увеличенным изобилием протеобактерий и цианобактерий , а также пониженным уровнем Verrucomicrobia . Трансплантация фекальной микробиоты (ТФМ) от мышей дикого типа этим прогероидным мышам увеличивавет продолжительности их максимальной и здоровой жизней. Этот же эффект можно получить введением verrucomicrobium . И противоположно, ТФМ от прогероидных мышей мышам дикого типа провоцирует пагубные метаболические изменения. Восстановление уровня вторичных жёлчных кислот и других метаболитов , нехватка которых наблюдается у прогероидных мышей, фенокопирует благоприятные эффекты восстановления здорового микробиома .
ТФМ также выявила причинную роль дисбактериоза кишечника в и спаде приобретённого иммунитета , ассоциированных с возрастом и возрастными болезнями. Трансплантация кишечной микробиоты от старых мышей молодым вызывает воспалительную реакцию, характеризующуюся увеличенным уровнем дифференцировки CD4+ T-лимфоцитов в селезёнке , активизацией воспалительных цитокинов и усилением циркуляции воспалительных факторов бактериального происхождения . ТФМ также предоставляет свидетельства причастности микробиомы кишечника к поддержанию здоровья мозга и иммунитета в процессе старения .
В мышах модели ускоренного старения SAMP8, пробиотическая бактерия GKM3 способствует долголетию и смягчает возрастное когнитивное ухудшение . В случае старых мышей и макак , хроническое воспаление, вызванное кишечной микробиотой, и происходящее вследствие него усиление инсулинорезистентности могут быть исправлены восстановлением изобилия благоприятных КЦЖК -производящих бактерий, таких как .
Вышеприведённые и другие исследования показывают наличие причинно-следственной связи между дисбактериозом и скоростью-качеством протекания старения. Исследования дают основание полагать, что восстановление юношеской микробиоты может увеличить продолжительности максимальной и здоровой жизней .
Дисбактериоз кишечника был признан фактором риска развития болезни Альцгеймера .
В марте 2022 в Копенгагене состоялся исследовательский симпозиум "New Hallmarks of Ageing". По результатам обсуждений на нём, большая группа учёных в августе 2022 года предложила расширить существовавший на тот момент классический список девяти механизмов ещё пятью :
Выделить нарушение аутофагии органел в отдельный механизм было предложено авторами коненгагенского списка в августе 2022 и поддержано авторами оригинального списки механизмов старения в январе 2023.
Сплайсинг РНК является частью процессинга РНК - процесса создания ДНК из мРНК . В исследовании на большом количестве людей было замечено, что по мере старения качество этого процесса ухудшается . Разрегулировка сплайсинга РНК не относится ни к механизму нестабильности генома (который об изменениях в ДНК), ни к механизму эпигенетических альтераций (который об изменениях эпигенетических меток поверх ДНК) .
Дополнить список ключевых механизмов старения нарушением микробиома было предложен авторами коненгагенского списка в августе 2022. Предложение было поддержано авторами оригинального списки в январе 2023.
Учёные предлагают рассматривать в рамках единого механизма как имежклеточные изменения, так и внутриклеточные .
По мере процесса жизнедеятельности во внеклеточном матриксе между молекулами коллагена вследствие гликирования образуются и накапливаваются . Из-за этого с годами постепенно теряется тканевая эластичность и изменяется поведение клеток .
Примером внутриклеточного изменения маханических свойств являются изменения в ядерной ламине — фибриллярной сетке внутри ядра, определяющей его механические свойства, участвующей в организации хроматина и ряде важных процессов. Нарушения в ядерной ламине являются ключевой особенностью прогерии — болезни, во многих чертах напоминающей преждевременное старение. Клинические испытание показали, что лекарственное вещество, восстанавливающее целостность ядерной мембраны , увеличивает продолжительность жизни людей, больных прогерией .
Возрастные механические изменения тканей препятствуют мобильности фибробластов и иммунных клеток .
Для решения этого класса проблем с целью омоложения организма перспективной выглядит направление .
Выделить старческое хроническое воспаление в отдельный механизм было предложено авторами коненгагенского списка в августе 2022 и поддержано авторами оригинального списки механизмов старения в январе 2023.
Учёные подчёркивают, что не исключено существование других механизмов, способных вносить вклад в постепенное развитие в организме старения .
Помимо подхода, что этот процесс обусловлен постепенным накоплением различных случайных поломок и разбалансировок, есть группа квазипрограммых теорий старений, согласно которым существуют механизмы, которые в некоторый период жизни ускоряют скорость старения .

В 2014 году другая группа учёных предложила несколько иную концептуальную модель, названную "Семь столпов старения" . Модель семи столпов подчёркивает взаимные взамосвязи между всеми выделенными в ней механизмами, что в явном виде не указывалось в работе "Hallmarks of aging" .
Подход SENS к старению, разработанный Обри ди Греем , предполагает, что существует 7 типов повреждений, исправление которых ликвидирует старение. Согласно этому подходу, все остальные возрастные изменения являются лишь следствием. По словам Обри ди Грея, существование 8-го типа повреждений возможно, но крайне маловероятно .
Авторы статьи "Hallmarks of aging: An expanding universe" также связали различные ключевые механизмы старения с .
Они заключили, что ключевые механизмы старения не только взаимосвязаны между собой, но также связаны с недавно предложенными " ", которые включают организационные особенности пространственной компартментализации, поддержание гомеостаза и адекватные реакции на стресс .
Научная статья "The hallmarks of aging" считается фундаментальной работой в области старения с момента публикации и предопределила направление многих исследований. Согласно Google Scholar , она цитируется более 1000 раз за год. Однако критики отмечают, что статья имеет ряд слабых мест и является скорее обзором различных аспектов старения, а не истинной парадигмой старения .
В частности, отмечаются следующие проблемные места:
Поэтому необходима дальнейшая работа, и чрезмерное зацикливание на статье может помешать созданию истинной парадигмы старения .
В своей последующей работе 2023 года “Hallmarks of aging: An expanding universe” авторы частично учли часть критики. В особенности, было уделено большое внимание на демонстрацию взаимосвязей между различными механизмами старения. Отмечается, что отличительной чертой эффективных анти-возрастных вмешательств (таких как метформин , , предшественники NAD+ , пероральный приём ) является то, что эффекты от их приёма проявляются одновременно в нескольких механизмах . Но большая часть критики по-прежнему актуальна.
Общий перечень ключевых механизмов старения стал полезным контрольным списком для исследователй, изучающих отдельные болезни старения и пытающихся разобраться в их причинах . Он побудил создание подобных перечней ключевых механизмов старения для отдельных органов и систем организма. К примеру, по старению лёгких , клеточному старению , экспрессии генов в стареющих клетках , старению мозга .
{{
cite journal
}}
:
Проверьте значение даты:
|date=
(
справка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
:
Cite journal требует
|journal=
(
справка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (формат даты) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
:
Проверьте значение даты:
|date=
(
справка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
:
|last14=
имеет числовое имя (
справка
)
| Вопросы |

|
|
|---|---|---|
| Журналы | ||
| Ресурсы | ||
| Люди | ||
| Организации | ||
|
Категория «
Продление жизни
»
|
||