Interested Article - Почка млекопитающего

- 2020-08-29
- 1
Почка млекопитающего — парный орган мочевыделительной системы млекопитающих , являющийся разновидностью метанефрической почки . Почка у млекопитающих обычно бобовидной формы , расположена забрюшинно на задней (дорсальной) стенке тела . Каждая почка состоит из фиброзной оболочки , периферического коркового вещества, внутреннего мозгового вещества, а также чашек и почечной лоханки, однако чашечки или почечная лоханка у отдельных видов могут отсутствовать. Выводится моча из почки через мочеточник . Строение почки может различаться между видами в зависимости от среды обитания, в частности от её засушливости . За фильтрацию крови ответственно корковое вещество, по своей сути оно аналогично типичным почкам менее развитых позвоночных . Азотсодержащие продукты жизнедеятельности выделяются почками у млекопитающих преимущественно в виде мочевины .
В зависимости от вида почки могут быть однодолевыми или многодолевыми, однососочковыми, с несколькими сосочками или многососочковыми , могут быть с гладкой поверхностью или бороздчатыми , также почки могут быть множественными, множественные встречаются в основном у морских млекопитающих . Наиболее простым типом почки у млекопитающих является однососочковая однодолевая почка . Различия в структуре почек являются результатом адаптации в ходе эволюции к различной массе тела и среде обитания . К почкам млекопитающих относятся почки человека .
Корковое и мозговое вещество почек содержит в себе нефроны . У млекопитающих нефроны разделяются на нефроны с короткой и с длинной петлёй Генле , ответственной за концентрацию мочи . Среди позвоночных вырабатывать концентрированную мочу могут только почки млекопитающих и птиц , но лишь у млекопитающих петлю Генле содержат все нефроны .
Почки млекопитающих являются жизненно-важными органами , которые поддерживают водно-электролитный и кислотно-щелочной баланс в организме, выводят азотистые продукты жизнедеятельности, регулируют артериальное давление , участвуют в формировании костей и в поддержании гомеостаза глюкозы . В почках происходят процессы фильтрации плазмы крови, канальцевой реабсорбции и канальцевой секреции, в результате этих процессов формируется моча . Из числа гормонов в почках вырабатываются ренин и эритропоэтин , также почки участвуют в преобразовании витамина D в его активную форму . При этом млекопитающие являются единственным классом позвоночных, у которого за поддержание гомеостаза внеклеточной жидкости в организме отвечают одни лишь почки . Регулируется деятельность почек вегетативной нервной системой и гормонами .
Почки могут быть подвержены неинфекционным и инфекционным заболеваниям, в редких случаях в почках млекопитающих встречаются врождённые и наследственные аномалии . Причиной пиелонефрита обычно являются бактериальные инфекции . Некоторые заболевания могут быть специфичными для видов , у некоторых видов распространены паразитарные заболевания почек . В силу особенностей устройства у млекопитающих почки склонны к ишемическим и токсическим повреждениям . Возможности регенерации в зрелых почках ограничены , поскольку новые нефроны не образуются , однако в случае ограниченных повреждений функция почек может быть восстановлена за счёт компенсационных механизмов . Постоянные повреждения могут приводить к хронической болезни почек . В процессе старения почки также претерпевают изменения, в них снижается количество функционирующих нефронов .
Структура
Расположение и форма

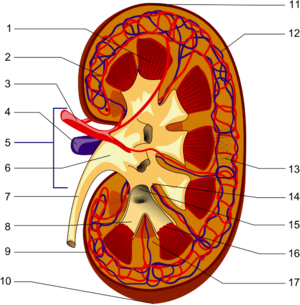
- 1. Фиброзная капсула
- 2. Корковое вещество
- 3. Почечная пирамида мозгового вещества
- 4. Почечный столб коркового вещества
- 5. Нефрон
- 6. Почечный сосочек
- 7. Малая почечная чашка
- 8. Большая почечная чашка
- 9. Почечная лоханка
- 10. Мочеточник
- 11. Почечная артерия
- 12. Почечная вена
- 13. Междолевая артерия
- 14. Почечная доля
- 15. Дуговая артерия
- 16. Междольковая артерия
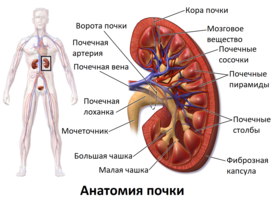
У млекопитающих почки обычно бобовидной формы , такая форма свойственна только млекопитающим . Располагаются забрюшинно на задней (дорсальной) стенке тела . Одним из ключевых факторов, которые определяют форму и морфологию почек у млекопитающих является их масса . Вогнутая часть бобовидных почек называется почечными воротами. В них в почку входят почечная артерия и нервы , а выходят почечная вена, лимфатические сосуды и мочеточник . Снаружи почка окружена массой жировой ткани .
Общее строение
Внешний слой каждой из почек состоит из фиброзной оболочки, называемой капсулой. Периферический слой почки представлен корковым веществом, а внутренний — мозговым. Мозговое вещество состоит из пирамид, восходящих своим основанием к корковому веществу и образующих вместе с ним почечную долю . Пирамиды между собой разграничиваются почечными столбами ( столбами Бертена ), образованными корковой тканью . Вершины пирамид оканчиваются почечными сосочками, из которых моча выводится в чашечки, в лоханку, мочеточник и мочевой пузырь , после чего она выводится наружу через мочеиспускательный канал .
Паренхима

- 1. Корковое вещество
- 2. Наружная зона мозгового вещества
- 3. Внутренняя зона мозгового вещества
- 4. Внутренняя полоса наружной зоны мозгового вещества
- 5. Наружная полоса наружной зоны мозгового вещества
- 6. Юкстамедуллярный нефрон
- 7. Мозговой луч
- 8. Почечная капсула
- 9. Почечный сосочек.
Подоциты Проксимальный извитой каналец Проксимальный прямой каналец Промежуточный каналец (петля Генле) Дистальный прямой каналец Macula densa (плотное пятно) Дистальный извитой каналец Соединительный каналец Собирательна трубочка
Паренхима , будучи функциональной частью почек, визуально делится на корковое и мозговое вещество . В основе коркового и мозгового вещества лежат нефроны в совокупности с разветвлённой сетью кровеносных сосудов и капилляров, а также собирательные трубочки, в которые нефроны впадают, собирательные протоки и почечный интерстиций . В корковом веществе расположена фильтрующая кровь часть нефрона — почечное тельце, от которого внутрь спускается почечный каналец, переходящий в мозговом веществе в петлю Генле , затем каналец возвращается назад в корковое вещество и своим дистальным концом впадает в общую для нескольких нефронов собирательную трубочку. Собирательные трубочки спускаются снова в мозговое вещество и объединяются в собирательные протоки, проходящие через внутреннее мозговое вещество .
По расположению почечных клубочков нефроны делятся на 3 типа: субкапсулярные (ближе к капсуле), интраконтикальные (в средней части) и юкстамедуллярные (ближе к мозговому веществу). По длине петли Генле — на нефроны с длинной петлёй и с короткой петлёй Генле . Соотношение коркового к мозговому веществу варьируется между видами, у одомашненных животных корковое вещество обычно занимает третью или четвёртую часть толщины паренхимы, в то время как у пустынных животных с длинными петлями Генле — лишь пятую часть .
Корковое вещество
Структурно корковое вещество состоит из коркового лабиринта и мозговых лучей . Корковый лабиринт содержит в себе междольковые артерии, сосудистые сети, образованные афферентными и эфферентными артериолами , почечные тельца, проксимальные извитые канальцы, плотные пятна ( лат. macula densa ) , дистальные извитые канальцы, соединительные канальцы и начальные части собирательных трубочек . Преобладают в корковом лабиринте проксимальные извитые канальцы . Сплошной слой коркового вещества, лежащий над мозговыми лучами, называется корой коркового вещества ( лат. cortex corticis ) . У некоторых млекопитающих есть нефроны, петли Генли которых не достигают мозгового вещества, такие нефроны называются корковыми . Мозговые лучи коркового вещества содержат в себе проксимальные прямые канальцы, корковую часть восходящих толстых ветвей петли Генле и корковую часть собирательных трубочек . При этом корковое вещество делится на дольки, каждая из которых представляет из себя мозговой луч в совокупности с ассоциированными с ним нефронами, а между дольками проходят междольковые артерии .
Мозговое вещество
Мозговое вещество у млекопитающих делится на наружную и внутреннюю зоны. Наружная зона состоит из коротких петлей Генле и собирательных трубочек, внутренняя — из длинных петлей и собирательных протоков . Наружная зона, в свою очередь, также подразделяется на наружную (лежащую прямо под корковым веществом) и внутреннюю полосы . Отличаются полосы тем, что в наружной присутствуют проксимальные прямые канальцы, а во внутренней — тонкие нисходящие части петли Генле (отдел нефрона, следующий за проксимальным прямым канальцем) .
Соотношение наружного и внутреннего мозгового вещества
У большинства видов есть нефроны и с короткими, и с длинными петлями, у отдельных же видов может быть лишь один тип. Например, у горных бобров есть только нефроны с короткой петлёй, и, соответственно, отсутствует внутреннее мозговое вещество. У собак и кошек, наоборот, есть только нефроны с длинными петлями. Соотношение нефронов с короткими петлями Генле к нефронам с длинными также варьируется между видами .
Структурные различия между видами

Структурно почки варьируются между млекопитающими . То, какой структурный тип будет у того или иного вида, в основном зависит от массы тела животных . У маленьких млекопитающих встречаются простые однодолевые почки с компактной структурой и одним почечным сосочком , в то время как у больших животных — почки многодолевые, как например, у крупного рогатого скота , при этом у крупного рогатого скота почки бороздчатые, визуально поделённые на доли . Сама по себе доля равноценна простой однососочковой почке, как у крыс или мышей . У крупного рогатого скота также отсутствует почечная лоханка, моча из больших чашек выводится напрямую в мочеточник .
По количеству почечных сосочков почки могут быть однососочковыми , как например у крыс и мышей , с несколькими почечными сосочками, как у паукообразных обезьян , или с большим количеством, как у свиней и человека . У большинства животных один почечный сосочек . У некоторых животных, например, у лошадей , окончания почечных пирамид сливаются друг с другом с образованием общего почечного сосочка, называемого почечным гребнем . Почечный гребень обычно появляется у животных, размером больше кролика .

- 1. Почечная артерия
- 2. Почечная вена
- 3. Общий собирательный проток, переходящий в мочеточник.
У морских млекопитающих , выдр и медведей почки множественные, состоящие из маленьких почечек , каждая из которых аналогична простой однодолевой почке . Почки морских млекопитающих могут состоять из сотен или тысяч почечек, у каждой из которых свои корковое и мозговое вещество и чашечка . Например, у китов порядка 7000 объединённых общей собирательной системой почечек . У ламантин , тоже являющихся морскими млекопитающими, фактически почки многодолевые, поскольку корковое вещество сплошное .
Размеры почек увеличиваются с массой млекопитающих, а количество нефронов в почках между млекопитающими возрастает алометрически . У мышей почки длиной примерно 1 см , массой 400 мг , c 16 000 нефронов, в то время как у косатки длина почки превышает 25 см , масса составляет примерно 4,5 кг , с количеством нефронов порядка 10 000 000. При этом почки косаток множественные, а каждая почечка сравнима с почкой мышей (длина почечки составляет 1 см , масса — примерно 430 мг ) .
Микроанатомия
С точки зрения микроанатомии почку структурно можно разделить на несколько основных элементов: почечные тельца, канальцы, интерстиций и сосудистую сеть . Интерстиций представляет собой клетки и внеклеточный матрикс в пространстве между клубочками, сосудами, канальцами и трубочками . Пространство интерстиция, окружающее клетки, заполнено тканевой жидкостью . Интерстиций между канальцами содержит в себе дендритные клетки , макрофаги , лимфоциты и фибробласты . В корковом веществе он включает в себя эндотелиальные клетки лимфатических капилляров , которые считаются частью интерстиция из-за отсутствия базальной мембраны . Интерстициальные фибробласты формируют каркас тканей почки . Через интерстиций проходят кровеносные сосуды, нервы и лимфатические сосуды . Каждый нефрон с продолжающей его собирательной трубочкой и снабжающая нефрон сосудистая сеть встроены в интерстиций. Нефрон вместе с продолжающей его собирательной трубочкой называется мочевым канальцем ( англ. uriniferous tubule ) .
В почках млекопитающих описаны порядка 18—26 различных типов клеток, при этом большой разброс диапазона обусловлен отсутствием консенсуса по тому, что считать отдельным видом клеток, и вероятно, межвидовыми различиями . Почечные тельца состоят из 4 типов клеток: фенестрированного эндотелия , мезангиальных клеток, подоцитов и . По крайней мере из 16 различных типов клеток состоят почечные канальцы . Сами канальцы поделены на по крайней мере 14 сегментов , которые различаются типами клеток и функциями . Нормальное функционирование почек обеспечивается всей совокупностью эпителиальных , эндотелиальных , интерстициальных и иммунных клеток .
Кровоснабжение

Кровь поступает в почку через почечную артерию , которая в многодолевой почке затем разветвляется в области почечной лоханки на крупные междолевые артерии, проходящие по почечным столбам . Междолевые артерии, в свою очередь, ветвятся у основания пирамиды, давая начало дуговым, от которых в корковое вещество отходят междольковые артерии . Междолевые артерии снабжают пирамиды и прилегающее корковое вещество разветвлённой сетью кровеносных сосудов . Само корковое вещество сильно пронизано артериями, в то время как в мозговом веществе артерии отсутствуют . Венозный отток крови идёт обратно параллельно артериям . У некоторых видов в корковом веществе под капсулой формируются сплетения из обособленных от артерий вен, которые у человека называются звёздчатыми, впадают эти вены в междольковые вены . у млекопитающих отсутствует , за исключением однопроходных . Млекопитающие являются единственным классом позвоночных (за исключением некоторых видов), у которых нет почечно-портальной системы .
Сосудистые клубочки нефронов получают кровь от афферентных артериол, которые, в свою очередь, берут начало в междольковых артериях с промежуточным формированием преартериол. От каждой афферентной артериолы отходит несколько почечных клубочков. Затем эти клубочки переходят в эфферентную артериолу, в которую от нефронов поступает отфильтрованная кровь. У нефронов с длинной петлёй Генле эфферентные артериолы разветвляются, формируя ( лат. vasa recta ) , нисходящие в мозговое вещество. Восходящие прямые сосуды, нисходящие прямые сосуды и петля Генле в совокупности формируют противоточную систему почки. В афферентную артериолу кровь подаётся под высоким давлением, что способствует фильтрации, а в эфферентной она оказывается под низким давлением, что способствует реабсорбции .
Несмотря на небольшие размеры, на почки млекопитающих приходится значимая часть минутного объёма кровообращения . Считается, что у сухопутных млекопитающих через почки проходит примерно пятая часть объёма крови, который проходит через сердце . У взрослых мышей , например, этот показатель составляет 9 %—22 % .
Лимфатическая система
Почка достаточно хорошо снабжена лимфатическими сосудами , которые удаляют из заполняющего пространство между канальцами и кровеносными сосудами интерстиция излишнюю жидкость, растворённые в ней вещества и макромолекулы . Анатомия лимфатический системы почки между млекопитающими схожа . Лимфатические сосуды в основном повторяют путь кровеносных .
Начинается лимфатическая система почек в корковом веществе с начальных внутридольковых лимфатических капилляров, проходящих вблизи канальцев и почечных телец, но при этом лимфатические сосуды не заходят внутрь почечных телец. Далее внутридольковые лимфатические капилляры соединяются с дуговыми лимфатическими сосудами . Дуговые переходят в междолевые, которые проходят вблизи междолевых артерий . Дуговые и междолевые лимфатические сосуды являются лимфатическими преколлекторами . Наконец, междолевые переходят в собирательные лимфатические сосуды почечных ворот, выходящие из почки . В мозговом веществе лимфатические сосуды у млекопитающих обычно не присутствуют, а роль лимфатических сосудов берут на себя ( лат. vasa recta ) .
У отдельных видов могут быть отличия в анатомии лимфатической системы почки. Например, у овец отсутствуют лимфатические сосуды в почечной капсуле, а у кроликов отсутствуют междольковые лимфатические сосуды . Что касается мозгового вещества, в большинстве исследований не удаётся обнаружить лимфатические сосуды в мозговом веществе почек животных, в частности, они не обнаружены у овец и крыс . Отдельные исследования обнаружили лимфатические сосуды в мозговом веществе почек свиней и кроликов . В зависимости от вида может также быть или не быть соединение между лимфатическими сосудами почечной капсулы и почечной лимфатической системой .
Снабжение нервами
Иннервация почки обеспечивается входящими в почку через почечные ворота эфферентными симпатическими нервными волокнами , берущими своё начало в солнечном сплетении , и афферентными, выходящими из почки к спинальному ганглию . Достоверных доказательств иннервации почки парасимпатическими нервами нет , существующие же свидетельства являются спорными . Эфферентные симпатические нервные волокна достигают сосудистой системы почки, почечных канальцев, юкстагломерулярных клеток и стенки почечной лоханки , при этом все части нефрона иннервируются симпатическими нервами . Проходят нервные волокна внутри соединительной ткани , расположенной вокруг артерий и артериол . В мозговом веществе нисходящие прямые сосуды ( лат. vasa recta ) иннервируются до тех пор, пока они содержат в себе гладкомышечные клетки . Большинство афферентных нервных волокон расположены в области почечной лоханки . Подавляющая часть нервов в почках являются немиелинизированными .
Нормальная физиологическая стимуляции эфферентных симпатических нервов почки участвует в поддержании баланса воды и натрия в организме. Активация эфферентных симпатических нервов почки снижает в ней кровоток, соответственно, фильтрацию и выведение с мочой натрия, а также увеличивает скорость секреции ренина . Афферентные нервы в почке также участвуют в поддержании баланса. Механочувствительные нервы почки активируются растяжением ткани почечной лоханки, что может произойти при увеличении скорости потока мочи из почки, в результате чего рефлекторно снижается активность эфферентных симпатических нервов. То есть активация афферентных нервов в почке подавляет активность эфферентных .
Функции
Выделительная функция
У млекопитающих азотистые продукты метаболизма выводятся преимущественно в форме мочевины , которая является конечным продуктом метаболизма млекопитающих и хорошо растворима в воде . Мочевина образуется преимущественно в печени в качестве побочного продукта метаболизма белков . Большая часть мочевины выводится именно почками . Фильтрация крови, как и у других позвоночных происходит в почечных клубочках, где кровь под давлением проходит через проницаемый барьер, который отфильтровывает клетки крови и большие белковые молекулы, образуя первичную мочу. Отфильтрованная первичная моча осмотически и по содержанию ионов такая же, что и плазма крови. В канальцах нефрона происходит последующее повторное всасывание полезных для организма веществ, растворённых в первичной моче, и концентрация мочи .
Осморегуляция
Почки млекопитающих поддерживают почти неизменный уровень осмолярности плазмы крови. Основной составляющей плазмы крови, определяющей её осмолярность являются натрий и его анионы . Ключевую роль в поддержании постоянного уровня осмолярности играет контроль соотношения натрия и воды в крови . Потребление большого количества воды способно разбавить плазму крови, в этом случае почки вырабатывают более разбавленную мочу по сравнению с плазмой, чтобы оставить соль в крови, но вывести излишки воды. Если же воды потребляется слишком мало, то моча выводится более концентрированной, чем плазма крови . Концентрация мочи обеспечивается осмотическим градиентом, который увеличивается от границы между корковым и мозговым веществом до вершины мозговой пирамиды .
Помимо почек в регулировании баланса воды участвуют гипоталамус и нейрогипофис посредством системы обратной связи. Осморецепторы гипоталамуса реагируют на повышение осмолярности плазмы крови, в результате чего стимулируется секреция вазопрессина задней долей гипофиса, а также возникает жажда . Почки посредством рецепторов реагируют на увеличение уровня вазопрессина повышением реабсорбции воды, в результате чего осмолярность плазмы снижается за счёт её разбавления водой .
Варьирование количества выводимой воды является важной в плане выживания функцией для млекопитающих, у которых доступ к воде ограничен . Особенностью почек млекопитающих являются петли Генле, они представляют собой наиболее эффективный способ реабсорбции воды и создания концентрированной мочи, что позволяет сохранять воду в организме . После прохождения петли Генле жидкость становится гипертонической по отношению к плазме крови . Почки млекопитающих сочетают в себе нефроны с короткой и с длинной петлёй Генле . Способность концентрации мочи определяется главным образом структурой мозгового вещества и длиной петель Генле .
Эндокринная функция
Помимо выделительной, почки также выполняют эндокриную функцию, то есть вырабатывают некоторые гормоны . В юкстагломерулярных клетках почек вырабатывается ренин , являющийся ключевым регулятором ренин-ангиотензиновой системы , которая отвечает за регулирование кровяного давления .
Выработка эритропоэтина почками отвечает за дифференцирование клеток-предшественников эритроидного ряда в костном мозге в эритроциты и индуцируется гипоксией . Таким образом при недостатке кислорода повышается количество красных кровяных телец в крови, которые отвечают за перенос кислорода .
Почки участвуют в метаболизме витамина D . В печени витамин D преобразуется в (25OHD), почки же преобразуют кальцифедиол в кальцитриол (1,25(OH) 2 D), который является активной формой витамина и по своей сути является гормоном . Витамин D участвует в формировании костей и хрящевой ткани, а также выполняет ряд других функций, например, участвует в работе иммунной системы .
Регуляция кровяного давления
Некоторые внутренние органы млекопитающих, включая почки и лёгкие, рассчитаны на функционирование в пределах нормального уровня кровяного давления и нормального уровня объёма крови, а само кровяное давление также зависит от изменений уровня объёма крови. Поэтому поддержание постоянства объёма крови для млекопитающих является очень важной функцией организма . На постоянство объёма крови оказывают влияние скорость клубочковой фильтрации, работа отдельный частей нефрона , симпатическая нервная система и ренин-ангиотензин-альдостероновая система .
В стенках афферентных артериол, у входа в почечные клубочки, расположены юкстагломерулярные клетки . Эти клетки являются чувствительными к изменению минутного объёма кровообращения, к составу и объёму внеклеточной жидкости, вырабатывая в ответ на изменения ренин . Попадая в кровоток, ренин преобразует ангиотензиноген в ангиотензин I . Ангиотензин I далее расщепляется ангиотензинпревращающим ферментом до ангиотензина II, который является сильным сосудосуживающим средством, повышающим кровяное давление . Помимо ангиотензина II у млекопитающих могут образовываться и другие биологически активные вещества. Ангиотензин II может расщепляться до ангиотензина III, ангеотензина IV и ангиотензина (1—7) .
Кислотно-щелочной баланс
Поддержка кислотно-щелочного баланса является жизненно важной функцией, поскольку изменения в уровне pH влияют практически на все биологические процессы организма . У типичного млекопитающего нормальный уровень pH в среднем равен 7,4, повышенный уровень называется алкалозом , а пониженный — ацидозом . Как и в случае других позвоночных у млекопитающих кислотно-щелочной баланс поддерживается главным образом (HCO 3 - /CO 2 ), которая позволяет поддерживать постоянный уровень pH крови и внеклеточной жидкости . Данная буферная система описывается следующим уравнением :
HCO 3 - + H + ↔ H 2 CO 3 ↔ CO 2 + H 2 O
Регулирование кислотно-щелочного баланса посредством бикарбонатной буферной системы обеспечивается работой лёгких и почек . Лёгкие регулируют уровень CO 2 ( углекислого газа ), а почки — уровень HCO 3 - и H + (ионов водорода и гидрокарбоната ) . При этом почки играют ключевую роль в поддержании постоянного уровня кислотно-щелочного баланса у млекопитающих . В почечных клубочках HCO 3 - полностью фильтруется в первичную мочу . Для поддержания постоянного уровня pH почки реабсорбируют обратно в кровоток почти весь HCO 3 - и секретируют в мочу H + , таким образом, происходит окисление мочи .
Реабсорбция HCO 3 - происходит в проксимальном канальце, в восходящей части петли Генле и в меньшей степени в дистальном извитом канальце нефрона . Секреция H + выполняется в основном посредством Na + /H + -обменников в канальцах нефрона . Собирательные трубочки участвуют в энергозависимой секреции H + . При попадании ионов H + в мочу они могут соединяться с отфильтрованным HCO 3 - с образованием угольной кислоты H 2 CO 3 , которая распадается на CO 2 и H 2 O (воду) под действием люминальной карбоангидразы. Образовавшийся CO 2 диффундирует в клетки канальцев, где он при участии цитозольной карбоангидразы соединяется с H 2 O и снова образует HCO 3 - , который затем возвращается в кровоток, а образовавшийся ион H + секретируется в мочу. Часть ионов H + секретируется с затратами энергии посредством АТФ -зависимого механизма .
Выводимая моча является слабокислой. Выделение H + вместе с мочой также происходит посредством буферных систем, в частности, NH 4 + ( аммония ) . Лишь небольшое количество NH 4 + фильтруется через клубочки , основная часть выводимого аммония является результатом окисления ионами H + образующегося в клетках проксимального извитого канальца NH 3 ( аммиака ), который секретируется в просвет канальца как NH 3 или как NH 4 + . Образование аммиака также сопровождается образованием нового HCO 3 - , который пополняет буферную систему крови . В толстом восходящем канальце петли Генли, наоборот, происходит всасывание NH 4 + , который затем перемещается в интерстиций . Окончательный этап окисления мочи происходит в собирательных трубочках, в которых с задействованием АТФ секретируются ионы H + , а из интерстиция транспортируется и секретируется NH 3 , который окисляется H + с образованием NH 4 + . За счёт регулирования реабсорбции HCO 3 - и секреции H + почки помогают поддерживать гомеостаз pH крови .
Гомеостаз глюкозы
Наряду с печенью , почки участвуют в поддержании гомеостаза глюкозы в организме . В почках происходят процессы фильтрации, реабсорбции и потребления глюкозы, а также производство глюкозы посредством глюконеогенеза . Потребление глюкозы ( гликолиз ) происходит преимущественно в мозговом веществе, глюконеогенез же происходит в корковом веществе . Гормонально процесс глюконеогенеза в почках регулируется с помощью инсулина и катехоламинов .
Эволюция
Возникновение у млекопитающих
Считается, что первые млекопитающие появились в Пермском периоде , который характеризуется холодными ночами в засушливых пустынях и выраженной сезонностью с длинными холодными зимами. Вероятно, холод и засушливость в те времена были значимыми факторами эволюционного давления. Развитие теплокровности у предшественников млекопитающих могло привести к увеличению интенсивности циркуляции крови, и, соответственно, к повышению артериального давления, которое, в свою очередь, увеличивало скорость клубочковой фильтрации почек. Однако увеличение скорости клубочковой фильтрации повлекло бы за собой и увеличение скорости вывода воды из организма . За концентрацию мочи и реабсорбцию воды отвечает тонкий сегмент канальца, являющийся частью петли Генле и присутствующий у всех млекопитающих . Можно предположить, что развитие механизма реабсорбции воды могло быть частью перехода к теплокровности, нежели прямой адаптацией к засушливости .
Адаптации к засушливости
Способность создавать более концентрированную мочу обратно зависит от массы тела животного, то есть чем меньше масса млекопитающего, тем более концентрированную мочу по отношению к животным с большей массой его почки могли бы производить в ходе адаптации к засушливому климату . Некоторые животные, обитающие в пустыне, эволюционно развили способность к куда большей концентрации мочи, чем у других животных . Наиболее концентрированную мочу среди изученных видов производит австралийская тушканчиковая мышь , в почках которой более длинные по сравнению с почками других млекопитающих петли Генли и удлинённый почечный сосочек . Более длинные петли у австралийских тушканчиковых мышей позволяют создавать очень концентрированную мочу и выживать в условиях недостатка воды . Исследования также показали, что эволюция концентрации мочи у разных пустынных млекопитающих была конвергентной .
Адаптации к увеличению массы
Самым простым типом почек у млекопитающих является однососочковая однодолевая почка, состоящая из коркового вещества, мозгового вещества и почечной лоханки . Однако однодолевая почка имеет некоторый верхний предел по количеству нефронов , при котором она работает оптимальным образом . Предположительно, именно этот тип почек мог у млекопитающих быть изначальным, от которого в ходе эволюции произошли многодолевые почки .
Многодолевые почки, вероятно, появились в качестве адаптации к увеличению массы тела млекопитающих и соответствующей необходимости в увеличении количества нефронов в почках . Дальнейшим адаптационным механизмом является увеличение размера почечных клубочков у крупных млекопитающих (и, соответственно, увеличения длины канальцев), как например, в случае слонов , у которых диаметр клубочка может быть в 2 раза больше, чем у косаток .
Появлению множественных почек, вероятно, способствовали как дальнейшее увеличение массы тела, так и особенности среды обитания . Множественные почки, вероятно, позволяют увеличивать количество нефронов добавлением отдельных почечек без необходимости увеличения длины канальцев при увеличении размера органа .
Множественные почки
Множественные почки характерны в основном для морских млекопитающих . Считается, что они являются адаптацией как к большой массе, позволяя наращивать количество нефронов с увеличением количества почечек, так и к диете с большим количеством солёной воды , а также к необходимости в течение длительного времени погружаться под воду. Потребление излишней соли приводит к внутриклеточной дегидратации , в результате возникает необходимость в скорейшем выводе излишней соли из организма, чему в случае множественных почек способствует увеличение общей площади между корковым и мозговым веществом . Необходимость погружаться на длительное время под воду требует сокращения потребления кислорода организмом , в то время как почки являются энергозатратным органом , поэтому во время погружений снижается скорость клубочковой фильтации . Между же погружениями скорость клубочковой фильтрации у множественных почек является достаточно высокой .
Развитие почек
Стадии развития почек
У млекопитающих конечной почкой является метанефрическая почка, однако развитие почек происходит в 3 этапа с развитием 3 различных видов почек на этапе эмбрионального развития : пронефроса , мезонефроса и метанефроса . Все 3 вида развиваются из промежуточной мезодермы последовательно в кранио-каудальном направлении (в направлении со стороны головы к хвостовой части тела) . Сначала формируется пронефрос (предпочка), у млекопитающих он считается рудиментарным , то есть не функционирует . Затем каудальнее пронефроса развивается мезонефрос (первичная почка), являющийся функционирующей почкой эмбриона . Впоследствии у самок мезонефрос деградирует, а у самцов участвует в развитии половой системы . Третьей стадией является формирование в каудальной части зародыша метанефроса — постоянной почки .
Развитие метанефроса
Метанефрос развивается из зачатка мочеточника , выступающего на каудальной части первичного почечного протока , и метанефрогенной бластемы, являющейся частью промежуточной мезодермы , окружающей зачаток мочеточника . Развитие метанефроса начинается с индуцирования мочеточниковым зачатком метанефрогенной бластемы , при этом по мере развития зачаток мочеточника и метанефрогенная бластема взаимно индуцируют друг друга . Прорастая в мезодерму, зачаток мочеточника постепенно ветвится и преобразуется в древовидную структуру, которая в конечном итоге станет мочеточником , почечной лоханкой, большими и малыми чашечками, почечными сосочками и собирательными трубочками . В то же время на концах прорастающих собирательных трубочек мезодерма дифференцируется в эпителиальные клетки, формирующие канальцы нефрона (происходят процессы эпителизации и тубулогенеза) . По мере развития нефронов развивается и кровоснабжение почки, при этом крупные сосуды берут начало ветвлением из спинной аорты .
У одних млекопитающих органогенез почек заканчивается до рождения, в то время как у других может продолжаться некоторое время и в послеродовой период (например, у грызунов он заканчивается примерно через неделю после рождения) . Когда формирование новых нефронов ( нефрогенез ) заканчивается, количество нефронов в почке становится окончательным .
Постнатальное созревание
После рождения и в постнатальный период почки функционально ещё незрелые, функциональное развитие почек у всех млекопитающих отстаёт от анатомического . В постнатальный период масса канальцев недостаточно большая, поэтому возможности по реабсорбции жидкостей снижены по сравнению с почками взрослых млекопитающих . В этот период происходит гипертрофия и гиперплазия канальцев, почки увеличиваются в размерах. Период, в течение которого формируются полностью функциональные почки, сильно варьируется между видами млекопитающих. У крыс почки достаточно быстро становятся полностью функциональными, в то время как у обезьян на это уходит 5 месяцев .
Повреждения и заболевания
Заболевания или нарушения, связанные с почками, могут быть врождёнными , наследственными , неинфекционными или инфекционными . Между отдельными видами млекопитающих заболевания могут различаться. Могут встречаться заболевания, специфичные для одних видов и не встречающиеся у других, либо встречающиеся у одних видов реже, чем у других . Например, характерна для мышей , крыс и голых землекопов , но в то же время у человека идентичного заболевания нет .
Врождённые и наследственные аномалии
Врождённые и наследственные аномалии почек млекопитающих встречаются достаточно редко, но могут оказывать значимое влияние на работу почек , в некоторых случаях могут приводить к смерти в раннем неонатальном периоде . Среди аномалий развития почек встречаются гипоплазия и дисплазия почек (дисплазия может быть односторонней или двусторонней), агенезия (отсутствие) одной или обеих почек, поликистоз почек , простые почечные кисты , периренальные псевдокисты , удвоение или утроение почечных артерий , неправильное расположение почек ( ) , и нефробластома .
Неинфекционные заболевания
В число неинфекционных заболеваний непосредственно почек входят острое повреждение почки, хроническая болезнь почек , гломерулярные болезни и заболевания, связанные с канальцами ( почечный тубулярный ацидоз , синдром Фанкони и почечная глюкозурия ) . Новообразования в почках маленьких млекопитающих встречаются редко, но обычно не являются доброкачественными . Новообразования и абсцессы являются редкими среди жвачных . Обструктивные уропатии (препятствие оттоку мочи из одной или обеих почек) могут приводить к гидронефрозу с расширением почечной лоханки . В почках также могут образовываться почечные камни ( почечнокаменная болезнь ) .
Причиной острого повреждения почек в большинстве случаев являются ишемические или токсические поражения . Почки млекопитающих предрасположены к ишемическому повреждению, поскольку у млекопитающих отсутствует почечно-портальная система кровообращения, в результате чего вазоконстрикция сосудов в почечных клубочках может приводить к снижению уровня снабжения кровью почки в целом. К токсическому же повреждению почки предрасположены, поскольку в канальцах вместе с большинством отфильтрованных веществ может происходить реабсорбция и токсинов . После острого повреждения почки способны восстановить свою работоспособность, однако оно способно перерасти в хроническое заболевание почек. При хроническом заболевании почек происходит потеря функциональности тканей почки, а само заболевание обычно является прогрессирующим .
Инфекционные заболевания
Инфекционные заболевания почек у маленьких млекопитающих обычно вызываются аэробными бактериями, среди которых кишечная палочка , стафилококки , энтерококки и стрептококки . Грибковые и паразитарные инфекции почек у маленьких млекопитающих встречаются редко . Пиелонефрит обычно развивается из-за бактерий, которые попадают в почки по восходящему пути из нижних частей мочевыделительной системы, реже через кровь . У жвачных пиелонефрит чаще всего вызывается бактерией и кишечной палочкой . Питающиеся рыбой млекопитающие (например, норки или собаки), могут заражаться гигантской нематодой . Свиньи могут заражаться червём , который встречается во всём мире, но больше в тропиках и субтропиках . Среди морских млекопитающих инфекции почек считаются редкими .
Старение
После созревания почек в них постепенно начинают происходить процессы старения, характерируемые изменениями в части анатомии , физиологии , функции и регенеративных возможностей. В течение жизни млекопитающих почечные клубочки подвергаются гломерулосклерозу, утолщается базальная мембрана , канальцы подвергаются атрофическим изменениям, а почечный интерстиций — фиброзу . Постепенно, но довольно медленно, снижается количество функционирующих нефронов. По части функции снижается скорость клубочковой фильтрации и способность концентрации мочи. Сами по себе возрастные изменения могут быть незаметными и могут не приводить в конечном итоге к почечной недостаточности или развитию заболевания, но являются фактором риска в случае возникновения заболевания почек или мочевыводящей системы .
Регенерация и способности к восстановлению
В отличие от более примитивных позвоночных , таких как рыбы , у млекопитающих нефрогенез заканчивается до или через некоторое время после рождения , когда заканчивается конденсированная мезенхима метанефротической бластемы, из которой новые нефроны образуются . В результате у взрослых особей новые нефроны образовываться не могут . Как следствие, после перенесённых повреждений почки взрослых млекопитающих не могут регенерировать путём образования новых нефронов . Однако существуют другие компенсаторные и регенеративные механизмы восстановления функции почек .
Компенсаторные возможности
В случае односторонней нефрэктомии нагрузка на оставшуюся почку увеличивается, увеличивается скорость фильтрации и реабсорбции , происходят изменения в самих нефронах. Почечный клубочек может увеличиться в диаметре в два или три раза. Подобные компенсаторные изменения схожи с изменениями в нефронах, которые происходят после рождения по мере роста почки . Резекция тканей почки также не вызывает процессов её регенерации , однако при повреждениях почек, в результате которых количество нефронов значительно сокращается, могут происходить компенсаторные изменения .
Регенерация нефрона
В рамках отдельно взятого нефрона регенеративные способности различаются между его частями . При острых токсических и ишемических повреждениях канальцы способны регенерировать для восстановления функциональности нефрона . В частности, способностью к регенерации обладает проксимальная часть нефрона , через которую всасывается до двух третей первичной мочи . Именно эта части нефрона у млекопитающих наиболее подвержена риску ишемических или токсических повреждений . Помимо этого, постоянное восстановление нефронов происходит в ходе нормальной физиологической деятельности из-за отслоения клеток эпителия канальцев . Почечное тельце имеет сложную структуру, и его возможности восстановления после повреждения ограничены . Мезангиальные (межсосудистые) и эндотелиальные клетки после повреждений способны пролиферировать и восстанавливать свою популяцию. Подоциты же в нормальных условиях не восстанавливаются .
Заживление тканей при повреждениях
В случае небольших повреждений канальцев нефрона потерянные клетки заменяются новыми, и эпителий регенерирует, восстанавливая свою структуру и функцию. При средних или тяжёлых повреждениях с большой потерей клеток шансы на регенерацию эпителия канальцев снижаются . В таких случаях не происходит регенеративное восстановление тканей , и в ответ на повреждения возникают воспалительный ответ, и развивается фиброз тканей , являющиеся следующей линией защиты организма , которая в ходе эволюции млекопитающих должна была снижать возможное кровотечение и противодействовать возможной инфекции . Подобная реакция характерна для острого повреждения почек , после которой почка способна восстановить свою работоспособность . Фиброз является результатом неудачного заживления тканей почки и ассоциируется с нарушением работы почек , однако существует мнение, что фиброз помогает поддерживать выживание неповреждённых и частично повреждённых нефронов . Хронические повреждения почек сопровождаются фиброзом, рубцеванием и потерей функциональности тканей , что характерно для хронической болезни почек .
Примечания
- ↑ , Abstract, p. 1.
- , 1.2.8 Excretion, p. 25.
- ↑ , 7.3. Mammals, p. 8.
- ↑ : [ англ. ] / Ed.: Jo Ann Eurell, Brian L. Frappier. — 6th Edition. — Blackwell Publishing , 2006, July . — P. 566. — ISBN 978-0-7817-4148-4 . — WD .
- ↑ , 3.6.3 The Kidney, p. 250.
- : [ англ. ] : [ 9 марта 2023] / Jane C. Fenelon , M. Caleb, G. Shaw [et al.] // Sexual Development . — 2021, 19 April . — Vol. 15, iss. 4. — P. 262—271. — ISSN , . — doi : . — WD .
- ↑ , Abstract, p. 679.
- , 3.4 Видовые, линейные и половые различия в строении и функции почек, с. 72-73.
- ↑ , Structure, p. 617.
- ↑ , Kidney structure, p. 1832.
- ↑ , Renal vasculature, p. 595.
- ↑ , Introduction, p. 2.
- ↑ , The Evolution of Renal Structures Was Driven by Body Size and Habitats in Mammals, p. 6.
- ↑ , Figure 1. Structure of the mammalian kidney.
- .
- , p. R1723.
- , с. R1722-R1723.
- , Summary, p. 1.
- , An Overview of Cell Players and Cellular Processes in Metanephric Kidney Development, p. 2.
- : [ англ. ] : [ 15 августа 2022] / Jing Yu, M. Todd Valerius, Mary Duah [et al.] // Development: the journal of the Society for International Development . — 2012, 1 May . — Vol. 139, iss. 10. — P. 1863—1873. — ISSN , , , , . — doi : . — PMID . — WD .
- ↑ Bobulescu IA . : [ англ. ] : [рукопись] / Bobulescu IA, Moe OW // Seminars in nephrology . — 2006, September . — Vol. 26, iss. 5. — P. 334—344. — ISSN , . — doi : . — PMID . — WD .
- ↑ Stanniocalcin 1 effects on the renal gluconeogenesis pathway in rat and fish : [ англ. ] / Vanessa Schein , Luiz C Kucharski, Pedro M. Guerreiro [et al.] // Molecular and Cellular Endocrinology . — 2015, 15 July . — Vol. 414. — P. 1—8. — ISSN , . — doi : . — PMID . — WD .
- : Adaptation, Diversity, Ecology : [ англ. ] / George A. Feldhamer , L. Drickamer , S. Vessey [et al.] . — 4th Edition. — Baltimore : JHU Press , 2015. — P. 198. — ISBN 978-1-4214-1588-8 . — WD .
- Maria L. S. Sequeira Lopez . : [ англ. ] : [рукопись] / Maria L. S. Sequeira Lopez, R. Ariel Gomez // Current Opinion in Nephrology and Hypertension . — 2010, 1 July . — Vol. 19, iss. 4. — P. 366—371. — ISSN , , . — doi : . — PMID . — WD .
- Sukanya Suresh . : [ англ. ] : [ 8 мая 2022] / Sukanya Suresh, Praveen Kumar Rajvanshi , Constance T. Noguchi // Frontiers in Physiology . — 2019, 1 January . — Vol. 10. — P. 1534. — ISSN . — doi : . — PMID . — WD .
- ↑ Bikle D. D. : [ англ. ] : [ 14 июля 2022] // Experimental Dermatology . — 2011, 1 January . — Vol. 20, iss. 1. — P. 7—13. — ISSN , . — doi : . — PMID . — WD .
- : Cells and Animals : [ англ. ] / Ed.: D. H. Evans. — CRC Press , 2008, 18 November . — P. 506. — ISBN 978-0-8493-8030-3 . — WD .
- Óscar Cortadellas Rodríguez . : [ англ. ] / Óscar Cortadellas Rodríguez, María Luisa Suárez Rey; illus.: Jacob Gragera Artal; transl.: Owen Howard. — Grupo Asís Biomedia S.L., 2018, February . — P. 5. — ISBN 978-84-17225-34-6 . — WD .
- ↑ : [ англ. ] : [ 27 сентября 2023] // Merck Veterinary Manual. — Дата обращения: 21 сентября 2023.
- ↑ : [ англ. ] : [ 27 сентября 2023] // Merck Veterinary Manual. — Дата обращения: 24 сентября 2023.
- ↑ : [ англ. ] / T Taghipur Bazargani, A Khodakaram-Tafti, I Ashrafi, A M Abbassi // Iranian Journal of Veterinary Research . — 2015, 1 January . — Vol. 16, iss. 1. — P. 114—116. — ISSN . — PMID . — WD .
- ↑ : [ англ. ]. Vol. 30. Kidney Disease / Ed.: G. W. Richter, Kim Solez . — Academic Press , 1988. — P. 231—232. — ISSN . — ISBN 0-12-364930-7 . — WD .
- ↑ : [ англ. ] : [энциклопедия] / Ed.: H. Mehlhorn . — 3rd Edition. — Springer , 2007, 28 November . — P. 1532. — ISBN 978-3-540-48996-2 , 978-3-540-48997-9 , 978-3-540-48994-8 . — doi : . — WD .
- ↑ : [ англ. ] : [ 4 июня 2023] // Merck Veterinary Manual. — Дата обращения: 24 сентября 2023.
- ↑ Katja Berger . : [ англ. ] / Katja Berger, Marcus J. Moeller // Seminars in nephrology . — 2014, 13 June . — Vol. 34, iss. 4. — P. 394—403. — ISSN , . — doi : . — PMID . — WD .
- ↑ , 7 Reassessing Renal Disease, Repair, and Regeneration Using Developmental Biology, p. 12.
- ↑ , Figure 1 Schematic illustration highlighting patchy regenerative/reparative processes after mammalian acute kidney injury, p. 28.
- ↑ , Introduction, p. 1435.
- ↑ , Postnatal regenerative response of the mammalian kidney, p. 1437-1439.
- ↑ Haller H. : [ англ. ] / H. Haller, S. Sorrentino // Regenerative Medicine : From Protocol to Patient / Ed.: G. Steinhoff . — Springer , 2011. — P. 811. — ISBN 978-90-481-9075-1 , 978-90-481-9074-4 . — WD .
- ↑ Chevalier Robert L. : [ англ. ] // KI reports . — 2017, 31 January . — Vol. 2, iss. 3. — P. 302—317. — ISSN . — doi : . — PMID . — WD .
- ↑ , Aging of the Kidney, p. 637.
- : [ англ. ] : [ 10 марта 2023] / Katerina Apelt, Roel Bijkerk , Franck Lebrin , Ton J. Rabelink // Cells . — 2021, 2 May . — Vol. 10, iss. 5. — ISSN . — doi : . — PMID . — WD .
- Lauralee Sherwood . : From Genes to Organisms : [ англ. ] / Lauralee Sherwood, Hillar Klandorf , Paul H. Yancey. — 2nd Edition. — Cengage Learning , 2012. — P. 569. — ISBN 978-0-8400-6865-1 . — WD .
- : [ англ. ] / P. C. Withers, C. E. Cooper, S. K. Maloney [et al.] . — Oxford University Press , 2016. — P. 250. — ISBN 978-0-19-964271-7 , 978-0-19-964272-4 . — WD .
- Christopher Thigpen . : [ англ. ] : [ 22 мая 2022] / Christopher Thigpen, Logan Best, Troy Camarata // Zoomorphology . — 2019, 28 September . — Vol. 139. — P. 111–121. — ISSN , . — doi : . — WD .
- ↑ Donald W. Linzey . : [ англ. ]. — 2nd Edition. — 2012, 13 February . — P. 319. — ISBN 978-1-4214-0040-2 . — WD .
- C. De Martino . : [ англ. ] / C. De Martino, D. J. Allen, L. Accinni // Basic, Clinical, and Surgical Nephrology / Ed.: L. J. A. Didio, P. M. Motta. — Boston : Martinus Nijhoff Publishers , 1985. — P. 53–82. — ISBN 978-1-4612-9616-4 , 978-1-4613-2575-8 . — WD .
- ↑ , Figure 1 Structure of the mammalian kidney.
- Moffat D. B. : [ англ. ]. — Cambridge University Press , 1975. — P. 16—18. — (Biological Structure and Function Books; 5). — ISBN 0-521-20599-9 . — WD .
- ↑ The Editors of Encyclopaedia . : [ англ. ] : [ 2 мая 2022] // Encyclopædia Britannica : online encyclopedia. — Дата обновления: 20 июня 2018. — Дата обращения: 2 мая 2022.
- James Arthur Ramsay . : [ англ. ] / James Arthur Ramsay, Fenton Crosland Kelley // Encyclopædia Britannica : online encyclopedia. — 2020, 2 April . — Дата обращения: 4 июня 2022.
- , Figure 20.9 Schematic of nephrons and collecting duct, p. 602.
- : [ англ. ] // SEER Training. — U. S. National Cancer Institute. — Дата обращения: 29 июля 2022.
- Dyce Keith M. : [ англ. ] / Keith M. Dyce, Wolfgang O. Sack, C. J. G. Wensing. — Fourth Edition. — Saunders , 2010. — P. 177. — ISBN 978-1-4160-6607-1 . — WD .
- , Figure 1. Structure of the mammalian kidney, p. 2.
- Zhuo Jia L. : [ англ. ] : [рукопись] / Jia L. Zhuo, Xiao C. Li // Comprehensive Physiology . — 2013, July . — Vol. 3, iss. 3. — P. 1079—123. — ISSN . — doi : . — PMID . — WD .
- , Anatomy, p. 379.
- ↑ , Kidney Structure, с. 1464.
- , Figure 1.1 A long-looped and short-looped nephron together with the collecting system, p. 4-5.
- , Nephrons and Collecting Duct System, p. 600.
- ↑ , Kidney Types and Renal Pelvis, p. 595.
- ↑ , Cortex, p. 6.
- ↑ : [ англ. ] / Ed.: Xin J. Zhou [et al.] . — Cambridge University Press , 2017, 2 March . — P. 19. — WD .
- , Anatomy, p. 378.
- , Overview of kidney structure and embryonic development.
- ↑ : Morphology, Biochemistry, Physiology : [ англ. ]. Vol. 1 / Ed.: C. Rouiller, A. F. Muller. — Academic Press , 1969. — P. 357. — ISBN 978-1-4832-7174-3 . — WD .
- ↑ , Outer Medulla, p. 8.
- Kriz W. : [ англ. ] // Paediatric nephrology . — Springer Verlag , 1984. — P. 3–10. — ISBN 978-3-540-13598-2 , 978-3-642-69863-7 . — doi : . — WD .
- ↑ , p. R1722.
- , 2.2.6 Mammals, p. 20.
- , 3.4 Видовые, линейные и половые различия в строении и функции почек, с. 72—73.
- , 3.4 Видовые, линейные и половые различия в строении и функции почек, с. 73.
- , 3. Results and discussion, p. 3.
- ↑ , 3.4 Видовые, линейные и половые различия в строении и функции почек, с. 72.
- : [ англ. ] : [ 27 декабря 2022] / Kendall S. Frazier, John Curtis Seely, Gordon C. Hard [et al.] // Toxicologic Pathology . — 2012, 1 June . — Vol. 40, iss. 4 Suppl. — P. 14S—86S. — ISSN , . — doi : . — PMID . — WD .
- Richard Nickel . = Lehrbuch der Anatomie der Haustiere : [ англ. ] : Transl. from German / Richard Nickel, August Schummer, Eugen Seiferle. — Second revised edition. — Berlin , 1979. — P. 286. — ISBN 978-1-4757-6816-9 . — doi : . — WD .
- , 2.2.6 Mammals, p. 19—20.
- Sur R. L., Meegan J. M., Smith C. R., Schmitt T., L'Esperance J., Hendrikson D., Woo J. R. (англ.) // — 2018. — Vol. 4, Iss. 1. — P. 62—65. — ISSN — —
- , 7.3.1. Mammalian kidneys: overall morphology, p. 8.
- ↑ Little Melissa H. : [ англ. ] : [ 12 марта 2023] // Developmental Biology . — 2020, 14 December . — ISSN , . — doi : . — PMID . — WD .
- , Detailed Features of Human and Mammalian Renal Lymphatic Anatomy : Renal Interstitium, p. 6.
- , Interstitium : Definition, p. 602.
- ↑ Michael Zeisberg . : [ англ. ] / Michael Zeisberg, Raghu Kalluri // Clinical Journal of the American Society of Nephrology . — 2015, 26 March . — Vol. 10, iss. 10. — P. 1831—1840. — ISSN , . — doi : . — PMID . — WD .
- ↑ , Detailed Features of Human and Mammalian Renal Lymphatic Anatomy : Morphology of Renal Lymph Vessels, p. 6.
- , Interstitial Fibroblasts, p. 602.
- , Interstitium, p. 622.
- : [ англ. ] : [ 13 марта 2023] / A. Schumacher, M. B. Rookmaaker, J. A. Joles [et al.] // Npj regenerative medicine . — 2021, 11 August . — Vol. 6. — Article 45. — ISSN . — doi : . — WD .
- C Schell . Glomerular development--shaping the multi-cellular filtration unit : [ англ. ] / C Schell, N Wanner, T B Huber // Seminars in Cell & Developmental Biology . — 2014, 18 August . — Vol. 36. — P. 39—49. — ISSN , . — doi : . — PMID . — WD .
- ↑ : [ англ. ] : [ 13 марта 2023] / Lihe Chen, Jevin Z Clark, J. Nelson [et al.] // Journal of the American Society of Nephrology . — 2019, 28 June . — Vol. 30, iss. 8. — P. 1358—1364. — ISSN , . — doi : . — PMID . — WD .
- Jae Wook Lee . : [ англ. ] : [ 13 марта 2023] / Jae Wook Lee, Chung-Lin Chou, Mark A Knepper // Journal of the American Society of Nephrology . — 2015, 27 March . — Vol. 26, iss. 11. — P. 2669—2677. — ISSN , . — doi : . — PMID . — WD .
- , Introduction, p. 1.
- ↑ , Vascular supply, p. 379.
- , Renal vasculature, p. 596.
- Holz Peter H. : [ англ. ] // Bulletin of the Association of Reptilian and Amphibian Veterinarians . — 1999, 1 January . — Vol. 9, iss. 1. — P. 4–14. — ISSN . — doi : . — WD .
- Kotpal R. L. : Vertebrates : [ англ. ]. Vol. 2. — Rastogi Publications, 2010. — P. 782. — ISBN 978-81-7133-891-7 . — WD .
- , 6. Renal portal system, p. 6.
- Schmidt-Nielsen K. : [ англ. ]. — Cambridge University Press , 1997, 9 June . — P. 371. — ISBN 0-521-57098-0 . — WD .
- : [ англ. ] / Ed.: John E. Reynolds. — Smithsonian Institution , 2013. — P. 299. — ISBN 978-1-58834-420-5 . — WD .
- David A D Munro . : [ англ. ] / David A D Munro, P. Hohenstein , J. A. Davies // Scientific Reports . — 2017, 12 June . — Vol. 7, iss. 1. — P. 3273. — ISSN . — doi : . — PMID . — WD .
- , Introduction, p. 1.
- : [ англ. ] : [ 13 марта 2023] / Elaine L. Shelton, Hai-Chun Yang, Jianyong Zhong [et al.] // American journal of physiology : Renal Physiology. — 2020, 26 October . — ISSN , , . — doi : . — PMID . — WD .
- , Renal Lymphatic Physiology under Normal Conditions : Formation of Renal Lymph, p. 7.
- ↑ Harald Seeger . : [ англ. ] : [ 13 марта 2023] / Harald Seeger, Marco Bonani, Stephan Segerer // Nephrology Dialysis Transplantation . — 2012, 23 May . — Vol. 27, iss. 7. — P. 2634—2641. — ISSN , . — doi : . — PMID . — WD .
- , Anatomy of Renal Lymphatics : Renal Vascular Anatomy, p. 2.
- ↑ , Comparative Renal Lymphatic Anatomy : Mammalian Renal Lymphatic Anatomy, p. 3.
- , Renal Lymphatic Physiology under Normal Conditions : Interstitial Fluid and Protein Drainage in the Medulla, p. 9.
- ↑ , Detailed Features of Human and Mammalian Renal Lymphatic Anatomy : Medullary Lymphatics, p. 5.
- , Table 1. Comparison of renal lymphatic anatomy between species, p. 5.
- ↑ , 2.2 Intrarenal Distribution of Efferent Renal Sympathetic Nerves, p. 6.
- // Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов . — М. : Большая российская энциклопедия, 2004—2017.
- Yasuna Nakamura . : [ англ. ] / Yasuna Nakamura, Tsuyoshi Inoue // JMA journal . — 2020, 8 July . — Vol. 3, iss. 3. — P. 164—174. — ISSN , . — doi : . — PMID . — WD .
- , Abstract.
- , Nerves, p. 610.
- Kopp Ulla C. : [ англ. ] // / Ulla C. Kopp. — San Rafael : Morgan & Claypool Life Sciences, 2011, 20 August . — WD .
- Kopp Ulla C. : [ англ. ] // / Ulla C. Kopp. — San Rafael : Morgan & Claypool Life Sciences, 2011, 20 August . — WD .
- , Introduction, p. 1.
- , 8.1. Activation of Afferent Renal Sensory Nerves by Physiological Stimuli.
- ↑ Knepper MA . : [ англ. ] / Knepper MA, Roch-Ramel F // Kidney International . — 1987, 1 February . — Vol. 31, iss. 2. — P. 629—633. — ISSN , . — doi : . — PMID . — WD .
- James Arthur Ramsay . : [ англ. ] / James Arthur Ramsay, Fenton Crosland Kelley // Encyclopædia Britannica : online encyclopedia. — Дата обновления: 2 апреля 2020. — Дата обращения: 4 июня 2022.
- Fenton R. : [ англ. ] : [ 9 марта 2023] / R. Fenton, Mark A Knepper // Journal of the American Society of Nephrology . — 2007, March . — Vol. 18, iss. 3. — P. 679—88. — ISSN , . — doi : . — PMID . — WD .
- , 8.4 Terrestrial vertebrates, p. 121.
- ↑ , Introduction.
- Bernard C Rossier . : [ англ. ] : [ 22 сентября 2022] // The Nephron Journals . — 2016, 23 February . — Vol. 134, iss. 1. — P. 5—9. — ISSN , . — doi : . — PMID . — WD .
- Mark A Knepper . : [ англ. ] / Mark A Knepper, Tae-Hwan Kwon, Soren Nielsen // The New England Journal of Medicine . — 2015, 1 April . — Vol. 372, iss. 14. — P. 1349—1358. — ISSN , . — doi : . — PMID . — WD .
- ↑ , Adapting to living on dry land: the water-retaining kidney was invented twice, p. 718.
- Lote Christopher J. : [ англ. ]. — Springer New York , 2012, 22 June . — P. 70–85. — ISBN 978-1-4614-3785-7 , 978-1-4614-3784-0 . — doi : . — WD .
- : [ англ. ] : [ 14 марта 2022] / Liu W, Morimoto T, Kondo Y [et al.] // Kidney International . — 2001, 1 August . — Vol. 60, iss. 2. — P. 680—693. — ISSN , . — doi : . — PMID . — WD .
- , 1. Introduction, p. 1—2.
- Maria L. S. Sequeira Lopez . : [ англ. ] : [рукопись] / Maria L. S. Sequeira Lopez, R. Ariel Gomez // Current Opinion in Nephrology and Hypertension . — 2010, 1 July . — Vol. 19, iss. 4. — P. 366—371. — ISSN , , . — doi : . — PMID . — WD .
- Sukanya Suresh . : [ англ. ] : [ 8 мая 2022] / Sukanya Suresh, Praveen Kumar Rajvanshi , Constance T. Noguchi // Frontiers in Physiology . — 2019, 1 January . — Vol. 10. — P. 1534. — ISSN . — doi : . — PMID . — WD .
- , 11.5 The mammalian kidney, p. 164.
- : [ англ. ] / David Fournier, Friedrich C Luft , Michael Bader [et al.] // Journal of Molecular Medicine . — Vol. 90, iss. 5. — P. 495—508. — ISSN , . — doi : . — PMID . — WD .
- ↑ Sequeira-Lopez Maria Luisa S. : [ англ. ] : [ 26 сентября 2022] / Maria Luisa S. Sequeira-Lopez, R. Ariel Gomez // Circulation Research : scientific journal. — 2021, 2 April . — Vol. 128, iss. 7. — P. 887–907. — ISSN , . — doi : . — PMID .
- : A Tribute to Professor W.T.W.Potts : [ англ. ] / Ed.: N. Hazon [et al.] . — Springer-Verlag , 2012, 6 December . — P. 153. — ISBN 978-3-642-64396-5 , 978-3-642-60415-7 . — doi : . — WD .
- , Introduction, p. 1623.
- Lauralee Sherwood . : From Genes to Organisms : [ англ. ] / Lauralee Sherwood, Hillar Klandorf , Paul H. Yancey. — 2nd Edition. — Cengage Learning , 2012. — P. 637. — ISBN 978-0-8400-6865-1 . — WD .
- ↑ : [ англ. ] / Ed.: C. A. Wagner , Stefan Broer . — Springer Science+Business Media , 2004, 31 March . — P. 66. — ISBN 978-1-4613-4761-3 , 978-1-4419-9023-5 . — doi : . — WD .
- ↑ James L. Lewis III . : [ англ. ] : [ 19 марта 2023] // MSD Manuals — Medical Professional Version. — 2021, July . — Дата обращения: 17 марта 2023.
- ↑ , Bicarbonate Absorption, p. 84.
- ↑ , Renal ammonia handling : Fig. 3, p. 1629.
- ↑ , Renal ammonia handling, p. 1627.
- ↑ , Renal ammonia handling, p. 1627-1628.
- , Renal ammonia handling : Fig.2, p. 1628.
- Klaus Urich . : [ англ. ] / Illus.: Charlotte Urich; transl.: P. J. King. — Springer Berlin Heidelberg , 2013, 17 April . — P. 534. — ISBN 978-3-642-08181-1 , 978-3-662-06303-3 . — doi : . — WD .
- ↑ Leszek Szablewski . : [ англ. ]. — Bentham Science Publishers , 2011, 14 April . — P. 77—78. — ISBN 978-1-60805-189-2 . — WD .
- ↑ , p. 352.
- , p. 351.
- ↑ : [ англ. ] / Joana L. Rocha, José C. Brito, Rasmus Nielsen, Raquel Godinho // Mammal Review . — 2021, 1 March . — Vol. 51, iss. 4. — P. 482—491. — ISSN , . — doi : . — WD .
- Dantzler W. : [ англ. ] // BioScience . — 1982, February . — Vol. 32, iss. 2. — P. 108—113. — ISSN , . — doi : . — OCLC . — WD .
- Abraham Allan Degen . : [ англ. ] / Ed.: J. Thompson . — Springer , 1997. — P. 118. — ISBN 978-3-642-64366-8 , 978-3-642-60351-8 . — doi : . — WD .
- , Introduction, p. 1.
- ↑ , Apnea/simulated diving, p. 1838.
- Ying Chen . : [ англ. ] : [рукопись] / Ying Chen, Brendan C Fry, Anita T Layton // Bulletin of Mathematical Biology . — 2016, 1 July . — Vol. 78, iss. 6. — P. 1318—1336. — ISSN , . — doi : . — PMID . — WD .
- ↑ , Overview, pp. 859.
- ↑ Davies J. A. Kidney Development : [ англ. ] // ELS . — 2013, 19 September . — ISBN 978-0-470-01590-2 . — doi : . — WD .
- , 1. Overview of kidney structure and embryonic development.
- : [ англ. ] / Cristina Cebrián, Karolina Borodo, Nikki Charles, Doris A. Herzlinger // Developmental Dynamics . — 2004, 1 November . — Vol. 231, iss. 3. — P. 601—608. — ISSN , . — doi : . — PMID . — WD .
- Chambers J. : [ англ. ] / J. Chambers, Rebecca A Wingert // Tissue Barriers . — 2020, 22 October . — P. 1832844. — ISSN , . — doi : . — PMID . — WD .
- , Overview, pp. 859-860.
- ↑ Kispert A. : [ англ. ] / A. Kispert, S. Vainio , A. P. McMahon // Development: the journal of the Society for International Development . — 1998, 1 November . — Vol. 125, iss. 21. — P. 4225—4234. — ISSN , , , , . — doi : . — PMID . — WD .
- , Development of the Metanephros, p. 861-862.
- , Development of the Metanephros, p. 860.
- ↑ , Development of the Metanephros, p. 861.
- ↑ , Termination of the Kidney Development, p. 882.
- Abrahamson Dale R. : [ англ. ] // Organogenesis . — 2009, 1 January . — Vol. 5, iss. 1. — P. 275—287. — ISSN , . — doi : . — PMID . — WD .
- ↑ John Curtis Seely . : [ англ. ] // Journal of Toxicologic Pathology . — 2017, 11 February . — Vol. 30, iss. 2. — P. 125—133. — ISSN , . — doi : . — PMID . — WD .
- Kendall S. Frazier . Species Differences in Renal Development and Associated Developmental Nephrotoxicity : [ англ. ] // Birth Defects Research . — 2017, 2 August . — Vol. 109, iss. 16. — P. 1243—1256. — ISSN . — doi : . — PMID . — WD .
- : [ англ. ] / Ed.: Karen A. Terio [et al.] . — Academic Press , 2018, 8 October . — P. 503. — ISBN 978-0-12-805306-5 . — WD .
- Hard Gordon C. A comparison of rat chronic progressive nephropathy with human renal disease-implications for human risk assessment : [ англ. ] / Gordon C. Hard, Kent J. Johnson, Samuel M. Cohen // Critical Reviews in Toxicology . — 2009, 1 January . — Vol. 39, iss. 4. — P. 332—346. — ISSN , . — doi : . — PMID . — WD .
- : [ англ. ] : [ 27 сентября 2023] // Merck Veterinary Manual. — Дата обращения: 23 сентября 2023.
- ↑ : [ англ. ] : [ 27 сентября 2023] // Merck Veterinary Manual. — Дата обращения: 23 сентября 2023.
- ↑ : [ англ. ] : [ 27 сентября 2023] // Merck Veterinary Manual. — Дата обращения: 24 сентября 2023.
- : [ англ. ] : [ 27 сентября 2023] // Merck Veterinary Manual. — Дата обращения: 24 сентября 2023.
- : [ англ. ] : [ 27 сентября 2023] // Merck Veterinary Manual. — Дата обращения: 24 сентября 2023.
- : [ англ. ] : [ 27 сентября 2023] // Merck Veterinary Manual. — Дата обращения: 24 сентября 2023.
- : [ англ. ] / Ed.: Bradford P. Smith. — Fifth Edition. — Elsevier , 2014, April . — P. 768. — ISBN 978-0-323-08839-8 . — WD .
- : [ англ. ] : [ 27 сентября 2023] // Merck Veterinary Manual. — Дата обращения: 24 сентября 2023.
- : [ англ. ] : [ 27 сентября 2023] // Merck Veterinary Manual. — Дата обращения: 24 сентября 2023.
- ↑ : [ англ. ] : [ 27 сентября 2023] // Merck Veterinary Manual. — Дата обращения: 24 сентября 2023.
- : [ англ. ]. Vol. 1 / Ed.: Edwin B. Howard. — CRC Press , 2017, 13 December . — P. 30. — WD .
- , Cessation of mammalian nephrogenesis, p. 1436.
- ↑ , Postnatal regenerative response of the mammalian kidney, p. 1437.
- Ariela Benigni . Kidney regeneration : [ англ. ] / Ariela Benigni, Marina Morigi , G. Remuzzi // . — 2010, 1 April . — Vol. 375, iss. 9722. — P. 1310—1317. — ISSN , . — doi : . — PMID . — WD .
- , Conclusions and perspectives, p. 1441.
- ↑ , What Is Kidney Regeneration?.
- ↑ : [ англ. ] // Kidney Transplantation in the Regenerative Medicine Era : Kidney Transplantation in the Regenerative Medicine Era / Ed.: David F. Williams [et al.] . — Academic Press , 2017, 8 June . — P. 997. — ISBN 978-0-12-801734-0 . — WD .
- , Postnatal regenerative response of the mammalian kidney, p. 1438.
- , Introduction.
- , Mechanisms of Kidney Regeneration.
- Qi Cao . Macrophages in kidney injury, inflammation, and fibrosis : [ англ. ] / Qi Cao, David C H Harris, Yiping Wang // Physiology . — 2015, 1 May . — Vol. 30, iss. 3. — P. 183—194. — ISSN , , , . — doi : . — PMID . — WD .
- : A Model for Organogenesis, Human Disease, and Evolution : [ англ. ] / Ed.: Kiyoshi Naruse [et al.] . — Tokyo ; NYC : Springer , 2011, 30 May . — P. 112. — ISBN 978-4-431-92690-0 , 978-4-431-92691-7 . — doi : . — WD .
- , Impact of Renal Fibrosis on Human Health, p. 3.
- , Renal Fibrosis: Aetiology and Pathophysiology, p. 2.
Литература
Книги
- = Principles and methods for the assessment of nephrotoxicity associated with exposure to chemicals : [ 11 сентября 2022] : Пер. с англ. — Женева : ВОЗ , 1994. — С. 72—73. — (Гигиенические критерии состояния окружающей среды; 119). — ISBN 5-225-01924-2 . — WD .
- Davidson Alan J. : [ англ. ] : [ 4 марта 2022] // . — 2009, 15 January . — doi : . — PMID . — WD .
- Dantzler W. : [ англ. ]. — Springer, 2016, 5 July . — 292 p. — ISBN 978-1-4939-3734-9 . — WD .
- Eladari D. : [ англ. ] / D. Eladari, U. Hasler, E. Féraille // : Physiology and pathophysiology : in 2 vols. / Ed.: R. J. Alpern [et al.] . — Fifth edition. — Amsterdam : Academic Press , 2012, 31 December . — Vol. 1—2. — P. 84—85. — ISBN 978-0-12-381463-0 , 978-0-12-381462-3 . — WD .
- Kriz W. : [ англ. ] / Kriz W., Kaissling B. // : Physiology and pathophysiology : in 2 vols. / Ed.: R. J. Alpern [et al.] . — Fifth edition. — Amsterdam : Academic Press , 2012, 31 December . — Vol. 1—2. — P. 595—691. — ISBN 978-0-12-381463-0 , 978-0-12-381462-3 . — WD .
- Sands J. M. : [ англ. ] / J. M. Sands, H. E. Layton // : Physiology and pathophysiology : in 2 vols. / Ed.: R. J. Alpern [et al.] . — Fifth edition. — Amsterdam : Academic Press , 2012, 31 December . — Vol. 1—2. — 3290 p. — ISBN 978-0-12-381463-0 , 978-0-12-381462-3 . — WD .
- Bush Kevin T. : [ англ. ] / Kevin T. Bush, Hiroyuki Sakurai, Sanjay K. Nigam // : Physiology and pathophysiology : in 2 vols. / Ed.: R. J. Alpern [et al.] . — Fifth edition. — Amsterdam : Academic Press , 2012, 31 December . — Vol. 1—2. — 3290 p. — ISBN 978-0-12-381463-0 , 978-0-12-381462-3 . — WD .
- : [ англ. ]. In 3 vols. Vol. 2 / Ed.: M. Grant Maxie. — Elsevier Health Sciences, 2015, 14 August . — P. 378—379. — ISBN 978-0-7020-5318-4 . — WD .
- : [ англ. ] / P. C. Withers, C. E. Cooper, S. K. Maloney [et al.] . — Oxford University Press , 2016. — 590 p. — ISBN 978-0-19-964271-7 , 978-0-19-964272-4 . — WD .
- Breshears M. A. : [ англ. ] : [ 30 июля 2022] / M. A. Breshears, A. W. Confer // Pathologic Basis of Veterinary Disease / Ed.: J. F. Zachary. — Sixth Edition. — St. Louis : Elsevier , 2017. — P. 617–681.e1. — ISBN 978-0-323-35775-3 . — WD .
- Kopp Ulla C. : [ англ. ]. — San Rafael : Morgan & Claypool Life Sciences, 2011, 20 August . — (Integrated Systems Physiology: from Molecule to Function to Disease). — PMID . — WD .
- Kopp Ulla C. : [ англ. ]. — Morgan & Claypool Publishers , 2018, 17 July . — 106 p. — (Colloquium Series on Integrated Systems Physiology From Molecule to Function; 1). — ISBN 978-1-61504-776-5 , 978-1-61504-775-8 , 978-1-61504-777-2 . — doi : . — WD .
- Bradley T. J. : [ англ. ]. — OUP Oxford , 2009. — P. 121, 164. — (Oxford Animal Biology Series). — ISBN 978-0-19-856996-1 . — WD .
- Sands Jeff M. : [ англ. ] / Jeff M. Sands, Jill W. Verlander // Toxicology of the Kidney / Ed.: Joan B. Tarloff, Lawrence H. Lash. — Third Edition. — CRC Press , 2004. — P. 3—54. — ISBN 0-203-67329-8 , 0-203-64699-1 , 0-415-24864-7 . — WD .
Статьи в журналах
- Schulte K. : [ англ. ] : [ 25 февраля 2022] / K. Schulte, Uta Kunter, Marcus J. Moeller // Nephrology Dialysis Transplantation . — 2014, 18 August . — Vol. 30, iss. 5. — P. 713—723. — ISSN , . — doi : . — PMID . — WD .
- Laura Keogh . : [ англ. ] : [ 3 марта 2022] / Laura Keogh, David Kilroy, Sourav Bhattacharjee // Annals of Anatomy . — 2020, 13 October . — Article 151610. — 15 p. — ISSN , . — doi : . — PMID . — WD .
- Casotti G. : [ англ. ] : [ 17 марта 2022] / G. Casotti, K. K. Lindberg, E. J. Braun // American journal of physiology. Regulatory, integrative and comparative physiology . — 2000, 1 November . — Vol. 279, iss. 5. — P. R1722—30. — ISSN , . — doi : . — PMID . — WD .
- Balzer Michael S. : [ англ. ] / Michael S. Balzer, Tibor Rohacs, Katalin Susztak // Annual Review of Physiology . — 2022, 10 February . — Vol. 84. — P. 507—531. — ISSN , . — doi : . — PMID . — WD .
- Abdalla M. A. : [ англ. ] : [ 26 июля 2022] // Heliyon . — 2020, 2 January . — Vol. 6, iss. 1. — Article e03139. — ISSN . — doi : . — PMID . — WD .
- Ortiz R. M. : [ англ. ] : [ 4 июня 2022] // The Journal of Experimental Biology . — 2001, 1 June . — Vol. 204, iss. Pt 11. — P. 1831—1844. — ISSN , . — doi : . — PMID . — WD .
- : [ англ. ] : [ 16 сентября 2022] / Peter Spencer Russell, Jiwon Hong, John Albert Windsor [et al.] // Frontiers in Physiology . — 2019. — Vol. 10. — P. 251. — ISSN . — doi : . — PMID . — WD .
- Sands J. : [ англ. ] : [рукопись] / J. Sands, Harold E. Layton // Seminars in nephrology . — 2009, 1 May . — Vol. 29, iss. 3. — P. 178—195. — ISSN , . — doi : . — PMID . — WD .
- Fenton R. : [ англ. ] : [ 9 марта 2023] / R. Fenton, Mark A Knepper // Journal of the American Society of Nephrology . — 2007, March . — Vol. 18, iss. 3. — P. 679—88. — ISSN , . — doi : . — PMID . — WD .
- Dominique Eladari . Renal acid-base regulation: new insights from animal models : [ англ. ] / Dominique Eladari, Yusuke Kumai // Pflügers Archiv : European Journal of Physiology. — 2014, 18 December . — Vol. 467, iss. 8. — P. 1623—1641. — ISSN , , . — doi : . — PMID . — WD .
- Alan J Davidson . Uncharted waters: nephrogenesis and renal regeneration in fish and mammals : [ англ. ] // Pediatric Nephrology . — 2011, 19 February . — Vol. 26, iss. 9. — P. 1435—1443. — ISSN , . — doi : . — PMID . — WD .
- Vize Peter D. : [ англ. ] / Peter D. Vize, Homer W. Smith // The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology . — 2004, 1 April . — Vol. 277, iss. 2. — P. 344—354. — ISSN , . — doi : . — PMID . — WD .
- Sanjeev Kumar . : [ англ. ] // Kidney International . — 2018, 1 January . — Vol. 93, iss. 1. — P. 27—40. — ISSN , . — doi : . — PMID . — WD .
- António Nogueira . : [ англ. ] / António Nogueira, Maria João Pires, Paula Alexandra Oliveira // In Vivo : International journal of experimental and clinical pathophysiology and drug research. — 2017, 1 January . — Vol. 31, iss. 1. — P. 1—22. — ISSN , . — doi : . — PMID . — WD .
- : [ англ. ] / Xu Zhou, Wenqi Rong, Boxiong Guo [et al.] // Genome Biology and Evolution . — 2023, 9 May . — Vol. 15, iss. 5. — ISSN . — doi : . — OCLC . — WD .
- Hai-Chun Yang . : [ англ. ] : [рукопись] / Hai-Chun Yang, Shao-Jun Liu, Agnes B. Fogo // The Nephron Journals . — 2014, 19 May . — Vol. 126, iss. 2. — P. 50. — ISSN , . — doi : . — PMID . — WD .
- Little Melissa H. : [ англ. ] : [ 21 января 2022] / Melissa H. Little, Andrew P. McMahon // Cold Spring Harbor perspectives in biology . — 2012, 1 May . — Vol. 4, iss. 5. — ISSN . — doi : . — PMID . — WD .
Дополнительная литература
- 2020-08-29
- 1