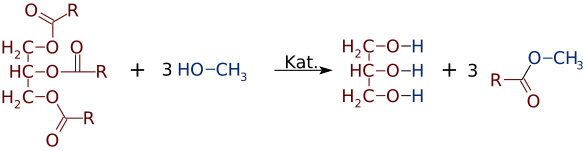
Метиловые эфиры жирных кислот
- 1 year ago
- 0
- 0


Ци́кл трикарбо́новых кисло́т (сокр. ЦТК , цикл Кре́бса , цитра́тный цикл , цикл лимо́нной кислоты́ ) — центральная часть общего пути катаболизма , циклический биохимический процесс, в ходе которого остатки (СН 3 СО-) окисляются до диоксида углерода (CO 2 ). При этом за один цикл образуется 2 молекулы CO 2 , 3 НАДН , 1 ФАД H 2 и 1 ГТФ (или АТФ ) . Электроны , находящиеся на НАДН и ФАДH 2 , в дальнейшем переносятся на дыхательную цепь , где в ходе реакций окислительного фосфорилирования образуется АТФ.
Цикл трикарбоновых кислот — это ключевой этап дыхания всех клеток , использующих кислород , центр пересечения множества метаболических путей в организме , промежуточный этап между гликолизом и электронтранспортной цепью . Кроме значительной энергетической роли циклу отводится также и существенная пластическая функция, то есть это важный источник молекул-предшественников, из которых в ходе других биохимических превращений синтезируются такие важные для жизнедеятельности клетки соединения, как аминокислоты , углеводы , жирные кислоты и др.
Цикл превращения лимонной кислоты в живых клетках (то есть цикл трикарбоновых кислот) был открыт и изучен немецким биохимиком Хансом Кребсом , за эту работу он (совместно с Ф. Липманом ) был удостоен Нобелевской премии (1953 год) .
У эукариот все реакции цикла Кребса протекают внутри митохондрий , а у большинства бактерий реакции цикла протекают в цитозоле .
В начале цикла трикарбоновых кислот ацетил-кофермент А (ацетил-КоА) отдаёт свою ацетильную группу четырёх углеродному соединению — оксалоацетату (щавелевоуксусной кислоте), при этом образуется шестиуглеродный цитрат (лимонная кислота). Ацетил-КоА является продуктом окисления таких соединений, как глюкоза , аминокислоты и жирные кислоты . Цитрат затем изомеризуется в изоцитрат (изолимонную кислоту), который далее дегидрируется и декарбоксилируется до пятиуглеродной кислоты — α-кетоглутарата . α-Кетоглутарат вновь декарбоксилируется, превращаясь в четырёхуглеродный сукцинат (янтарная кислота). Сукцинат затем в три этапа ферментативно превращается в четырёхуглеродный оксалоацетат, который готов прореагировать с новой молекулой ацетил-КоА. В каждый оборот цикла одна ацетильная группа (то есть два атома углерода) приходит в цикл в виде ацетил-КоА, и два же атома углерода покидают цикл в виде двух молекул CO 2 ; одна молекула оксалоацетата используется для образования цитрата, и одна же впоследствии регенерируется. Оксалоацетат не покидает цикл, и одна молекула оксалоацетата теоретически может связывать неограниченное количество ацетильных групп и, на самом деле, оксалоацетат присутствует в клетках в очень низких концентрациях. Четыре из восьми стадий цикла представляют собой окислительные процессы, выделяющаяся при этих процессах энергия окисления эффективно запасается в виде восстановленных коферментов НАДН и ФАДH 2 .

Хотя цикл трикарбоновых кислот занимает центральное место в энергетическом метаболизме, его роль не сводится к получению и запасанию энергии. Четырёх- и пятиуглеродные промежуточные соединения цикла служат предшественниками для синтеза многих соединений. Для восполнения этих промежуточных соединений, покинувших цикл, в клетке существуют специальные .
Как упоминалось выше, все реакции цикла трикарбоновых кислот происходят в митохондриях, и в митохондриях же располагается дыхательная цепь (на внутренней мембране ). У большей части бактерий ферменты цикла трикарбоновых кислот находятся в цитозоле, а плазматическая мембрана выполняет функции, аналогичные функциям внутренней мембраны митохондрий .
Цикл трикарбоновых кислот включает 8 основных стадий, которые подробно рассматриваются ниже.
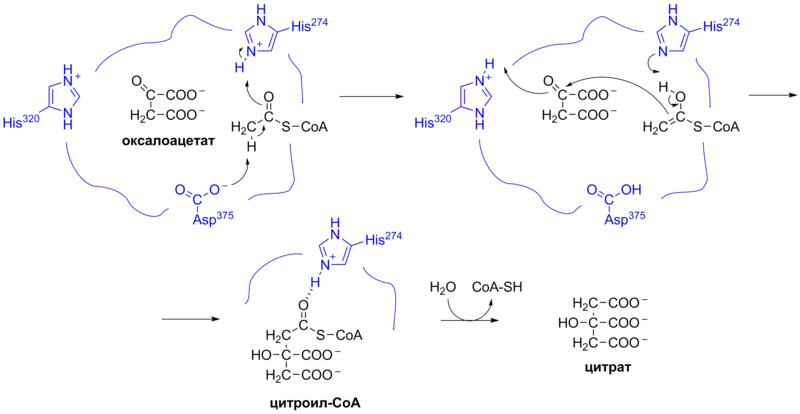
Первой реакцией цикла является необратимая конденсация ацетил-КоА с оксалоацетатом с образованием цитрата , катализируемая ферментом цитратсинтазой (реакция 1 на общей схеме):

В этой реакции метильная группа в составе ацетильной группы ацетил-КоА присоединяется к карбонильной группе (второму атому углерода, C2-атом) оксалоацетата . В ходе этой реакции в активном центре фермента образуется промежуточное соединение — цитроил-КоА . Оно быстро подвергается гидролизу и расщепляется на свободный КоА и цитрат, которые удаляются из активного центра фермента . Гидролиз этого высокоэнергетичного тиоэфирного промежуточного соединения делает эту реакцию весьма экзергонической . Большое отрицательное изменение стандартной свободной энергии цитратсинтазной реакции необходимо для управления циклом, поскольку, как отмечалось ранее, в норме концентрация оксалоацетата в клетке очень мала. КоА, высвобождаемый при этой реакции, далее участвует в окислительном декарбоксилировании следующей молекулы пирувата при помощи пируватдегидрогеназного комплекса .
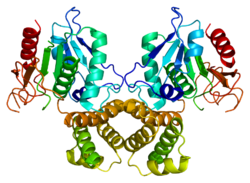
Цитратсинтаза
.png)

Цитратсинтаза была закристаллизована, и был проведён её рентгеноструктурный анализ в присутствии и отсутствии её субстрата и ингибиторов . Каждая субъединица этого гомодимерного фермента представляет собой единый полипептид с двумя доменами , один из которых — крупный и жёсткий, а другой — менее крупный и более пластичный; между этими доменами располагается активный центр фермента. Оксалоацетат — первый из субстратов, связывающихся с цитратсинтазой, — индуцирует значительные конформационные изменения в пластичном домене, создавая сайт связывания для молекулы второго субстрата — ацетил-КоА (см. справа). Когда в активном центре фермента образуется цитроил-КоА, в ферменте происходит второе конформационное изменение, обусловленное гидролизом тиоэфира с высвобождением КоА. Эти изменения конформации, вызванные сначала связыванием с субстратом, потом — промежуточным продуктом, препятствуют преждевременному и непродуктивному разрыву тиоэфирной связи в ацетил-КоА. Кинетические исследования цитратсинтазы подтверждают описанный выше двусубстратный механизм её работы. Вышеописанная цитратсинтазная реакция представляет собой альдольную конденсацию (впрочем, некоторые авторы рассматривают её как конденсацию Клайзена ). Ниже представлен механизм цитратсинтазной реакции:

Фермент аконитаза (точнее, аконитатгидратаза) катализирует обратимую изомеризацию цитрата в изоцитрат через образование промежуточного соединения — трикарбоновой кислоты , которая при этом в норме не покидает активный центр. Аконитаза присоединяет воду по двойной связи связанного с её активным центром цис -аконитата двумя разными способами: в результате одного из них образуется цитрат, в результате другого — изоцитрат (реакции 2 и 3 на общей схеме) :

Хотя в равновесной смеси при pH 7,4 и температуре 25 °С содержится менее 10 % изоцитрата, в клетке реакция смещена вправо, поскольку изоцитрат быстро вовлекается в следующую стадию цикла, и его концентрация уменьшается. Аконитаза содержит железосерный кластер , который служит как для связывания субстрата в активном центре, так и для каталитической гидратации или дегидратации. В клетках, не содержащих достаточного количества железа , аконитаза утрачивает свой железосерный кластер и приобретает регуляторную роль в (подробнее см. IRE (биология) ). Таким образом, аконитаза является одним из многих ферментов, имеющих две различные функции .
Ниже представлена схема, иллюстрирующая, как железосерный кластер аконитазы связывает изоцитрат и преобразует его в цис -аконитат:

В следующей стадии фермент катализирует изоцитрата с образованием α-кетоглутарата (оксоглутарата). Ион Mn 2+ (или Mg 2+ ) в активном центре фермента взаимодействует с карбонильной группой промежуточного соединения , который образуется быстро, однако не покидает активного центра до тех пор, пока не декарбоксилируется и не превратится в α-кетоглутарат .
Ниже подробно рассмотрены эти превращения (реакции 4 и 5 на общей схеме):

В клетках обнаружены две различных формы (изозима) изоцитратдегидрогеназы. Для функционирования одной из них нужен НАД + , для другой — НАДФ + (причём для активности последней нужен ион Mg 2+ , а не Mn 2+ ). Осуществляемые ими реакции в остальном идентичны. У эукариот НАД-зависимый изозим локализован в митохондриальном матриксе и участвует в цикле трикарбоновых кислот. Главной функцией НАДP-зависимого изозима, встречающегося как в митохондриальном матриксе, так и в цитозоле, возможно, является образование НАДФH , который необходим для восстановительных анаболических процессов .
В следующей стадии цикла трикарбоновых кислот также происходит окислительное декарбоксилирование, при котором α-кетоглутарат превращается в сукцинил-КоА и СО 2 под действием ; в качестве акцептора электронов выступает НАД + , а КоА функционирует как переносчик сукцинильной группы. Энергия окисления α-кетоглутарата запасается при образовании тиоэфирной связи в сукцинил-КоА (реакция 6 на общей схеме):

Эта реакция практически идентична пируватдегидрогеназной реакции окислительного декарбоксилирования пирувата, а α-кетоглутаратдегидрогеназный комплекс чрезвычайно близок к пируватдегидрогеназному комплексу (ПДК) по структуре и функциям. Он включает 3 фермента, гомологичных ферментам E 1 , E 2 и E 3 ПДК, и его кофакторами также являются тиаминпирофосфат , липоат , ФАД, НАД и кофермент А. Несомненно, оба комплекса имеют общего эволюционного предка. Хотя ферменты E 1 обоих комплексов структурно схожи, их аминокислотные последовательности различаются и, конечно, они специфичны к разным субстратам: E 1 комплекса ПДК связывает пируват, а E 1 α-кетоглутаратдегидрогеназного комплекса связывает α-кетоглутарат. Ферменты E 2 обоих комплексов также очень похожи, и оба ковалентно связываются с липоатом. Субъединицы E 3 обоих комплексов идентичны .
Сукцинил-КоА, как и ацетил-КоА, содержит тиоэфирную связь с большой отрицательной стандартной свободной энергией гидролиза (ΔG′ о ≈ −36 кДж/моль). В следующей стадии цикла трикарбоновых кислот энергия, выделяемая при расщеплении тиоэфирной связи, используется на образование в ГТФ или АТФ, при этом сукцинил-КоА превращается в сукцинат (реакция 6 на общей схеме):

Эта обратимая реакция катализируется ферментом (сукцинилтиокиназой); из обоих названий этого фермента следует, что в этой реакции участвует нуклеозидтрифосфат .

Эта энергозапасающая реакция включает промежуточные этапы, на которых молекула фермента сама становится фосфорилированной по остатку гистидина в активном центре. Эта фосфорильная группа, которая имеет высокий потенциал для переноса, переносится на АДФ или ГДФ с образованием АТФ или ГТФ соответственно. В клетках животных имеется два изозима сукцинил-КоА-синтетазы, один из которых специфичен к АДФ, а другой — к ГДФ. Сукцинил-КоА-синтетаза состоит из двух субъединиц: α-субъединица (M r = 32 000) содержит фосфорилируемый остаток гистидина (His 246 ) и место связывания КоА, а β-субъединица (M r = 42 000) обеспечивает специфичность связывания с AДФ или ГДФ. Активный центр находится в промежутке между субъединицами. Кристаллическая структура сукцинил-КоА-синтетазы содержит две «силовые спирали» ( англ. power helices ), по одной в каждой субъединице, причём эти спирали ориентированы таким образом, что их электрические дипольные моменты смещают частично положительные заряды к отрицательно заряженному гистидинфосфату (P—His); благодаря этому стабилизируется промежуточная фосфорилированная форма фермента . Ниже представлена схема реакции, катализируемой сукцинил-КоА-синтетазой:

Образование ATФ (или ГТФ) за счёт энергии, запасённой при окислительном декарбоксилировании α-кетоглутарата, является реакцией субстратного фосфорилирования , как и синтез ATФ при гликолизе , катализируемый и пируваткиназой . ГТФ, синтезированный сукцинил-КоА-синтетазой, может отдать свою терминальную фосфорильную группу АДФ с образованием АТФ в ходе обратимой реакции, катализируемой :
Таким образом, конечным результатом активности любого изозима сукцинил-КоА-синтетазы является запасание энергии в виде АТФ. Изменение энергии Гиббса в нуклеозидифосфаткиназной реакции равно нулю, и АТФ и ГТФ энергетически эквивалентны друг другу .
Сукцинат, образовавшийся из сукцинил-КоА, окисляется в фумарат под действием флавопротеина сукцинатдегидрогеназы (реакция 8 на общей схеме):


У эукариот сукцинатдегидрогеназа прочно связана со внутренней митохондриальной мембраной , у бактерий она располагается на плазматической мембране. Этот фермент содержит 3 различных железосерных кластера и одну молекулу ковалентно связанного с ним ФАД, являющегося простетической группой фермента. Электроны от сукцината проходят через ФАД и железосерные кластеры, а далее они в составе переносчиков электронов попадают на дыхательную электронтранспортную цепь, расположенную на внутренней мембране митохондрий (плазматической мембране у бактерий). ФАД при этом восстанавливается до ФАДH 2 , однако дальнейшим акцептором электронов является убихинон . Переход электронов от сукцината через эти переносчики на конечный акцептор электронов — кислород — сопряжён с синтезом АТФ, причём на пару электронов образуется 1,5 молекулы АТФ. Малонат , обычно отсутствующий в клетках, является сильным конкурентным ингибитором сукцинатдегидрогеназы, и добавление этого соединения в митохондрии блокирует активность цикла трикарбоновых кислот .
Обратимая гидратация фумарата с образованием L- малата катализируется ферментом (точнее, фумаратгидратазой ). Переходным продуктом этой реакции является карбанион (реакция 9 на общей схеме):

Более детально механизм фумаразной реакции представлен ниже:

Фумараза является ферментом: она катализирует гидратацию двойной связи в фумарате ( транс -изомер ), но не в малеате ( цис -изомер фумарата). Фумараза также проявляет стереоспецифичность и при осуществлении обратной реакции: D-малат не может служить субстратом для него .
В последней реакции цикла трикарбоновых кислот НАД-зависимый фермент L- малатдегидрогеназа катализирует окисление L-малата до оксалоацетата (реакция 10 на общей схеме):


В стандартных термодинамических условиях равновесие этой реакции сильно смещено влево, однако в живой клетке оксалоацетат постоянно вовлекается в высокоэкзергоническую цитратсинтазную реакцию (стадия 1). Так поддерживается крайне низкая концентрация оксалоацетата в клетке (< 10 −6 М), благодаря чему равновесие малатдегидрогеназной реакции смещается вправо .
Хотя ферменты цикла трикарбоновых кислот обычно описывают как растворимые компоненты митохондриального матрикса (кроме мембраносвязанной сукцинатдегидрогеназы), поступает всё больше доказательств того, что внутри митохондрий эти ферменты существуют в виде мультиферментных комплексов . Ферменты цикла были успешно выделены из экстрактов разрушенных клеток, однако при этом были разрушены мультибелковые комплексы, образованные за счёт одного белка с другим, или со структурным компонентом клетки ( мембраной , микротрубочкой , микрофиламентом ). Однако при приготовлении клеточного экстракта содержимое клеток, в том числе и ферменты, разбавляется в 100 или 1000 раз .
Ряд доказательств говорит о том, что в клетках мультиферментные комплексы обеспечивают эффективный переход продуктов реакций одного фермента к следующему ферменту пути. Такие комплексы называются метаболонами . Несколько ферментов цикла трикарбоновых кислот были изолированы в составе супрамолекулярных комплексов или были обнаружены связанными со внутренней митохондриальной мембраной, или же для них была показана более низкая скорость диффузии, чем для отдельных белков в растворе. Это служит убедительным доказательством обмена субстратами между мультиферментными комплексами и в других метаболических путях, и многие ферменты, считающиеся «растворимыми», в действительности образуют высокоорганизованные комплексы, обменивающиеся промежуточными соединениями .
Выше были рассмотрены реакции, составляющие один оборот цикла трикарбоновых кислот. Двухуглеродная ацетильная группа входит в цикл, соединяясь с оксалоацетатом. Два атома углерода покидают цикл в виде двух молекул СО 2 , образовавшихся при окислении изоцитрата и α-кетоглутарата. Энергия, выделившаяся при этих реакциях окисления, запасается в виде восстановленных трёх молекул НАДН, одной молекулы ФАДH 2 и одной молекулы АТФ или ГТФ. В конце цикла молекула оксалоацетата регенерируется. Стоит отметить, что те два углеродных атома, которые покидают цикл в виде двух молекул СО 2 , отличны от тех двух углеродных атомов, которые поступили в цикл (на этом обороте) в виде ацетильной группы. Атомы углерода, которые принесены ацетильной группой, могут покидать цикл в виде СО 2 только на последующих оборотах цикла .
Хотя в ходе цикла трикарбоновых кислот непосредственно образуется лишь одна молекула АТФ на оборот (при превращении сукцинил-КоА в сукцинат), четыре окислительные реакции цикла обеспечивают дыхательную цепь значительным числом электронов, поставляемых НАДН и ФАДH 2 , и тем самым обеспечивают образование значительного количества АТФ в ходе окислительного фосфорилирования .
В ходе гликолиза из одной молекулы глюкозы образуется две молекулы пирувата, 2 АТФ и 2 НАДН. В ходе окислительного фосфорилирования переход двух электронов с НАДН на O 2 обеспечивает образование 2,5 АТФ, а переход двух электронов с ФАДH 2 на O 2 даёт 1,5 АТФ. Когда обе молекулы пирувата окисляются до 6 СО 2 пируватдегидрогеназным комплексом и в ходе цикла трикарбоновых кислот, а электроны переносятся на O 2 в ходе окислительного фосфорилирования, то суммарный выход АТФ составляет 32 молекулы на молекулу глюкозы :
| Реакция | Выход АТФ или восстановленных коферментов | Суммарный выход АТФ |
|---|---|---|
| глюкоза → глюкозо-6-фосфат | −1 АТФ | −1 |
| фруктозо-6-фосфат → фруктозо-1,6-бисфосфат | −1 АТФ | −1 |
| 2 глицеральдегид-3-фосфат → 2 1,3-бисфосфоглицерат | 2 НАДН | 3 или 5 |
| 2 1,3-бисфосфоглицерат → 2 3-фосфоглицерат | 2 АТФ | 2 |
| 2 фосфоенолпируват → 2 пируват | 2 АТФ | 2 |
| 2 пируват → 2 ацетил-КоА | 2 НАДН | 5 |
| 2 изоцитрат → 2 α-кетоглутарат | 2 НАДН | 5 |
| 2 α-кетоглутарат → 2 сукцинил-КоА | 2 НАДН | 5 |
| 2 сукцинил-КоА → 2 сукцинат | 2 АТФ (или 2 ГТФ) | 2 |
| 2 сукцинат → 2 фумарат | 2 ФАДH 2 | 3 |
| 2 малат → 2 оксалоацетат | 2 НАДН | 5 |
| Итого : 30—32 |
32 молекулы АТФ эквивалентны 32 × 30,5 кДж/моль = 976 кДж/моль, что составляет 34 % от теоретического максимума при полном окислении глюкозы — 2840 кДж/моль. Эти вычисления произведены с учётом стандартных значений изменений свободной энергии, однако, если учитывать реальную нужду клетки в свободной энергии, заключённой в АТФ, то эффективность процесса окисления приближается к 65 % от теоретического максимума .
| Стадия | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| ΔG'°, кДж/моль | -32,2 | 13,3 | -7,1 | -33,5 | -2,9 | 0 | -3,8 | 29,7 |
Регуляция ферментов метаболического пути может осуществляться при помощи аллостерических эффекторов и ковалентных модификаций, поддерживая концентрацию промежуточных и конечных продуктов в клетке постоянной и препятствуя их чрезмерному образованию. Переход углеродных атомов от пирувата к циклу трикарбоновых кислот тонко регулируется на двух уровнях: превращение пирувата в ацетил-КоА, стартовое соединение для цикла (пируватдегидрогеназная реакция), и вход активного ацетата в цикл (цитратсинтазная реакция). Ацетил-КоА образуется не только пируватдегидрогеназным комплексом (PDH), но и также при окислении жирных кислот (β-окисление) и некоторых аминокислот, поэтому для регуляции окисления пирувата и цикла трикарбоновых кислот также важен контроль этих путей. Кроме того, цикл регулируется на изоцитратдегидрогеназной и α-кетоглутаратдегидрогеназной реакциях. Ниже будет рассмотрена регуляция собственно цикла трикарбоновых кислот ; о регуляции окислительного декарбоксилирования пирувата см. статью Окислительное декарбоксилирование пирувата .
Итак, поступление метаболитов в цикл трикарбоновых кислот строго регулируется. Поступления метаболитов определяется тремя факторами: доступностью субстрата, подавлением накапливающимися продуктами и аллостерическим подавлением по типу обратной связи ферментов, катализирующих начальные этапы цикла .
Каждая из трёх экзергонических стадий цикла — стадии, катализируемые цитратсинтазой, изоцитратдегидрогеназой и α-кетоглутаратдегидрогеназой — при некоторых условиях могут стать . Доступность субстратов для цитратсинтазы (ацетил-КоА и оксалоацетата) варьирует в зависимости от состояния клетки и иногда сдерживает скорость образования цитрата. НАДН, продукт окисления изоцитрата и α-кетоглутарата, при некоторых условиях накапливается, и при большом значении отношения [НАДН]/[NAD + ] обе дегидрогеназные реакции строго подавляются. Аналогично, в клетке малатдегидрогеназная реакция находится в строгом равновесии (то есть она лимитируется субстратом), и при большом значении отношения [НАДН]/[NAD + ] и низкой концентрации оксалоацетата первая стадия цикла замедляется. Накопление продуктов ограничивает все три лимитирующие стадии цикла: сукцинил-КоА подавляет α-кетоглутаратдегидрогеназу (а также цитратсинтазу); цитрат блокирует цитратсинтазу; конечный продукт, АТФ, подавляет цитратсинтазу и изоцитратдегидрогеназу. АДФ — аллостерический активатор цитратсинтазы — уменьшает ингибирующее влияние на этот фермент АТФ. В мышечной ткани позвоночных ионы Са 2+ , служащие сигналом к сокращению и сопутствующие увеличению потребности в АТФ, активируют изоцитратдегидрогеназу и α-кетоглутаратдегидрогеназу, а также пируватдегидрогеназный комплекс (PDH). Таким образом, концентрация субстратов и промежуточных соединений в цикле трикарбоновых кислот задаёт такой поток углерода через него, при котором концентрации образующихся АТФ и НАДН будут оптимальны .
В норме скорости гликолиза и цикла трикарбоновых кислот тесно связаны, так что в пируват превращается лишь такое количество глюкозы, которое обеспечит цикл достаточным количеством «топлива» — ацетильными группами ацетил-КоА. Концентрации пирувата, лактата и ацетил-КоА в норме поддерживаются постоянными. Скорость гликолиза связана со скоростью цикла трикарбоновых кислот не только через ингибирование гликолиза высокими уровнями АТФ и НАДН, которое характерно и для гликолиза, и для дыхательного этапа окисления глюкозы, но также и концентрацией цитрата. Цитрат, первый продукт цикла трикарбоновых кислот, является важным аллостерическим ингибитором фосфофруктокиназы-1 , гликолитического фермента .
Восьмишаговый циклический процесс окисления простой ацетильной двухуглеродной группы до СO 2 может показаться излишне сложным и не отвечающим биологическому . Однако роль цикла трикарбоновых кислот не исчерпывается окислением ацетат-иона (а значит, и углеводов, жирных кислот и некоторых аминокислот, при окислении которых он образуется). Этот путь составляет сердцевину метаболизма промежуточных соединений. Четырёх- и пятиуглеродные конечные продукты многих катаболических процессов вступают в цикл на промежуточных этапах. Оксалоацетат и α-кетоглутарат, например, являются продуктами катаболизма аспарагиновой и глутаминовой кислот, которые образуются при расщеплении белков. В некоторых метаболических процессах задействованы многие промежуточные соединения цикла, они служат предшественниками во многих анаболических процессах. Таким образом, цикл трикарбоновых кислот является амфиболическим путём, он связывает катаболические и анаболические процессы .
Пируват является продуктом окисления углеводов. Далее пируват превращается в ацетил-КоА и вовлекается в цикл трикарбоновых кислот. Кроме того, ацетил-КоА является также продуктом окисления жирных кислот, так что цикл трикарбоновых кислот участвует и в катаболизме жиров . Стоит отметить, что пируват может вовлекаться в цикл трикарбоновых кислот и без превращения в ацетил-КоА, а преобразовавшись в малат под действием малик-фермента .
Из α-кетоглутарата, образующегося в цикле трикарбоновых кислот, синтезируются аминокислоты глутамин , глутамат, пролин и аргинин . Сукцинил-КоА выступает в качестве предшественника при синтезе порфиринов и гема . Цитрат участвует в и стеролов (из цитрата образуется ацетил-КоА, кроме того, он выступает в роли регулятора ). Малат может транспортироваться из митохондрий в цитоплазму, где он обратимо превращается в оксалоацетат. Получившийся оксалоацетат может служить предшественником для синтеза аминокислот аспартата, аспарагина , метионина , треонина и изолейцина , а также пиримидинов . Он может также превратиться в фосфоенолпируват с затратой ГТФ, а фосфоенолпируват (ФЕП) может служить предшественником в биосинтезе фенилаланина , тирозина , триптофана , серина , глицина и цистеина . Пируват, получающийся из ФЕП в процессе гликолиза, может стать предшественником аланина , лейцина и валина , а также может участвовать в глюконеогенезе .
Промежуточные соединения цикла трикарбоновых кислот, покинувшие цикл и задействованные в синтезе различных соединений, замещаются за счёт специальных . В нормальных условиях реакции, в ходе которых промежуточные соединения цикла вовлекаются в другие метаболические пути, и реакции, возмещающие их уход, находятся в динамическом равновесии, поэтому концентрация промежуточных соединений цикла трикарбоновых кислот поддерживается постоянной .
Ниже в таблице представлены важнейшие анаплеротические реакции :
| Реакция | Фермент | Ткань/организм |
|---|---|---|
| пируват + НСО 3 − + АТФ ⇌ оксалоацетат + АДФ + Ф н | печень , почки | |
| фосфоенолпируват + СО 2 + ГДФ ⇌ оксалоацетат + ГТФ | фосфоенолпируваткарбоксикиназа | сердце , скелетные мышцы |
| фосфоенолпируват + НСО 3 − ⇌ оксалоацетат + Ф н | фосфоенолпируваткарбоксилаза | высшие растения , дрожжи , бактерии |
| пируват + НСО 3 − + НАД(Ф)H ⇌ малат + НАД(Ф) + | малик-энзим | широко распространена среди эукариот и бактерий |
В печени и почках млекопитающих наиболее важной анаплеротической реакцией является обратимое карбоксилирование пирувата с образованием оксалоацетата, катализируемое ферментом пируваткарбоксилазой . Когда в цитратном цикле уменьшается количество оксалоацетата или других промежуточных соединений, пируват карбоксилируется, образуя дополнительный оксалоацетат. Ферментативное добавление карбоксильной группы к пирувату требует энергии, которая берётся из АТФ: свободная энергия, необходимая для присоединения карбоксильной группы к пирувату практически равна свободной энергии, которую можно получить из АТФ. Пируваткарбоксилаза — регуляторный фермент, и она инактивируется в отсутствие ацетил-КоА — положительного аллостерического модулятора. Когда же ацетил-КоА — «топливо» для цикла трикарбоновых кислот — присутствует в избытке, он стимулирует пируваткарбоксилазную реакцию и тем самым способствует образованию оксалоацетата, что, в свою очередь, даёт возможность вовлекать в цикл трикарбоновых кислот больше ацетил-КоА. Для осуществления пируваткарбоксилазной реакции необходим витамин биотин , выступающий в качестве простетической группы фермента, переносящей СО 2 . Биотин должен присутствовать в рационе человека, он содержится во многих продуктах и синтезируется кишечными бактериями .
Другие анаплеротические реакции, указанные в таблице выше, также регулируются таким образом, чтобы обеспечивать концентрацию промежуточных соединений, достаточную для функционирования цикла трикарбоновых кислот. Например, фосфоенолпируваткарбоксилаза активируется фруктозо-1,6-бисфосфатом — промежуточным соединением гликолиза, которое накапливается в условиях избытка пировиноградной кислоты .
У растений и бактерий в ходе глиоксилатного цикла ацетил-КоА может превращаться в сукцинат. Таким образом, эти организмы могут осуществлять анаплеротическую деградацию нейтральных жиров (подробнее о глиоксилатном цикле см. ниже) .
Существуют и другие анаплеротические пути. Аминокислоты гистидин, пролин, аргинин, глутамин и глутамат могут превращаться в α-кетоглутарат и восстанавливать его концентрацию; изолейцин, валин, метионин, триптофан — в сукцинил-КоА, аспартат, фенилаланин и тирозин — в фумарат; аспартат и аспрагин — в оксалоацетат. Аминокислоты аланин, серин, треонин, цистеин и глицин могут преобразовываться в пируват, необходимый для протекания цикла трикарбоновых кислот .


Как упоминалось выше, неполный цикл трикарбоновых кислот имеется у некоторых анаэробных организмов. У них он служит не для получения энергии, а для получения предшественников для биосинтетических процессов. Эти организмы используют первые три реакции цикла, чтобы получить α-кетоглутарат, однако, лишённые α-кетоглутаратдегидрогеназы, они не могут осуществлять все превращения цикла. У них, однако, имеются 4 фермента, катализирующих последовательное превращение оксалоацетата в сукцинил-КоА, поэтому они могут образовывать малат, фумарат, сукцинат и сукцинил-КоА из оксалоацетата в ходе реакций, обратных «нормальным» (окислительным) реакциям цикла. Этот путь представляет собой брожение , в ходе которого НАДН, образующийся при окислении изоцитрата , переводится в НАД + восстановлением оксалоацетата до сукцината .
У растений , некоторых беспозвоночных и некоторых микроорганизмов (например, дрожжей, Escherichia coli ) ацетил-КоА превращается в сукцинат в ходе глиоксилатного цикла , тесно связанного с циклом трикарбоновых кислот. Общее уравнение глиоксилатного цикла выглядит так:
Образующийся сукцинат далее участвует в биосинтетических процессах. У растений глиоксилатный цикл локализован в особых органеллах — глиоксисомах .
Некоторые бактерии способны осуществлять обратный цикл трикарбоновых кислот . В ходе этого процесса реакции цикла трикарбоновых кислот осуществляются в обратном направлении: там, где в цикл поступают атомы углерода в виде ацетил-КоА и впоследствии окисляются до СО 2 , в обратном цикле, наоборот, выделяется ацетил-КоА. Для его осуществления необходимы доноры электронов, и для этих целей бактерии используют водород , сульфиды или тиосульфаты . К ферментам обратного цикла, отличным от соответствующих ферментов прямого цикла, относятся , 2-оксоглутарат: ферредоксиноксиредуктаза , . Обратный цикл трикарбоновых кислот считается альтернативным фотосинтезу путём образования углеводов .
Цикл трикарбоновых кислот является общим путем окисления ацетильных групп, к которому сводятся практически все метаболические пути живых организмов. Он вовсе не является кратчайшим путём окисления ацетата до СO 2 , однако в процессе естественного отбора оказалось, что он обладает наибольшими преимуществами. Ранние анаэробы, возможно, использовали некоторые реакции цикла трикарбоновых кислот в линейных биосинтетических процессах. В самом деле, некоторые современные анаэробные микроорганизмы используют неполный цикл трикарбоновых кислот, но не как источник энергии, а как источник предшественников для биосинтетических процессов (подробнее см. раздел ). Вместе с эволюцией цианобактерий , образующих O 2 из воды, земная атмосфера становилась аэробной, и под влиянием естественного отбора у организмов развивался аэробный метаболизм, гораздо более эффективный, чем анаэробное брожение .
Когда механизмы регуляции таких путей, как цикл трикарбоновых кислот, нарушаются, то результатом может стать развитие серьёзного заболевания. Ферменты цикла кодируются генами домашнего хозяйства , и отсутствие функциональных копий этих генов может быть объяснено наличием тканеспецифичных особенностей проведения цикла . Среди людей мутации , затрагивающие гены ферментов цикла, очень редки, однако те из них, что происходят, имеют губительные последствия.
Дефекты гена фумаразы приводят к появлению опухолей гладких мышц ( лейомиомы ) и почек ; мутации сукцинатдегидрогеназы вызывают рак надпочечников ( феохромоцитому ). В культурах клеток с такими мутациями накапливаются фумарат (в случае мутаций фумаразы) и, в меньшей степени, сукцинат (в случае мутаций сукцинатдегидрогеназы), и это накопление активирует транскрипционный фактор HIF-1α , индуцируемый гипоксией . Развитие рака может быть следствием состояния псевдогипоксии. В клетках с такими мутациями наблюдается повышенная экспрессия генов , в норме регулируемых HIF-1α. Такие последствия мутаций генов фумаразы и сукцинатдегидрогеназы позволяют относить их к супрессорам опухолей .
Показана связь дефектов фумаразы и нарушений работы нервной системы .
Мутации, изменяющие активность α-кетоглутаратдегидрогеназы, приводят к накоплению в моче продуктов распада аминокислот, из-за чего моча приобретает запах кленового сиропа. Это заболевание называется лейциноз ( англ. Maple syrup urine disease ) .

Несколько соединений и реакций цикла трикарбоновых кислот были открыты в 1930 году Альбертом Сент-Дьёрди , в частности, он установил роль фумарата, ключевого компонента цикла. За свои открытия в 1937 году Сент-Дьёрди был удостоен Нобелевской премии по физиологии и медицине . Полностью последовательность реакций и образующихся соединений в 1937 году установил Ханс Адольф Кребс, за что в 1953 году он получил Нобелевскую премию (совместно с Ф. Липманом) (в его честь цикл трикарбоновых кислот получил одно из своих названий). В 1948 году Э. Кеннеди и Альберт Ленинджер установили, что у эукариот все реакции цикла происходят в митохондриях .
Когда около 60 лет назад стали доступны тяжёлый изотоп углерода 13 C и радиоактивные изотопы 11 C и 14 C, они были использованы для того, чтобы проследить путь атомов углерода в цикле трикарбоновых кислот. Один из таких экспериментов дал весьма неожиданные результаты. Ацетат, меченный по гидроксильной группе, был соединён с немеченым оксалоацетатом с образованием меченого цитрата. Поскольку цитрат — симметричная молекула, то предполагалось, что он будет преобразован в α-кетоглутарат, среди которого будут молекулы, меченые по разным атомам углерода. Тем не менее, из клеток был выделен лишь один «сорт» молекул α-кетоглутарата, и исследователи заключили, что цитрат и любая другая симметричная молекула не может быть промежуточным соединением на пути от ацетата к α-кетоглутарату; они предположили, что в результате конденсации ацетата и оксалоацетата образовывалась несимметричная трикарбоновая кислота, например, цис -аконитат или изоцитрат . В 1948 году Александр Огстон установил прохиральность цитрата (склонность к асимметричным реакциям в отсутствие хирального центра ), объяснив тем самым результаты экспериментов и подтвердив, что именно цитрат образуется в первой стадии цикла .
Для более лёгкого запоминания кислот, участвующих в цикле Кребса, существует мнемоническое правило :
Целый Ананас И Кусочек Суфле Сегодня Фактически Мой Обед , что соответствует ряду — цитрат, цис -аконитат, изоцитрат, альфа-кетоглутарат, сукцинил-CoA, сукцинат, фумарат, малат, оксалоацетат.
Существует также следующее мнемоническое стихотворение (его автором является ассистент кафедры биохимии КГМУ Е. В. Паршкова ):
(щавелевоуксусная кислота, лимонная кислота, цис -аконитовая кислота, изолимонная кислота, α-кетоглутаровая кислота, сукцинил-KoA, янтарная кислота, фумаровая кислота, яблочная кислота, щавелевоуксусная кислота).
Другой вариант стихотворения: