Гаплогруппа R (Y-ДНК)
- 1 year ago
- 0
- 0


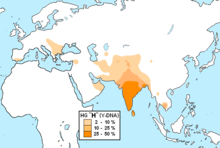
R1b — гаплогруппа Y-хромосомы , наиболее распространённая в Западной Европе и на Южном Урале . Встречается также в Центральной Азии , Восточной Европе , Северной Африке , Западной Азии . После миграций европейцев в Америку и Австралию она составляет значительную долю и там. Определяется однонуклеотидным полиморфизмом M343 , открытым в 2004 году . С 2001 по 2005 год R1b определяли наличием ОНП P25. В старых системах классификации — Hg1 и Eu18 .
Наиболее часто встречается в Западной Европе (Испания, Португалия, Франция, Британские острова, Северная Италия), а также в некоторых частях России — у башкир и в Дагестане. В центральной Африке — Чад и Камерун. В более низком содержании гаплогруппа есть в Восточной Европе, Западной Азии, Северном Иране, в некоторых частях Северной Африки и Центральной Азии (особенно в Северном Туркменистане).
Происходит из гаплогруппы R1 , так же, как и гаплогруппа R1a .
Гаплогруппа R1b возникла от мутации гаплогруппы R1 , произошедшей у мужчины , жившего ок. 22 800 лет назад (дата определена по снипам компанией YFull). Последний общий предок носителей R1b жил 20,4 тыс. лет назад .
Британские генетики Брайан Сайкс и Стивен Оппенгеймер утверждают, что гаплогруппа R1b не имеет отношения к индоевропейскому заселению Западной Европы и происходит от (по их мнению) палеолитического (доиндоевропейского) населения Иберии ( баски ). Большинство исследователей не считает басков палеолитическими. Взгляды Сайкса и Оппенгеймера получили широкое распространение в Европе благодаря написанным ими популярным бестселлерам о генетической истории Европы.
С другой стороны, «европейская палеолитичность» R1b сталкивается с непреодолимыми противоречиями. Последующие исследования Барбары Арреди с коллегами установили, что разнообразие субклад данной гаплогруппы увеличивается по мере движения на восток, что скорее говорит о восточном происхождении данной гаплогруппы и связано с распространением сельского хозяйства из Западной Азии в Европу . Ряд современных генетиков полагают, что R1b зародилась в Центральной или Западной Азии .
Этот вывод подтверждается дальнейшими исследованиями подгруппы R1b1a2, носителем которой, возможно, являлись фараоны Эхнатон и Тутанхамон . Она, предположительно, зародилась на Кавказе около 9500 лет назад и начала миграцию в Европу около 7000 лет назад (см. Балканский неолит ). Часть её представителей мигрировала в Северную Африку (примером массовой миграции из Ближнего Востока в Египет является нашествие гиксосов , которое имело место за полтысячелетия до рождения Тутанхамона ). Ок. 5700—7300 лет назад ( неолитический субплювиал ) из Евразии в район озера Чад пришли носители субклады R1b1a2-V88 . Приход R1b1a2-V88 в Сахару датируется периодом от 8,67 до 7,85 тыс. лет назад, а его основная внутренняя ветвь R1b-V1589 датируется возрастом 5,73 тыс. лет назад (время последнего периода Зелёной Сахары ) .
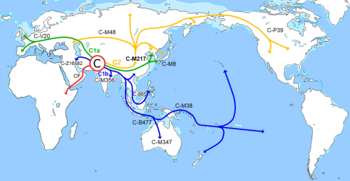
R1b возникла от Y-хромосомной гаплогруппы R1 , которая произошла от гаплогруппы R. Место возникновения гаплогруппы R — Северная Азия . Промежуток — между 26 500 — 19 000 лет назад, в этот период был последний ледниковый максимум . Самая древняя находка Y-хромосомной гаплогруппы R* — останки мальчика MA1 со стоянки Мальта́ в Иркутской области возрастом 24 000 лет . Популяция из Мальты произошла от популяции северо-восточных сибиряков (Ancient North Siberians, ANS), представленных обитателями Янской стоянки с Y-хромосомной гаплогруппой P1 , предковой для Y-хромосомных гаплогрупп Q и R .
Носители гаплогруппы R внесли большой вклад в происхождение современных жителей Европы и Южной Азии . В этих регионах гаплогруппа R наиболее распространена — R1b в Западной Европе , R1a в Восточной Европе , Центральной и Южной Азии и R2 в Южной Азии.
Три основные субклады R1b1 — R1b1a, R1b1b, R1b1c происходят с Ближнего Востока . Южная ветвь R1b1c — R1b1a2-PF6279/V88 обнаруживается в Леванте и Африке . Северная ветвь R1b1a — R1b1a1a-P297/PF6398 возникла в районе Кавказа , восточной части Анатолии и возможно в Северной Месопотамии . Затем носители R1b1a1a-P297 двинулись в Европу и Центральную Азию , разделившись на две ветви.

Взрывообразный рост числа потомков основателя субклады произошёл примерно 5,9—4,8 тыс. л. н., что совпадает с распространением культуры колоколовидных кубков в Западной Европе и культуры боевых топоров в Восточной Европе .
R1b1a2-V88 определили у образца I5235 (9221—8548 лет до н. э., Железные Ворота , Padina, Сербия ) . R1b1a2b1-Y8451 определили у образца R6 (7160 (7192—7127) л. н., Апулия, Италия ) . У представителей субнеолитической льяловской культуры ( Московская область ) со стоянки Сахтыш IIA (4993-4791 лет до н. э.) определили Y-хромосомные гаплогруппы R1b .
R1b1a2-V1636 (ISOGG2019-2020) определили у энеолитического образца ART038 (3356—3121 лет до н. э.) из Арслантепе ( Турция ) , у энеолитических образцов PG2001 и PG2004 (4233—4047 лет до н. э.) со стоянки Прогресс-2 ( Ставропольский край ), у представителя ямной культуры SA6010 (2884—2679 лет до н. э.) из могильника Шарахалсун-6 (Sharakhalsun 6) в Ставропольском крае , у поздненеолитического по археологической хронологии образца Gjerrild 5 (SGC/LN, 2283—2035 лет до н. э.), найденного в могильнике культуры одиночных могил ( Дания ) .

Современные лингвисты помещают место зарождения праиндоевропейцев в Понтийско — Каспийские степи. Они простираются от Устья Дуная до Уральских гор и до Северного Кавказа . Культуры неолита , энеолита и раннего бронзового века в Понтийско Каспийской степи были названы единой курганной культурой , которая развивалась между 6200 — 4200 лет назад, данное определение было дано Марией Гимбутас в рамках её Курганной теории происхождения индоевропейцев . В современный период данная теория почти отвергнута, в силу того, что курганные захоронения датируются не позже 6000 лет назад и возникли на Юге Кавказа . Генетическое разнообразие R1b больше в Восточной Анатолии , этот факт говорит о том, что эволюционировали носители R1b именно там и южнее, и лишь затем пришли на Северный Кавказ и попали в Понтийско Каспийские степи .
R1b-P297 сформировалась 15 600 л. н. (95%-й доверительный интервал 17 600 — 13 800 л. н.) .
Лошади были одомашнены около 6600 лет назад в Каспийской степи или в районе Дона или в районе Нижней Волги . Этот прорыв стал определяющим для степных культур. Однако R1b не одомашнивали лошадей, они пришли в Понтийско-Каспийские степи, когда эти животные уже были поставлены на службу людям. Наиболее вероятно, что лошадей одомашнили носители гаплогруппы R1a , вкупе со старой ветвью R1b-P297, немногочисленной по сравнению с R1a. R1b-P297 ещё ни в коем случае не была индоевропейской, это были люди ещё жившие в ледниковом периоде и не знавшие скотоводства. Эта ветвь поселилась в Понтийско-Каспийских степях в позднем палеолите , что подтверждается находками в Самаре и Латвии . R1b-P297 и R1a по составу геному, были почти чисто коренными людьми, периода мезолита, жившими в Восточной Европе . В них имелась небольшая примесь от сибирских охотников палеолита и мезолита, но отсутствовала примесь от жителей Кавказа (CHG). При этом примесь жителей Кавказа найдена у культур халколита — афанасьевоской , ямной и культуры шнуровой керамики .
Гаплогруппа R1b1a1a2-M269 сформировалась в позднем палеолите 13 300 л. н. (95%-й доверительный интервал 14 900 — 11 800 л. н.) . Время до самого недавнего общего предка (TMRCA) потомков R1b1a1a-M269 составляет 6400 л. н., а непосредственно вышестоящего узла R1b-P297 — намного раньше 13 300 л. н. Y-хромосомная гаплогруппа R1b-P297(xM269) обнаружена у охотников-собирателей из Прибалтики, а также у охотников-собирателей из Самарской области России. «Длинная ветвь» R1b1a1a2-M269 сильно снижает любую уверенность в близости самых ранних носителей R1b1a1a2-M269 к этим восточноевропейским родственникам. Тем не менее, данные в равной степени согласуются со сценарием, в котором у основателя R1b1a1a2-M269 не было компоненты * .
Точный маршрут миграции R1b1a1a2-M269 от Южного Кавказа до Понтийско-Каспийской степи пока не ясен. Возможно это было в период днепро-донецкой культуры , 7 100 — 6 300 лет назад. Данная культура стала первым обществом уровня неолита в Прикаспийской степи, у днепро-донецких носителей R1a и/или I2a1b, коровы и козы уже паслись в степи и появились похоронные ритуалы, они так же импортировали медь и медные изделия с территории Балкан и Карпат . Часть своих культурных традиций они переняли у охотников периода мезолита . Знания о домашних животных пришли к данной культуре от земледельцев с Ближнего Востока , это были носители I , I2a2 , R1a , R1b1a (L754) и один R1b1a2-L388. Что важно — среди этих земледельцев ещё не было носителей ветвей R1b1a1a2-M269 или нижестоящей R1b-L23 — которые затем доминировали в ямной культуре. МтДНК днепро-донецкой культуры представлена исключительно женщинами европейского происхождения периода мезолита — U4a, U4b, U4d , U5a1, U5a2, U5b2 , а также одна J2b1 и одна U2e1, все эти мтДНК гаплогруппы не индоевропейские, а доиндоевропейские. Вплоть до периода бронзового века , индоевропейские мтДНК не обнаруживаются в образцах периода неолита, найденных на территории Украины . Таким образом, днепро-донецкая культура , которая уже имела и скотоводческие знания и земледельческие и являлась самой ранней подобной культурой в Понтийской степи, не относится ни к индоевропейской культуре ни к протоиндоевропейской культуре .
Типично индоевропейскими мтДНК являются — H2a1, H6, H8, H15 , I1a1 , J1b1a , W3, W4 и W5 , эти гаплогруппы женской линии будут обнаруживаться в ямной культуре и унетицкой культуре и затем уже в других индоевропейских культурах .
Первыми протоиндоевропейскими культурами были хвалынская (7200 — 6500 лет назад) и среднестоговская (6600 — 5900 лет назад). В этот период появляются небольшие захоронения курганного типа, с характерным положением мёртвых на спине, с поднятыми коленями, ориентированными на северо-восток. В этих культурах население активно вступало в связи с другими племенами, это заметно по разнообразным формам черепов. 7000 лет назад элитная прослойка этих культур начала активное развитие с использованием стад крупного рогатого скота , лошадей и меди. Медь была именно статусным металлом, дорогим и относительно трудно добываемым и обрабатываемым. На рубеже периодов хвалынской и среднестоговской культур, появилась от R1b1a1a2-M269 ветвь R1b-L23, это произошло примерно 6400 л. н. (95%-й доверительный интервал 7100 — 5700 л. н.) . В итоге в современный период 99 % индоевропейской R1b ведёт свой корень от этой ветви — R1b-L23. R1b-L51 и R1b-Z2103 отошли от R1b-L23 6100 л. н. От R1b1a1a2-M269 6400 л. н. так же произошла другая клада R1b-PF7562 , но она немногочисленна и представлена на Балканах, в Турции , Армении ; на территории современной Греции она появилась в эпоху поздней бронзы .
Вторая миграция через Кавказ произошла около 5700 лет назад и совпадает с возникновением майкопской культуры , первой в мире культуры бронзового века. Наиболее вероятно, что носители Майкопской культуры родом из Северной Месопотамии , Сирии и Восточной Анатолии . На Южном Кавказе в Грузии и Азербайджане обнаружена лейлатепинская культура (6350 — 6000 лет назад). Керамика майкопской и лейлатепинской культур имеет схожие черты, так же похожа на их керамику посуда из поселений куро-аракской культуры . Важно, что археологические находки показывают явное распространение курганных погребений с бронзовыми изделиями из майкопской культуры в Понтийской степи, именно в этой степи вскоре возникал ямная культура — около 5500 лет назад. Курганный тип погребений стал доминирующими является одним из маркеров для определения принадлежности культуры к индоевропейской. Курганы как способ погребения знати затем использовались кельтами, римлянами, германскими племенами, скифами .
Между 5 500 — 4 500 лет назад активно развивалась ямная культура бронзового века. Она считается бесспорно индоевропейской [ источник не указан 861 день ] и наиболее важной в основании индоевропейской культуры и общества. R1b-M269 носители с Ближнего Востока в ямной культуре относительно активно вступали в отношения с земледельцами и пастухами-скотоводами R1a на протяжении более чем 2-х тысяч лет и так же после ямной культуры, в других сообществах. Эти контакты между R1a и R1b не носили спорадического характера, а были плотными и многоуровневыми, как на уровне знати, так на уровне простых людей и торговцев. Вероятно, это привело к постепенному формированию своеобразной общности населения ямной культуры, у ямной культуры появился относительно единый язык, который принято называть праиндоевропейским , который стал впоследствии основой для современных индоевропейских языков . Большинство современных лингвистов сошлись во мнении, что именно регион Понтийско-Каспийских степей является местом возникновения праиндоевропейского языка. Это подтверждается сходством праиндоевропейского языка с кавказскими и хурритскими языками, ведущими своё начало с Ближнего Востока и с другой стороны — с уральскими языками . Только в Понтийско-каспийской степи исторически могли пересекаться так плотно пути носителей этих языков, что бы в итоге возник праиндоевропейский язык. .
Ямная культура наиболее чётко характеризуется тем, что люди её составлявшие использовали телеги на четырёх колёсах, для того чтобы менять место жительства, перевозили на них еду и домашние принадлежности. Подобный транспорт обеспечивал возможность длительных походов в степях, что делало ямников одним из самых мобильных народов, при этом занимавшимся и земледелием и скотоводством одновременно. Этот набор умений положил начало великим миграциям индоевропейцев. В более поздние времена, в период Древнего Рима , галлы и древние германцы так же вели свои миграции .
Ямная культура, учитывая её большие размеры, не была единой. На юге вдоль северного побережья Чёрного моря до Северного Кавказа была полноценная степь, почти без леса, эта степь простиралась дальше, до Каспийского моря , до Сибири и до Монголии и в англоязычной литературе называется Евразийской степью.
1 — Западный ареал ямной культуры, между реками Дон и Днестр и затем Дунаем , был ареалом проживания в основном R1b, а носителей R1a было там не больше 5 — 10 % [ источник не указан 861 день ] .
Таким образом Западная часть ямников впоследствии мигрировала на Балканский полуостров и в Грецию . Затем в Центральную , а уже в относительно поздние исторические периоды — обратно, в виде волн миграций, состоявших из хеттов , фригийцев и армян , эти волны достигли Анатолийского полуострова .
2 — Восточная часть ареала ямной культуры, от Волги до Уральских гор была населена R1a , и так же уже с меньшинством носителей R1b. Следы от R1b остались в генетике некоторых родов башкир , у туркмен , уйгуров и хазарейцев .
Эта ветвь в дальнейшем мигрировала в Центральную Азию , в Синьцзян Уйгурский регион и в Южную Азию — Иран , Пакистан , Индия .
Майкопская культура (развивалась между 5 700 — 4 500 лет назад) — имела ареал на Северо Западе Кавказа . В культурном развитии была тесно связана с южным ареалом ямной культуры . Жители Северного Кавказа в тот период имели тесные контакты с жителями Понтийско-каспийских степей . Это подтверждается многочисленными находками керамической посуды, медного и бронзового оружия и ювелирных украшений. При этом торговля между культурами на Северном побережье Чёрного моря и Северным Кавказом, старше периода Майкопской культуры. Уже между 6 400 — 5 700 лет назад люди торговали между этими регионами. Суворовско-новоданиловская культура, имевшая ареал на севере Чёрного моря контактировала с Северным Кавказом и со среднестоговской культурой . Поселение на месте Нальчика , существовавшее между 7000 — 6500 лет назад, имело схожую с хвалынской культуру . При этом хвалынская культура распространялась в степях рядом с Каспийским морем и в Поволжье . Именно на этот ранний период приходится время взаимодействия и начала смешивания между R1b и R1a .
Жители ямной и майкопской культур схожи в системе погребений — обе культуры использовали курганный тип. Покойников помещали в лежачем положении с поднятыми коленями с ориентацией по оси Северо Восток — Юго Запад. Могилу обильно посыпали красной охрой , рядом с человеком хоронили принесённое в жертву домашнее животное. У этих культур были одомашненные лошади , колёсные повозки и телеги, в домашнем хозяйстве было большое количество скота. Овец в тот период держали не для мяса, а для получения шерсти. В качестве оружия пользовались медными и бронзовыми топорами и кинжалами. Важно — наиболее древние находки колёсных повозок и бронзовых изделий найдены именно на Северном Кавказе и скорее всего распространялись в Понтийско-каспийских степях именно оттуда.
Майкопская культура была одной из передовых в бронзовом веке и судя по современным находкам — первой в мире, где начали разрабатывать месторождения меди и изобрели выплавку меди и металлообработку как таковую. В итоге эта культура первой на Земле освоила оружие из металла. Самый древний меч найден в майкопской культуре, на объекте Курган 31. Стиль меча напоминает стиль длинных кельтских мечей , но менее сложный по внешнему виду. В курганах майкопцев так же находятся кости лошадей и изображения этих животных. Это говорит о возможно преемственности между майкопцами и культурой в Понтийско-каспийских степях. Однако есть ряд кардинально различных культурных особенностей между этими культурами, что говорит за гибридную основу Майкопской культуры. Сложность в определении генетики жителей этой культуры состоит в том, что население жило там в небольших селениях и в соседних сёлах могли быть носители анатолийской R1b, кавказской G2a и степной R1a . За гибридное развитие майкопцев и ямников говорит то, что типичная кавказская генетическая линия — G2a-L13 встречается по всей Европе , а также в Центральной Азии и Южной Азии . На Балканах , в бассейне реки Дунай и в Центральной Европе распространение G2a-L13 коррелирует по процентам с R1b.
Предполагается, что так называемые Народы моря — это западная ветвь индоевропейцев R1b, которая находилась тогда у побережья Чёрного моря и совпадает по срокам развития и миграций с народами моря. Образование Хеттской державы , с явно индоевропейским языком в Центральной Части Анатолийского полуострова произошло спустя несколько столетий после исчезновения майкопской и ямной культур. Принимая во внимание, что большая часть индоевропейской R1b в Анатолии относится к подкладу R1b-Z2103, нет сомнений в том, что хетты пришли в Анатолию с территории Балканского полуострова. До их миграции, ещё существовавшие культуры ямная и майкопская вторглись в Юго-Восточную Европу. Именно миграция носителей R1b, которые ушли в Юго-Восточную Европу и Анатолию, носители R1a-Z282 смогли продвинуться из северной части ямной культуры южнее и заняли берега Чёрного моря. Это событие связывают с возникновением Срубной культуры (3 600 — 3 200 лет назад), срубная культура R1a заняла ареал ямной и майкопской культур, заполнив таким образом образовавшийся «вакуум».
В конце неолита — примерно 5 000 лет назад, R1b пересёк Кавказ и разделился на две ветви. Западная ветвь R1b-L51 поселилась на востоке и севере Чёрного моря. Восточная ветвь R1b-Z2103 продвинулась Дона и до Поволжья, в регионы где были одомашнены лошади уже около 6 600 лет назад. Восточная ветвь R1b-Z2103 в этом регионе смешалась с носителями R1a и в итоге зародилась репинская культура (сущ. 5 700 — 5 300 лет назад). Эта культура не намного, но древнее ямной культуры, которая зародилась южнее. Уже из репинской культуры, используя уже и лошадей, R1b продолжили миграции и вместе с некоторыми мигрантами из западной ветви L51 двинулись на восток, в глубину Евразийского континента, идя по степям. Они дошли в итоге до Алтая и основали афанасьевскую культуру. (сущ. 5 600 — 4 400 лет назад). Лингвисты склонны указывать, что афанасьевцы подходят на роль предшественников тохарской языковой ветви индоевропейских языков. В 2014 году анализ ДНК у трёх образцов Y-ДНК, показал, что все они — R1b, включая два R1b-M269. [ источник не указан 942 дня ] В 2020 году в афанасьевской культуре в Западной Монголии обнаружен образец Y-ДНК западной ветви R1b-L51 > L52
Те носители R1b, которые остались жить на Волге и около Урала, по всей видимости основали со временем полтавкинскую культуру (сущ. 4 700 — 4 100 лет назад). Затем полтавкинская культура соединилась и растворилась в синташтинской и петровской культурах (сущ. 4 100 — 3 750 лет назад). В синташтинской культуре доминировала гаплогруппа R1a , это связывают с завоеваниями индо-арийцев в Центральной и Южной Азии. В современный период R1b в России встречается среди небольших народов Волги и Урала — удмурты , коми , мордва , татары . Среди русских R1b распространена незначительно. Так же R1b заметно представлена в Центральной Азии: уйгуры — до 20 %, ягнобцы в Таджикистане — до 32 %, узбеки — до 21,4 % (Кашкадарьинская обл.), туркмены — до 37 %, среднее значение для Башкир — 47 % , доминирует у башкирских родов бурзян и гайна (87 %).
R1b1a1a1-Y13200/M73 определена у неолитического образца VK531 из норвежского Тромсё (2400 лет до н. э.) .

Когда индоевропейцы освоили бронзовое оружие и научились использовать повозки , запрягаемые лошадьми , их мобильность резко возросла. Они стали активно мигрировать в Европу, где жили в тот период автохтонные, коренные на тот момент культуры, носители C1a2 , F и I (в том числе несшие прямые генетические связи с кроманьонцами ). Так же в Европе, в период начала индоевропейской экспансии жили неолитические фермеры — земледельцы , которые мигрировали в период раннего неолита и несли гаплогруппы G2a , H2 , E1b1b и T1a . Индоевропейцы R1a и R1b, имея явный перевес как в мобильности, так и в вооружении, сумели в относительно короткий срок полностью заменить мужские роды у старых культур Европы, скорее всего большую часть мужчин они убили или отстранили от возможности зачатия новых детей. При этом женская часть этих культур пострадала значительно меньше, что можно объяснить тем, что пришлые носители R1a и R1b активно брали местных женщин в жёны или наложницы.
Если привести в пример Южную Азию , то выводится следующие — после индо-иранского (Арийского) вторжения в Северной Индии стало 40 % мужских линий R1a, но при этом лишь 10 % женских линий имеют индоевропейский корень, остальные женские линии остались коренными. Воздействие экспансии индоевропейцев на Европу было значительно более сильным, в силу меньшей развитости Европы 4000 лет назад, по сравнению с очень развитой в тот период Южной Азией, где уже активно развилась Индская Цивилизация . В Европе 4000 лет назад, ещё не было бронзового оружия, земледелие было менее успешным. В Западной Европе сельское хозяйство появилось значительно позже, чем на Балканах и в Центральной Европе .
Греция , Балканы и Карпаты наоборот, были передовыми в Европе регионами по развитию сельского хозяйства 4 000 лет назад и в итоге их генетические линии меньше всего подверглись воздействию индоевропейцев. Генетические гаплогруппы Старой Европы смогли сохраниться относительно цельно лишь в горных регионах, где было неудобно передвигаться на лошадях — Альпы , Динарские Альпы, Аппенины и Сардиния .
Анализ ДНК носителей культуры шнуровой керамики указывает, что уже 4700 лет назад её носители имели в Польше как гаплогруппу R1a, так и R1b, но не ямный субклад .
Продвижение R1b шло в более развитую часть Европы, населённую аграрными культурами и потому шло медленнее. 4500 лет назад западная ветвь R1b набрала силы для новой сильной миграции в сторону современной Германии и Западной Европы. К этому моменту носители R1b сильно смешались с жителями бассейна Дуная , жившими там до их прихода, которые были охотниками уровня мезолита и земледельцы неолита .
R1b были явно патриархальным сообществом . Они вытесняли местные мужские линии из культур, заменяя их на R1b и активно брали в жёны женщин этих культур. Это привело к тому, что кожа протоиндоевропейцев стала темнее и сравнялась по оттенку с жителями современной Южной Европы , поскольку жители культур неолита были выходцами со Среднего и Ближнего Востока и имели тёмные волосы и тёмный оттенок кожи
R1b-L51 продвинулись в Центральную Европу ( Венгрия , Австрия и Богемия ) 4500 лет назад. Именно к этому времени, спустя 2000 лет после появления здесь культур неолита — земледельческих, их уровне развития достиг высокого уровня, они стали добывать золото и медь и в итоге стали намного более желанной добычей для завоевателей
На данным момент ставится под сомнение, что традиция колоколовидных кубков (4800 — 3900 лет назад) в Западной Европе, была полностью индоевропейской. Поскольку она преемственна именно местных европейским культурам, более старым, которые относятся к мегалитическим культурам . В период позднего неолита и раннего халколита Колоколовидные кубки стали распространяться со стороны Португалии на Северо Восток, в направлении Германии. Степные культуры периода бронзового века наоборот шли от Германии в стороны Франции, Иберии и Великобритании и привнесли постепенно R1b на территорию культуры колоколовидных кубков. Найденные в колоколовидной культуре образцы индоевропейской посуды и фигурки лошадей, скорее всего были не результатом индоевропейской составляющий в этой культуре, а результатом торговли с индоевропейскими носителями. Именно после этой торговли, начался резкий рост миграции и завоеваний со стороны индоевропейских культур и к 2200 г. до н. э. они достигли Атлантического побережья. Это может быть связано с тем, что колоколовидная культура, когда вела торговлю, принесла индоевропейцам информацию о том, что в Западной Европе живут племена, которые ещё не знают металла и находятся на уровне каменного века — палеолита и потому являются лёгкой добычей как сами, так и территории, на которых они живут .
ДНК тесты у носителей культуры колоколовидных кубков показали, что у них есть R1b-L51 (и более древние субклады — P312 и U152). У более ранних культур до колоколовидной таких привнесений нет. Носители колоколовидной культуры, которые жили в Испании и Португалии не имеют примеси от ямной культуры и не несли в себе кроме R1b-L51 типичные гаплогруппы мегалитических культур , которые жили в Европе задолго до индоевропейцев — G2a , I2a1 , I2a2, а также R1b-V88, но эта ветвь не индоевропейская, а более древняя. [ источник не указан 942 дня ] Культуры мегалита в Великобритании принадлежали исключительно к Y-гаплогруппе — I2 (в основном I2a2 и I2a1b-L161), но затем были полностью заменены R1b-L51 в период раннего бронзового века, что говорит о крайне интенсивной миграции её носителей и по сути тотальном завоевании и замещении местных жителей э
Гальштатская культура (между 3200 — 2750 лет назад) считается первой культурой кельтов в Европе — она быстро расширила ареал на Францию, Великобританию, Иберию, Север Италии и долину реки Дунай . Гальштатская культура несла с собой новый язык — кельтский , но технологии бронзы уже были распространены до неё .

Прото итало-кельто-германские племена достигли Германии около 4500 лет назад. Франция была ими заселена 4200 лет назад. Великобритания — 4100 лет назад. Ирландия — 4000 лет назад. Иберия — 3800 лет назад. Это была первая волна расселения и она характеризуется линией — .
Первые ветви R1b пришедшие во Францию — R1b1a1a2a1a2c1-L21 и . 3700 лет назад эти линии достигли нынешней Испании . При изучении древних Y-хромосом, подтвердилась активная экспансия R1b с выдавливанием мужского коренного населения. В Иберии люди неолита и халколита были обладателями гаплогрупп I , I2a1 и G2a . А в бронзовом веке все мужчины уже были R1b (один M269 и два P312), но по мтДНК, наследуемой по женской линии, эти мужчины были сыновьями коренных иберийских линий — H1 , U5b3 , X2b . Развитие бронзовой культуры в Иберии шло довольно медленно и окончательно бронзовый век наступил около 3300 лет назад в культуре полей погребальных урн , которая расширилась от Германии до Каталонии . После неё активно развивалась и расширяла ареал гальштатская культура (3200 — 2750 лет назад) — она заняла весь Иберийский полуостров. К этому времени Иберия уже была связана с Западной Европой сложной торговой сетью .

Начиная с 3300 лет назад вокруг Альп существовала область развития металлургии бронзы , которая заложила в себе основу для образования классической кельтской культуры. Культуры в этой области создали преемственность — от культура полей погребальных урн , которая развилась в гальштатскую культуру начиная с 3200 лет назад и затем культура Гальштата развилась в латенскую культуру — с 2450 лет назад. Это развитие являет собой вторую волну экспансии носителей R1b в Европе, которая затронула Центральную Европу и продвигалась к Атлантическому океану, на Север Скандинавии и на восток к долине реки Дунай. Затем вторая волна дошла до Греции, Украины и России .
Кельтский железный век с 800 года до н. э. мог иметь начало от степей рядом с Северным Кавказом — кобанская культура , существовавшая между 3100 — 2400 лет назад. Кельты населявшие Альпы имели мутацию S28 (другие названия — U152, PF6570)
Гальштатская культура связана с мутацией S28. Данная культура в дальнейшем эволюционировала в культуру Вилланова (между 3100-2700 лет назад). Эта культура уже полностью индоевропейская, кремированные останки люди этой культуры помещали в урны в виде двойного конуса, захоронения разделились, на простые и на захоронения богатых людей и знатных людей. Знатные захоронения содержали ювелирные украшения, бронзовые доспехи и конскую упряжь. В этой культуре в том числе появились типичные индоевропейские символы — свастика. За культурой Вилланова последовали этруски , демонстрирующие прямую преемственность с этой культурой. У этруссков, живших в Центральной Италии от 800 года до н. э. до 1 года до н. э., с частотой 70 % выявлена Y-хромосомная гаплогруппа R1b (R1b-P312 и R1b-L2), с частотой 20 % — Y-хромосомная гаплогруппа G2, а также — Y-хромосомная гаплогруппы J2 (5 %). Во времена Римской империи генетический вклад в популяцию этрусков от людей, имеющих происхождение из Восточного Средиземноморья увеличился до около 50 % .

Основной ветвью прото германцев является — R1b-S21 (он же U106 или M405). [ источник не указан 942 дня ]
Анатолийская ветвь
С Балканского полуострова скорее всего пришли фригийцы Они пришли в Малую Азию — Анатолию около 3200 лет назад в период « великих потрясений » в Восточном Средиземноморье. Фригийцы основали королевство 3200 лет назад, оно просуществовало 500 лет и находилось в Центральной части Анатолии, в западном её сегменте. Это королевство вобрало в себя большую часть разрушавшейся империи хеттов. Прото-армяне пересекли Кавказ, дошли до озера Ван и обосновались в Армянском нагорье . В современный период 30 % армян это носители R1b с субкладой Z2103. Большая часть R1b в современной Греции это балканская разновидность Z2103. [ источник не указан 942 дня ] . В Болгарии и Албании распространён субклад R1b-Z2106.
Анализ шести образцов ДНК из Восточных районов ямной культуры в Волго - Уральском регионе, показал, что все они принадлежат R1b. Четыре из них мутация — Z2103. Это указывает на то, что скорее всего R1b-Z2103 был основной мужской линией полтавкинской культуры , данная культура пришла на смену ямной культуре в этом регионе. Затем полтавкинская культура слилась с абашевской (основная мужская линия в ней — R1a-Z93) и в итоге образовалась синташтинская культура . В итоге в Башкирии , Туркменистане и Уйгурском автономном регионе сохранились заметные очаги R1b-Z2103.
У современного населения Башкирии R1b S28 (U152) встречается у 70 % (в племени Гайна) людей в этой республике. Так же высока концентрация на границе Казахстана и Киргизии. Это была миграция населения в период Античности и Средневековья. Кельты при миграции вдоль Дуная , создали Галатское королевство . R1b-S28 так же является доминирующей ветвью на Аппенинском полуострове , в том числе в период Древнего Рима. Скорее всего римские купцы, при походах по Великому шёлковому пути , существовавшему уже 2200 лет назад, могли оставлять потомство от местных женщин в том числе и в регионе Центральной Азии и Китае. Небольшой процент R1b есть среди населения христианских общин в Ливане и скорее всего он остался от крестоносцев . [ источник не указан 942 дня ]
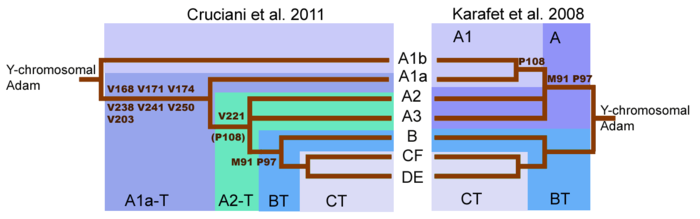
Возраст R1 оценивался Татьяной Карафет (2008) на уровне от 12 500 до 25 700 лет назад, и, скорее всего, обособление этой гаплогруппы произошло около 18 500 лет назад. Доисторические человеческие останки, определённые как принадлежащие к R1b, включают:
Субклад R1b1a найден у представителя культуры Лепенски-Вир . У мезолитчика I1734 из Василевки-на-Днепре (Украина), жившего 7446—7058 лет до н. э., определён субклад R1b1a2-V88>PF6287>PF6362. Субклад R1b1 обнаружен у неолитчиков Дериевки , живших 7500—6800 лет назад . Мезолитический охотник-собиратель, живший 7500 лет назад в Поволжье на реке Сок (Самарская область) и представитель хвалынской культуры , живший 6700 лет назад, были носителями гаплогруппы R1b1 . Субклад R1b1c был обнаружен у обитателя стоянки в Пиринеях (Северный Арагон), жившего около 7100 лет назад.
У представителя неолитической нарвской культуры из Звейниеки определён субклад R1b1a1a-P297* (родительская и для европейской ветви М269 ( ), и для азиатской ветви М73) .
Субклад R1b1a1a2a2-CTS1078/Z2103 обнаружен у представителя энеолитической вучедольской культуры .
Гаплогруппа R1b была обнаружена у одного представителя культуры полей погребальных урн из саксонской пещеры Лихтенштейн в Германии (около 3000 лет назад, поздний бронзовый век), у представителей культуры колоколовидных кубков (около 2800—2000 лет до н. э.) , куро-аракской , ямной (Z2103), межовской культур, у представителей средней ( Nerquin Getashen ) и поздней ( Kapan ) бронзы Армении .
Редкая азиатская ветвь R1b1b-PH155, то есть сестринская для упомянутой R1b1a-L754 линия, определена у двух таримских мумий .
Ветвь R1b1a1a2a1a2a1-Z195 (дочерняя к DF27), выявленная у представителей эль-аргарской культуры , практически полностью заместила энеолитические линии Y-хромосомы .
Поскольку самый ранний пример данной гаплогруппы датирован не позднее 14 000 лет назад и принадлежит R1b1a-L754, то R1b должен был возникнуть относительно скоро после появления R1.
R1b является подклассом внутри макро-гаплогруппы K-M9 , которая является одной из преобладающих гаплогрупп за пределами Африки . Место возникновения гаплогруппы K — Азия. Татьяна Карафет (2014) утверждает, что: "быстрый процесс диверсификации K-M526, вероятно, произошел в Юго-Восточной Азии, с последующим распространением на западе предков гаплогрупп R и Q .
От субклада K2b гаплогруппы K выделилась гаплогруппа P-M45 (K2b2) , а от неё произошла гаплогруппа P1 — непосредственный предок гаплогруппы R , из которой в свою очередь выделилась гаплогруппа R1 .
Три генетических исследования в 2015 году оказали поддержку курганской гипотезе Марии Гимбутас относительно прародины индоевропейцев. Согласно этим исследованиям, гаплогруппы R1b и R1a, теперь наиболее распространенные в Европе (R1a также распространена в Южной Азии), были распространены из Западной Евразийской степи вместе с индоевропейскими языками. Также обнаружили аутосомный компонент, присутствующий в современных европейцах, которого не было у неолитических европейцев, который был привнесен с отцовскими линиями R1b и R1a, а также с индоевропейскими языками .
Как заметили Барбара Арреди и её коллеги, распределение R1b в Европе образует клин с Востока на Запад, что соответствует теории проникновения её носителей в Европу из Западной Азии с распространением земледелия . Дальнейшие исследования 2009 года Chiaroni et al., указывает на то, что гаплогруппа R1b имеет наиболее старые субклады в Западной Азии и наиболее поздние в Западной Европе.
При этом исследователи не отрицают того, что гаплогруппа R1b могла попасть в Европу в период неолита, то есть в более позднее время относительно прошлых оценок, в связи с миграциями земледельцев с Ближнего Востока .
.jpg)
В 2016 году, генетики Монголии и Южной Кореи, провели исследования останков тел, найденных на территории Таван Толгой — это месторождение каменного угля на юге Монголии , в пустыне Гоби , в аймаке Умнеговь . Монгольские археологи, участвовавшие в раскопках могил Тавана Толгоя, предполагали, что 5 из 7 могил Тавана Толгой принадлежали Золотой семье, и одна из этих 5 могил считалась принадлежащей монгольской королеве . Проведены исследования в мтДНК (женская) и Y-ДНК (мужская). Члены семьи являлись носителями гаплогруппы R1b-M343 . Изначально генетики предполагали, что сам Чингисхан и его семья были носителями Y- гаплогруппы C3c-M48 , которая распространена в высокой концентрации среди современных монголов, казахов, эвенов, эвенков, долганов . Люди, которых исследовали, могли произойти от браков между династией Борджигинов (к этой династии принадлежал Чингисхан ) и кланами Онгуд или Хонгирад .


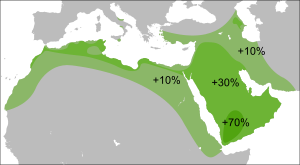
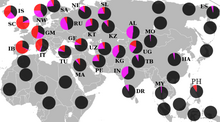
Современная концентрация R1b максимальна на территориях, связанных с кельтами: в южной Англии около 70 %, в северной и западной Англии, Уэльсе , Шотландии , Ирландии — до 90 % и более, в Испании — 70 %, во Франции — 60 % . По-видимому она связана с докельтским субстратом, поскольку высока её концентрация и у не-кельтов басков — 88,1 % [ источник не указан 1329 дней ] и испанцев — 70 % . Кроме того, известно, что, например, строителями Стоунхенджа в Англии было население, обитавшее на острове до прихода кельтов.
У соседних народов концентрация данной гаплогруппы падает: у итальянцев — 40 % , немцев — 40 % , норвежцев — 25,9 % и других.
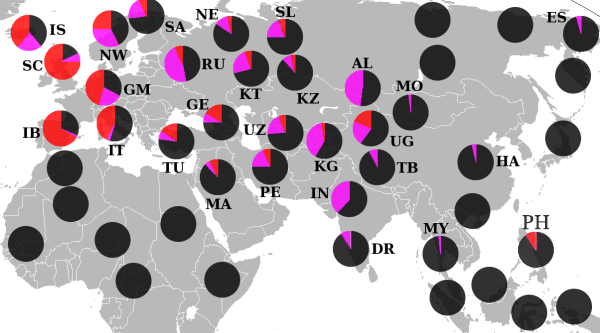
У народов Восточной Европы она встречается ещё реже. У чехов и словаков — 22,6 % [ источник не указан 1329 дней ] , удмуртов — 21,8 % , поляков — 12,6 % , латышей — 10,5 % , венгров — 17,3 % , татар — 14,7 % [20] , эстонцев — 9 % , литовцев — 5 % , белорусов — 4,2 % , русских — 7 % [ источник не указан 1329 дней ] , украинцев — 5,2 % [ источник не указан 1329 дней ] .
На Балканах — у греков — от 13,5 % , словенцев — 21 % , албанцев — 17,6 % [ источник не указан 1329 дней ] , болгар — 17 % , хорватов — 11,7 % , румын — 13 % , сербов — 9,6 % , герцеговинцев — 3,6 % , боснийцев — 2,4 % .
На Кавказе найдена у табасаран 45 %, лезгин 21,5 %, кумыков 25 % и лакцев , аварцев (2,5 %), андийцев, даргинцев (2 %), кумыков, а также жителей багулалы , осетин -дигорцев — по разным данным до 43 % у армян — 25-30 % , азербайджанцев — 15,2 % .
За пределами Западной Европы высокая концентрация данной гаплогруппы встречается лишь у некоторых популяций башкир ( бурзян , гайна и др.) — до 87 % , что, возможно, связано с присутствием дотюркского субстрата, а могут башкирские варианты, наоборот, быть исконной тюркской гаплогруппой, пришедшей из Азии. У башкир высокие показатели как M269, так и M73 [ источник не указан 2489 дней ] .
На Алтае гаплогруппа R1b (субклад R1b1a1a1-Y13200/M73) — встречается у кумандинцев — 49 % .
У Ассирийцев достигает более 40%.
В Турции достигает 16 % , Ираке — 11,3 % и в других странах Западной Азии. В Турции у турок — 21 % .
В Центральной Азии обнаружена, в частности, у следующих тюркских народов: у туркменов — 37 % , узбеков — 30 - 65 % , казахов (в основном у кыпшаков — 47 % )— 5,6 % , уйгуров — 17,2 % , киргизов 1,9 % .
В Пакистане — 6,8 % , в Индии незначительна — 0,55 % .
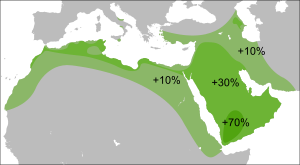
У народа фульбе и у носителей чадских языков доминирует субклад R1b1a2 (PF6279/V88). У камерунского народа мандара R1b достигает 65 % , у алжирских арабов из Орана — 10,8 % , тунисских арабов — 7 % , алжирских берберов — 5,8 % , в Марокко — около 2,5 % , у арабов Египта — от 1 % до 7 % . Аномально высокая для Африки концентрация гаплогруппы R1b, найденная у одной изолированной популяции коренного населения Северного Камеруна, как полагают, возникла в результате доисторического перемещения в Зелёную Сахару древнего евразийского населения.
| Ключевые понятия |

|
|
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
|
Формирование славянских народов — генофонд, история
|
|||||||
|---|---|---|---|---|---|---|---|
| Основное | |||||||
| Генофонд славян |
|
||||||
|
Древнеславянские
археологические культуры |
|
||||||
|
|
Некоторые
внешние ссылки
в этой статье
ведут на сайты, занесённые в
спам-лист
|