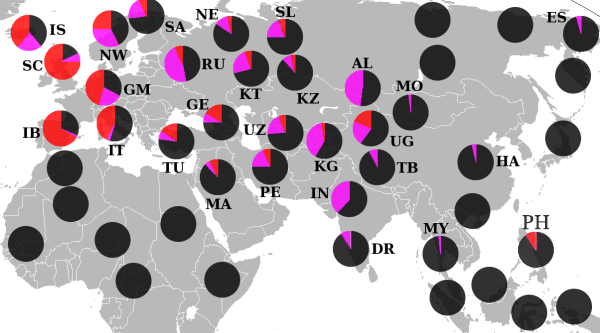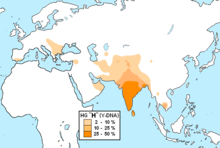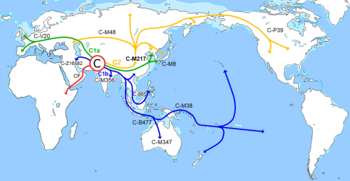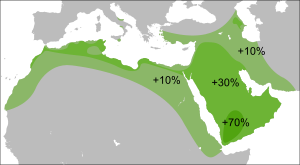
Гаплогруппа R (Y-ДНК)
- 1 year ago
- 0
- 0

Гаплогруппа I2 — гаплогруппа Y-хромосомы . Наивысшей концентрации достигает у жителей Боснии и Герцеговины , Сербии и Хорватии .
До 2008 года была известна под названием I1b . Имеет две субклады — I-L460 и I-L1251.
Гаплогруппа I2 происходит от мутации гаплогруппы I , произошедшей у мужчины, жившего ок. 27 500 лет назад. Сама гаплогруппа возникла между 26 000 — 31 000 лет до нашей эры. Последний общий предок современных носителей гаплогруппы I2-Y283911 жил 21 600 лет назад (даты определены по снипам компанией YFull ).
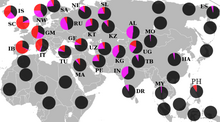
Встречается практически на всей территории современной Европы , максимальной частоты достигает в Динарском нагорье. Мужчины там имеют рекордный для мировой статистики средний рост — 185.6 см.
I2 * (I-M438), если говорить о крупных гаплогруппах, значительно представленных в генофонде европейцев, является самой древней гаплогруппой Европейского континента. По всем признакам она также является автохтонной гаплогруппой Европы, возникшей именно в границах данного континента. Такой же автохтонно-европейской гаплогруппой является C1a2 — но данная ветвь очень слабо представлена в генофонде европейцев.
Изначально с Ближнего Востока в Европу пришли люди, нёсшие гаплогруппу IJ — 35 000 лет назад [ источник не указан 611 дней ] . В процессе миграций населения она постепенно превратилась в гаплогруппу I .
Гаплогруппа I2 (M438/P215/S31) возникла в период позднего палеолита, во время последнего ледникового максимума, между 26 500 — 19 000 лет назад. Определить место или регион в Европе, где произошла гаплогруппа — невозможно, поскольку в тот период все Homo sapiens были охотниками-собирателями — кочевниками, которые постоянно меняли места своих стоянок. Самый старый мужчина-носитель I2 — грот Бишон в Швейцарии , он был представителем азильской культуры , мтДНК у него была U5b1h . К 2016 году проанализировано 15 образцов мужской Y ДНК периода мезолита. Среди этих образцов, один был C1a2 (Испания), один — F (Германия), двое — I (Франция) и шесть образцов — I2 (Люксембург и Швеция), в том числе к I2a1 (P37.2), I2a1a1a (L672), I2a1b (M423) и I2c2 (PF3827) , мтДНК образцов — U2e, U4, U5a1, U5a2 и U5b . Четыре образца из России принадлежали Y-хромосомные гаплогруппам — J, R1a1 и R1b1a . Уже на стадии мезолита заметно сильное разнообразие мужских и женских гаплогрупп, но на сегодняшний день C1a2, F, I, J — чрезвычайно мало представлены в генофонде европейцев. В этом своеобразном конкурентном соревновании гаплогруппа I2a1 вышла «победителем», но причины данной «победы» пока не объяснены научно [ источник не указан 941 день ] .
Анализ образцов раннего неолита в Анатолии в работе Mathieson et al. (2015) показал, что I2c-L596 существовала 8500 лет назад на территории современной Турции, но носители этой гаплогруппы в итоге были ассимилированы земледельцами периода неолита на пути из Плодородного полумесяца в Европу. В итоге в период неолита все гаплогруппы мезолита в Европе попали под массивную ассимиляцию многочисленными земледельцами неолита, пришедшими с Ближнего Востока. Но даже после этого I2a выделяется на общем фоне европейских гаплогрупп как очень распространённая (второе место — 16 % из 69 образцов периода неолита). Первое место — G2a — это линия, прямо происходящая от земледельцев, пришедших с территории Анатолии [ источник не указан 941 день ] .
I2a распространились в старчево-кришской культуре (сущ. между 8000 — 6500 лет назад) и до сих пор являются основной мужской ДНК в Юго Восточной части Европы. В целом I2 активно продолжила развиваться на территории Европы в период медного и бронзового веков. Находки обнаруживаются в Испании — I2a2a-M223, I2a2, I2a2a1 (CTS616) . На Юге России (Ямная культура I2a2a1 (CTS9183) ), в Венгрии (культура Ватья I2a1 ), в Германии (Унетицкая культура I2a2b (L38) и I2c2 ), в Северной Италии (культура Ремеделло I2a1a1 ) [ источник не указан 941 день ]
Концентрация гаплогруппы снизилась, когда в Европу пришли индоевропейцы . В настоящее время гапплогруппа I2 остаётся одной из наиболее распространённых в Европе, уступая Y-хромосомным гапплогруппам R1a и R1b .
На данный момент известно об определении гапплогруппы I2 у представителей большого количества различных археологических культур:
Примечание: на протяжении первого десятилетия XXI века классификация гаплогрупп неоднократно уточнялась и пересматривалась, поэтому в литературе и некоторых статьях википедии могут быть использованы устаревшие названия.
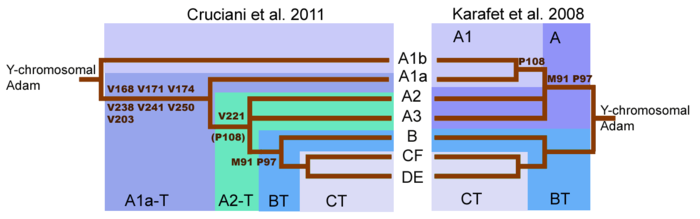
Приведённая ниже схема основана на исследовании ISOGG 2019 , в котором обновлены данные прежней работы Т. М. Карафет и др. (Karafet et al. 2008). Более ранние схемы опубликованы в ISOGG 2007 и ISOGG 2006 .
I2 (L68, M438/P215/S31) (ранее I1b)
Гаплогруппа I2a-M26/L158 "Sardinian" (старое название I2a1) составляет около 40 % всех мужских линий на Сардинии (I2a1a1 ~ 39% ). Кроме того, она также обнаружена с низкой или умеренной частотой в Стране Басков и на Иберийском полуострове , а также составила 1,6 % (1/64) в выборке албанцев , живущих в Македонии и 1,2 % (3/257) в выборке чехов . Также данная гаплогруппа встречается на Британских островах .
Предполагается, что ветвь M26 возникла около 18200 лет назад, а последний общий предок всех современных носителей данного варианта жил около 11700 лет назад.
I2a-M423 был типичен для населения Европы доиндоевропейского завоевания (см. раздел " Палеогенетика "). Согласно данным компании YFull субклад образовался за 18 200 лет до настоящего времени , расхождение субклады на ветви произошло около 13 400 лет назад. В настоящий момент состоит из следующих ветвей:
Субклад I2a-L161.1 "Isles" встречается практически исключительно на Британских островах , в Ирландии , и по всей Великобритании , с очень низкой частотой - в континентальной Европе. Согласно данным компании YFull образовалась за 10 600 до настоящего времени , расхождение субклад произошло около 7900 лет назад.
I2a1b2-L621/S392 возникла между 11 400 л. н. Последний общий предок современных носителей гаплогруппы I2a1b2-L621 жил 6 500 лет назад (даты определены по снипам компанией YFull) .
Субклад I2a-L621 встречается в основном в славянских странах. Максимальная частота наблюдается у динарских славян - словенцы, хорваты, боснийцы, сербы, черногорцы и македонцы. Так же высока частота в Болгарии, Румынии и Молдавии, на Западной Украине и в Белоруссии. В меньшей степени распространена в Албании, Греции, Словакии и в Польше. Незначительно представлена на юго-западе России. L621 в настоящее время встречается в пять - десять раз чаще, чем гаплогруппа G2a в Юго-Восточной Европе. Напротив, в период неолита G2a была в четыре раза более распространённой гаплогруппой. Подобная смена позиций объясняется индоевропейской миграцией и неразрывно связанными с ними славянскими миграциями населения. В частности, последним исторически зафиксированным рывком в этом распространении являются миграции славян между 6 - 9 веками нашей эры. Большая часть восточно-европейских славян это носители субклада L147.2 (он же CTS10228, CTS2180 или Y3111) "Dinaric" который возник около 5100 лет назад, незадолго до начала экспансии людей из ямной культуры . Но данный субклад имеет общего предка (TMRCA) около 3800 лет назад. И около 1900 лет назад I2a1b - L147.2 резко стал увеличивать свою численность, что совпадает по срокам с окончательным формированием славян. Когда готы , вандалы и бургунды из Восточной Германии и в Польше вторглись в Римскую Империю, славяне смогли заполнить образовавшийся этнический вакуум и заселили эти территории. К 476 году, когда распалась Западная Римская империя, славяне расселились по Динарским Альпам и Балканам . К IX веку нашей эры славяне сформировали современные славяноязычные территории. [ источник не указан 941 день ]
Субклад I2a-Disles (Dinaric Isles) - парафилический субклад, в который включают все субклады I2a-L621, кроме I2a-CTS10228 "Dinaric". Данный субклад с очень низкой частотой встречается на Британских островах и во Франции .
Субклад I2a1a2b1a1-CTS10228 (ранее I2a1a2b - Din, в ISOGG2017 - I2a1b2a1a) известен как «Динарская» гаплогруппа (Dinaric). Также этот субклад определяется снипами CTS10228/V4023, CTS5966, CTS2180, CTS5996, FGC12075/YP194, FGC12076/YP217, FGC12077/YP193, FGC12078/Y3107, FGC12081/SK1237/V2129/Y3111, FGC12082/Y3774, FGC12084/Y3775, FGC12096/Y3121, L147.2, S12235, S15187/YP191, S17076/YP188, S17790/YP223, S19015 (ISOGG 2017). Субклад I2a1b2a1a-CTS10228 возник 5 100 л. н. Последний общий предок современных носителей гаплогруппы I2a1b2a1a-CTS10228 жил 3 400 лет назад (даты определены по снипам компанией YFull ).
I2a1b2a1a-CTS5966, CTS2180, CTS5996, CTS10228/V4023 распространён по всей юго-восточной Европе ( Греция , Македония , Сербия , Хорватия , Словения , Босния и Герцеговина , Албания , Черногория , Чехия , Словакия , Польша , Болгария , Белоруссия , Россия , Украина , Венгрия , Австрия , Германия , Швейцария , Румыния , Италия , Молдавия , Литва , Латвия ), а также найдена в Фенноскандии ( Финляндия , Швеция , Норвегия ) и в западной Анатолии . Среди «Динарской гаплогруппы» разделяют 2 кластера: разнообразный северный Динарский кластер (Dinaric North cluster - "Din N"); и южный Динарский кластер (Dinaric South cluster - "Din S") - крупнейшую Динарскую ветвь (около 40% всех представителей Din), которая характеризуется снипом I2a1b2a1a1a3-PH908. Наиболее высокая плотность CTS10228 — в Далмации ( Хорватия ) и Боснии-Герцеговине (>50 %) . Расстояние до времени жизни общего предка субклады I2a-L147.2/CTS5966/CTS10228 — 2757±404 года .
Распространение субгаплогруппы I2a1b1 хорошо коррелирует с распространением гаплогруппы I1 , за исключением Фенноскандии , что позволяет предположить её наличие как минимум в одном из палеолитических убежищ, где также находилась гаплогруппа I1 . Отсутствие корреляции между I1 и I2a1b1 в Фенноскандии может быть связано с тем, что на гаплогруппу I2a1b1 в наиболее ранних поселениях региона сильнее повлиял « эффект основателя » и генетический дрейф из-за её редкости, поскольку гаплогруппа I2a1b1 составляет менее 10 % от всего Y-хромосомного разнообразия населения на указанной территории за пределами Нижней Саксонии .
Распространение гаплогрупп I1 и I2a1b1 относительно хорошо коррелирует с историческими границами распространения носителей германских языков . Гаплогруппа I2a1b1 обнаружена более чем у 4 % населения лишь в Германии , Нидерландах , Бельгии , Дании , Англии (кроме Уэльса и Корнуолла ), Шотландии , на южной оконечности Швеции и Норвегии , а также в провинциях Нормандия , Мэн , Анжу и Перш на северо-западе Франции , в Провансе на юго-востоке Франции , в исторических областях Италии — Тоскане , Умбрии и Лациуме ; а также в Молдавии ; в России — Рязанской области и в Мордовии . Представляется достаточно вероятным, что присутствие I1 и I2b1 в нынешних Франции , Англии и Италии , а также в восточной Европе связано уже с кельтской и германской экспансией , а в доиндоевропейское время эти гаплогруппы концентрировались лишь в северной Европе . Одна из субклад гаплогруппы I2a1b1 , а именно I2a2a1a1-M284, обнаружен почти исключительно у населения Великобритании , что может говорить о его длительной истории существования на Британских островах . Интересно, что с небольшой частотой гаплогруппы I1 и I2b встречаются на территории исторических регионов Вифиния и Галатия в современной Турции , куда их могли принести кельты , мигрировавшие туда по приглашению Никомеда I Вифинского .
Гаплогруппа I2a1b1 также встречается примерно у 1 % населения Сардинии . Дивергенция субклада для мутации I2a2a-M223 произошла около 14,6±3,8 тысяч лет назад (Rootsi 2004). Возраст вариации YSTR для субклада I2a2a-M223 составляет 13,2±2,7 тысяч лет (Rootsi 2004) или 12,3±3,1 тысяч лет (Underhill 2007).
Гаплогруппа I2a1b2a - L38/S154 была обнаружена в скелетных останках, найденных в пещере Лихтенштейн — археологическом памятнике бронзового века в центральной Германии , где также были найдены артефакты культуры полей погребальных урн . Из 19 мужских останков пещеры у 13 была обнаружена гаплогруппа I2a1b2a , у одного — R1b , и у двоих — R1a . Предположительно, пещера находилась в центре тогдашнего распространения гаплогруппы I2a1b2 .
| Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||||||||||||||||||||||||||
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||||
| A0-T | |||||||||||||||||||||||||||||||||
| A00 | A0 | A1 | |||||||||||||||||||||||||||||||
| A1a | |||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||
| F1 F2 F3 | |||||||||||||||||||||||||||||||||
| G | |||||||||||||||||||||||||||||||||
| H | IJK | ||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||
| I | J | K2 | |||||||||||||||||||||||||||||||
| L (K1a) | T (K1b) | K2a/K2a1/ NO /NO1 | |||||||||||||||||||||||||||||||
| N | O | P (K2b2) /P1 | |||||||||||||||||||||||||||||||
| S (K2b1a) | M (K2b1b) | Q | R | ||||||||||||||||||||||||||||||
| Ключевые понятия |

|
|
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||
|
Формирование славянских народов — генофонд, история
|
|||||||
|---|---|---|---|---|---|---|---|
| Основное | |||||||
| Генофонд славян |
|
||||||
|
Древнеславянские
археологические культуры |
|
||||||