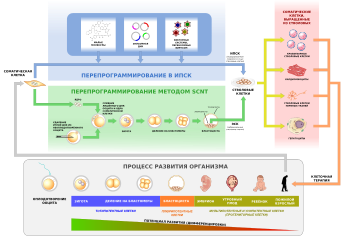
Онтогенез начинается от одной клетки — зиготы.
Зигота
и каждый из
бластомеров
эмбриона
на ранней стадии являются тотипотентными с потенциалом развития в целый организм. По мере развития потенциал бластомеров последовательно снижается сначала до плюрипотентных, затем до мультипотентных и унипотентных и, в конце концов, до терминально дифференцированных соматических клеток. Тем не менее, потенциал развития соматических клеток может быть восстановлен до тотипотентной стадии методом пересадки ядер, взятых из соматических клеток, в яйцеклетку, из которой предварительно удалено ядро (SCNT)
или перепрограммированием до плюрипотентного состояния — например, с помощью факторов Яманаки (Oct4, Klf4, Sox2 и c-Myc)
, РНК
или малых молекул
.
В настоящее время существует три пути перепрограммирования соматических клеток в плюрипотентные стволовые клетки
:
пересадка ядер, взятых из соматических клеток, в оплодотворённую яйцеклетку, из которой предварительно удалено ядро
слияние соматических клеток с плюрипотентными стволовыми клетками
;
модификация соматической клетки, индуцирующая её превращение в стволовую клетку, с помощью: генетического материала, кодирующего
; рекомбинантных белков
; микроРНК
, синтетической самореплицирующейся полицистронной РНК
и низкомолекулярных биологически активных веществ
.
Содержание
Природные процессы индукции
Ещё в 1895 году
Томас Морган
, удалив один из двух
бластомеров
лягушки, обнаружил, что оставшаяся часть
эмбриона
способна, тем не менее, воссоздать цельный эмбрион. Это означало, что клетки, при необходимости, способны изменять направление своей специализации и такое изменение скоординировано. Позднее в 1924 году, Шпеманн и Мангольд (Spemann and Mangold) показали, что важнейшую ключевую роль в процессах развития животных играют межклеточные взаимодействия называемые индукцией
.
Метаплазией
называют обратимую замену одного дифференцированного типа клеток на другой тип зрелых дифференцированных клеток
. Этот переход от одного типа клеток к другому может быть частью нормального процесса созревания или вызван каким-то индуцирующим его стимулом. Примерами этого перехода можно назвать трансформацию клеток радужной оболочки глаза в линзу в процессе созревания и превращение клеток пигментного эпителия
сетчатки
в нейрональную сетчатку при регенерации глаза у взрослых тритонов. Этот процесс позволяет организму заменить исходные клетки, не подходящие к новым условиям, на новые которые больше подходят к новым условиям.
В опытах на клетках
имагинальных
дисков дрозофилы было обнаружено, что существует ограниченное число стандартных дискретных состояний дифференцировки и клеткам приходится выбирать одно из них. Тот факт, что трансдетерминация (смена пути дифференцировки) часто происходит не в одной, а сразу в группе клеток доказывает, что она вызвана не мутацией, а именно индуцирована
.
К настоящему времени удалось выявить минимальные условия и факторы, наличия которых достаточно для индукции каскада молекулярных и клеточных процессов, направляющих дифференцировку и самоорганизацию плюрипотентных клеток в
эмбрион
. Роль
морфогенов
, как оказалось, выполняют противоположно направленные градиенты концентрации
морфогенетического белка костной ткани (BMP)
и белка
.
В основе
вегетативного размножения
растений
лежит
соматический эмбриогенез
, в ходе которого из соматической клетки путём индукции
фитогормонами
образуются тотипотентные клетки дающие начало образованию нового организма без полового процесса.
Некоторые типы зрелых, специализированных клеток взрослого организма
позвоночных животных
также способны естественным путём вернуться к стадии стволовой клетки
. Например, дифференцированные клетки желудка, называющиеся аделоморфными или «главными клетками» и синтезирующие маркер стволовых клеток Troy, обычно производят пищеварительные жидкости. Однако, они могут при необходимости превратиться обратно в стволовые клетки для «ремонтных работ» в случае травм желудка, таких как порез или повреждение вызванное инфекцией. Более того, они осуществляют этот переход даже в отсутствие заметных травм и способны восполнить пул всех клеточных линий желудочного эпителия, по существу, выступая в качестве покоящихся «резервных» стволовых клеток
. При повреждении трахеи, дифференцированные эпителиальные клетки дыхательных путей могут вернуться к фенотипу стабильных и функциональных стволовых клеток, если, однако, они не имеют непосредственного контакта с базальной стволовой клеткой, которая предотвращает подобную дедифференцировку
. Зрелые терминально дифференцированные эпителиальные клетки почки, после травмы, способны дедифференцироваться в свои более ранние версии, а затем снова дифференцироваться в типы клеток нуждавшихся в замене в повреждённой ткани
. Макрофаги могут самообновляться путём локальной пролиферации зрелых дифференцированных клеток
. Это происходит, когда понижаются концентрации или происходит ингибирование двух факторов транскрипции
и
, препятствующих активации программы самообновления
. У
тритонов
мышечная ткань восстанавливается из специализированных
мышечных клеток
, которые для этого дедифференцируются забыв свою прежнюю специализацию. Эта способность к
регенерации
тканей не уменьшается с возрастом, что вероятно связано со способностью тритонов при необходимости образовывать из мышечных клеток новые стволовые клетки
.
В организме существует небольшой процент стволовых клеток, способных генерировать множество различных типов клеток. Например,
(
англ.
) стволовые клетки взрослого человека обладают способностью к самообновлению и образуют в суспензионной культуре характерные скопления (кластеры) плюрипотентных клеток, которые могут дифференцироваться как in vitro, так и in vivo в энтодермальные, эктодермальные и мезодермальные клетки
. Они также легко перепрограммируются в ИПСК
.
Подробное описание некоторых других хорошо документированных примеров
in vivo и их роль в развитии и
регенерации
рассматриваются в обзоре
.
Система отбора факторов плюрипотентности
В самом начале XXI века были обнаружены
белки F-box
один из которых
активен в раннем эмбрионе и только в недифференцированных клетках. Позднее он для поддержания жизни уже не нужен. Это позволило использовать его в качестве маркера плюрипотентности и разработать систему отбора факторов плюрипотентности (названных факторами Яманаки).
]]
Индуцированные тотипотентные клетки обычно используют для
клонирования
и получения генетически модифицированных животных
. Эти клетки можно получить с помощью перепрограммирования соматических клеток путём
переноса ядер соматических клеток
(Somatic cell nuclear transfer — SCNT) в ооциты-реципиенты
. При этом ооциты не обязательно должны принадлежать тому же виду. Иногда удаётся использовать ооциты других видов, например овец
или поросят
. И хотя эффективность межвидовой SCNT была примерно в три раза ниже обычной, такие эмбрионы удавалось довести до стадии
бластоцисты
. Эффективность перепрограммирования можно повысить в два раза, если за сутки до пересадки остановить мейоз ооцитов-реципиентов с помощью бутиролактона1 в комбинации с
нейротрофическим фактором мозга
(BDNF)
. Кроме того, эффективность клонирования может быть значительно повышена, а процедура SCNT упрощена благодаря использованию ингибиторов
гистондеацетилазы
, таких как трихостатин А
и ингибиторов полимеризации
цитоскелетного
актина
таких как цитохалазин В или латранкулин A (latrunculin A)
. Для полноценного развития образующихся эмбрионов необходимо также предварительно уменьшить метилирование лизина 4 в молекуле гистона H3 в клетках донорах ядра
. Кроме того для полноценного развития эмбриона необходимы также
экзосомы
выделяемые его клетками.
Повторное клонирование на протяжении 25 поколений жизнеспособных мышей с помощью метода SCNT, основанном на добавлении в среду клеточной культуры ингибитора
деацетилазы гистонов
— трихостатина А,
показало, что можно достаточно долго (на протяжении 16 лет) неоднократно повторно клонировать животных без видимого накопления нарушений в геноме
.
До настоящего времени бытует представление о возможности преждевременного старения клонированных животных, полученных методом SCNT. Показано, что
теломеры
у эмбрионов клонированных свиней, полученных с помощью стандартных методов SCNT хуже восстановлены, по сравнению с эмбрионами образующимися по естественному пути. Обработка же трихостатином А значительно увеличивает длину теломер у клонированных свиней и это может быть одним из механизмов, лежащих в основе улучшенного развития клонированных животных, после обработки трихостатином А
.
При использовании технологии SCNT разработанной Миталиповым
можно получать ЭСК человека используя ядра из фибробластов кожи даже пожилых людей, что открывает широкие перспективы для технологий
регенеративной медицины
Разработан метод, открывающий новые возможности для создания генетически модифицированных животных с помощью
гаплоидных
эмбриональных стволовых клеток, которые могут быть использованы вместо спермы. Для этого из ооцита удаляют ядро. Затем в него вводят микроинъекцией сперму. Из образующейся в результате этого бластоцисты получают гаплоидные эмбриональные стволовые клетки. Эти клетки, синхронизованные в М фазе, вводят в ооцит вместо спермы, в результате чего развивается жизнеспособное потомство
. Эти разработки, вместе с данными о возможности неограниченного получения ооцитов из митотически активных половых стволовых клеток
, открывают возможность промышленного производства трансгенных сельскохозяйственных животных. Так, в Китае с помощью упрощённой техники клонирования получены трансгенные овцы, у которых улучшено качество мяса и молока за счёт увеличения в них незаменимых ненасыщенных жирных кислот, которые снижают риск развития ишемической болезни сердца и необходимы для поддержки глаз и головного мозга. Ген, вызывающий синтез ω-3 полиненасыщенных жирных кислот успешно удалось передать трансгенной овце.
Клонирование животных для исследовательских целей в Китае уже приобрело промышленные масштабы. Одних только различных клонов поросят производится порядка 500
.
Подобные технологии могут также найти клиническое применения для преодоления цитоплазматических дефектов в ооцитах человека
. Например, разработаны технологии, которые могут воспрепятствовать нежелательному наследованию митохондриального заболевания, которое передаётся следующему поколению. Митохондрии, которые часто называют «электростанцией клетки», содержат генетический материал, который передаётся от матери к ребёнку. Мутации митохондриальной ДНК могут вызвать диабет, глухоту, заболевания глаз, желудочно-кишечные расстройства, болезни сердца, деменцию и ряд других неврологических заболеваний. Пересадкой ядра из яйцеклетки одного человека (несущей дефектную митохондриальную ДНК) в другую (здоровую) можно эффективно заменить цитоплазму клетки и вместе с ней митохондрии (и их ДНК)
. Полученная таким образом яйцеклетка может рассматриваться как имеющая двух матерей. Эмбрион образующийся после оплодотворения такой яйцеклетки будет иметь здоровую митохондриальную ДНК
. Однако насколько оправданы подобные манипуляции с клетками человека с точки зрения
биоэтики
пока не ясно
.
Подробнее о новейших достижениях техники клонирования и получении тотипотентных клеток с помощью SCNT см.:
Перепрограммирование в иТК без помощи SCNT
До недавнего времени получить тотипотентные клетки удавалось лишь с помощью SCNT. Однако появились работы где было продемонстрировано получение иТК с помощью перепрограммирования факторами Яманаки in vivo
, а также in vitro с помощью таких эпигенетических факторов ооцита как зародышевая изоформа
гистонов
. Перевести
эмбриональные стволовые клетки
в состояние тотипотентности характерное для ранних эмбрионов 2-клеточной стадии можно также путём подавления активности
CAF-1
необходимой для сборки хроматина
.
От превращения ЭСК и ИПСК в тотипотентные клетки, способные дать начало таким внеэмбриональным тканям как
плацента
и
желточный мешок
, плюрипотентные стволовые клетки, по-видимому, так же удерживает
. Ингибирование её синтеза приводит к активации экспрессии эндогенного
ретровируса
MuERV-L и расширяет потенциал плюрипотентных стволовых клеток до способностей клеток стадии двух бластомеров
.
Разработан химический коктейль для получения из клеток мочи человека так называемых плюрипотентных стволовых клеток с расширенными возможностями по дифференцировке — они могут дать начало как клеткам эмбриона, так и внеэмбриональным тканям.
Такие супертотипотентные клетки (иСТК) могут быть использованы для получения
химер
с целью
выращивания органов в организме животных
В частности разработана комбинация из трёх малых молекул: производного
ретинола
тератогена
TTNPB
(4-[(1E)-2-(5,5,8,8-тетраметил-5,6,7,8-тетрагидро-2-нафталинил)-1-пропен-1-ил]бензойная кислота), высокоселективного ингибитора
1-Азакенпауллона
(1-Azakenpaullone) и
митогена
WS6
(N-[6-[4-[2-[4-[(4-метилпиперазин-1-ил)метил]-3-(трифторметил)анилино]-2-оксоэтил]фенокси]пиримидин-4-ил]циклопропанкарбоксамид), которая позволила проводить индукцию и длительное поддержание культуры тотипотентных стволовых клеток из плюрипотентных стволовых клеток мыши
.
Получение репродуктивных клеток из ИПСК
Используя среды, содержащие
ретиноевую кислоту
и
фолликулярную жидкость
свиньи, можно получить in vitro, дифференцировкой из ИПСК, клетки ранних стадий
гаметогенеза
подобные репродуктивным клеткам, из которых образуются сперма и ооциты
. Примечательно, что для образования примордиальных половых клеток человека требуется активность двух ключевых регуляторов: гена SOX17 направляющего дифференцировку в сторону образования предшественников половых клеток и
подавляющего энтодермальные и другие соматические гены во время этой специализации
.
В статье китайских учёных с первым автором Чжоу (Zhou) описана технология дифференциации мышиных эмбриональных стволовых клеток которые претерпевают мейоз in vitro превращаясь в гаплоидные сперматиды, способные к оплодотворению, о чём свидетельствовало получение с их помощью жизнеспособного и фертильного потомства
.
Подробный обзор методов искусственного получения мужских половых клеток можно найти в статье Hou с соавторами
и Irie, Kim, Surani
, а также.
Разработана технология позволяющая получать зрелые ооциты in vitro из эмбриональных стволовых клеток, а также из индуцированных плюрипотентных стволовых клеток, полученных из взрослых фибробластов взятых с кончика мышиного хвоста. Более того оплодотворив такие яйцеклетки in vitro и подсадив их в матку мыши удалось с выходом в 1 % получить жизнеспособное потомство
.
Эта технология послужит платформой для выяснения молекулярных механизмов, лежащих в основе тотипотентности и для разработки методов производства ооцитов из других (в том числе редких) видов млекопитающих в лабораторных условиях.
ИПСК как результат радикального омоложения
Предыстория открытия
Впервые ИПСК были получены в виде перевиваемой
тератокарциномы
, индуцированной трансплантатом, взятым из мышиных эмбрионов
. Было доказано, что тератокарциномы образуются из соматических клеток
. Тот факт, что из клеток тератокарциномы можно получить нормальную мышь доказывал их плюрипотентность
. Оказалось, что клетки тератокарциномы, выделяя в культуральную среду различные факторы, способны поддерживать культуру плюрипотентных стволовых клеток эмбриона в недифференцированном состоянии
. Таким образом, ещё в 1980-е годы стало ясно
, что трансплантация плюрипотентных или эмбриональных стволовых клеток во взрослый организм млекопитающих обычно приводит к образованию
тератомы
, которая затем может превратиться в злокачественную опухоль — тератокарциному
. Если, однако, поместить клетки тератокарциномы в ранний зародыш млекопитающего (на стадии бластоцисты), то они включаются в состав
клеточной массы бластоцисты
и из такого химерного (то есть состоящего из клеток от разных организмов) эмбриона нередко развивается нормальное химерное животное. Почти во всех органах и тканях которого часть дифференцированных клеток происходит из клеток тератокарциномы, которые совместно с клетками нормального происхождения участвуют в построении здорового организма
. Это свидетельствовало о том, что причиной образования тератомы является диссонанс в стадии развития донорных клеток и окружающих их клеток реципиента (так называемой
ниши
). Уже тогда, используя ретровирусные векторы, удалось ввести инородные гены в мышиные химеры, полученные с помощью клеток тератокарциномы
.
Открытие роли репрограммирующих факторов
В августе 2006 года японские исследователи сумели превратить клетки мышиной кожи (фибробласты) в индуцированные плюрипотентные стволовые клетки — ИПСК (induced pluripotent stem cells — iPSC), используя для модификации клетки всего четыре репрограммирующих фактора:
Oct4
,
,
и
, доставленных в ядро ретровирусами
. Этим они доказали, что гиперэкспрессия небольшого количества факторов иногда может подтолкнуть клетки к переходу в новое стабильное состояние, связанному с изменениями активности тысяч генов. По своим свойствам ИПСК оказались очень похожи на эмбриональные стволовые клетки (ЭСК)
. Так, сравнение протеома и фосфопротеома ЭСК и ИПСК, проведённое на 4-х линиях человеческих эмбриональных стволовых клеток и 4-х линиях индуцированных плюрипотентных стволовых клеток, показало, что большинство идентифицированных белков и участков фосфорилирования в белках всех линий совпадают. Хотя были и небольшие, но статистически воспроизводимые различия, свидетельствующие об определённом функциональном различии
. Не было отмечено и особых изменений в последовательности ДНК, особенно если ИПСК были получены с помощью неинтегрирующихся в геном плазмид
.
Позднее, с развитием технологии перепрограммирования, лучшим доказательством идентичности ИПСК и ЭСК стала возможность получения взрослой мыши полностью из некоторых линий ИПСК
. Несмотря на то, что рядом исследований была доказана идентичность ЭСК и ИПСК
, получаемые клоны сильно отличаются друг от друга и не для всех из них можно доказать идентичность с ЭСК
, далеко не все клоны способны дать жизнь химерным мышам или подвергнуться эффективной дифференциации в те или иные соматические клетки. Одной из причин таких различий является разница между составом транскрипционных факторов при репрограммировании в ИПСК и набором факторов в материнском ооците. К числу таких «упущенных» факторов относится, в частности способствующий процессу репрограммирования особый, характерный для ооцитов линкерный (связующий, компонующий нуклеосомы) гистон H1foo
. Замена одного из факторов Яманаки, а именно c-Myc на H1foo, значительно повысили количество и качество получаемых клонов ИПСК — они стали более однородными по свойствам, из них чаще стали получаться мыши-химеры
.
Oct4 положительно регулирует гены, связанные с плюрипотентностью и самообновлением, а также подавляет гены, способствующие дифференцировке
. Избыточная экспрессия Oct4 во время перепрограммирования ухудшает качество ИПСК — по сравнению с OSKM (Oct4, Sox2, Klf4 и c-Myc), перепрограммирование
SKM
(Sox2, Klf4 и c-Myc) генерирует ИПСК с высоким потенциалом развития (почти в 20 раз выше, чем у OSKM), что доказано по их способности генерировать мышей методом
из эмбрионов полностью состоящих из ИПСК
. В то же время перепрограммирование SKM видоспецифично, оно может быть достигнуто в клетках мыши, но не в клетках человека
Технология получения ИПСК является способом радикального омоложения
Важным преимуществом ИПСК перед ЭСК является то, что они могут быть получены из клеток взрослого организма, а не из эмбриона. Поэтому стало возможным получать ИПСК от взрослых и даже пожилых пациентов
. Перепрограммирование соматических клеток в ИПСК приводит к их омоложению о чём свидетельствуют данные исследования теломеров— концевых участков хромосом состоящих из коротких следующих друг за другом повторов эволюционно консервативной последовательности ДНК. Выяснилось, что перепрограммирование приводит к удлинению теломеров и их нормальному укорочению по мере дифференцировки ИПСК обратно в фибробласты
. Таким образом, при индуцированной плюрипотенции восстанавливается эмбриональная длина теломеров
, а значит, увеличивается потенциальное число делений клетки
, ограниченное так называемым
лимитом Хайфлика
(Hayflick limit). Более того омолаживаются и митохондрии клетки при этом восстанавливается характерный для молодых клеток уровень дыхания
Поэтому технология получения ИПСК является способом радикального омоложения
.
Тератомы как препятствие на пути внедрения технологии в клинику
Удаление опасных плюрипотентных клеток с помощью индуцируемого гена клеточного самоубийства
. iCasp9 был сконструирован из
такролимус
(FK506) связывающего домена димеризации FKBP12 с мутацией F36V, повышающей его способность к связыванию индуцирующей малой молекулы; линкерной последовательности аминокислот (Ser-Gly-Gly-Gly-Ser), которая обеспечивает гибкость конструкции, и человеческого гена Casp9, в котором удалён домен связывания каспазы ΔCasp9. Благодаря такой конструкции димеризация генноинженерного белка iCasp9, необходимая для активации ферментативной активности, происходит только при наличии синтетической малой молекулы (например, AP1903), вызывающей димеризацию с помощью домена FKBP12.
Из-за диссонанса в стадии развития омоложённых клеток и окружающих их старых клеток реципиента, инъекция пациенту его же собственных ИПСК, обычно приводит к иммунной реакции
, что может быть использовано в медицинских целях
, или образованию опухолей типа тератомы
.
Одной из причин иммуногенности аутологичных ИПСК и ЭСК считается группа из 9 генов (Hormad1, Zg16, Cyp3a11, Lce1f, Spt1, Lce3a,Chi3L4, Olr1, Retn), синтез которых повышен в тератомах, полученных из этих клеток
Очевидно, некоторые клетки, дифференцированные из ИПСК и ЭСК, продолжают синтезировать эмбриональные изоформы белков
и неадекватно интерпретируют сигналы окружающих их клеток реципиента.
Образование тератомы из плюрипотентных стволовых клеток может быть вызвано низкой активностью фермента
PTEN
, способствующей выживанию, в процессе дифференцировки, небольшой популяции (не превышающей 0,1-5 % от общей численности клеток) высоко онкогенных клеток карциномы, инициирующих тератомы. Выживание этих инициирующих тератомы клеток связано с недостаточной репрессией
Nanog
, а также с повышением метаболизма глюкозы и холестерина.
Эти, инициирующие образование тератом, клетки характеризуются также более низким соотношением
p53
/
p21
по сравнению с неонкогенными клетками.
Недавно методом отбора удалось найти небольшие молекулы (цитотоксические селективные ингибиторы плюрипотентных стволовых клеток человека), которые предотвращают образование тератомы у мышей после трансплантации им плюрипотентных стволовых клеток человека. Самое мощное и селективное из этих соединений — PluriSIn #1, вызывало ингибирование стеароил-КоА десатуразы (ключевого фермента в биосинтезе
олеиновой кислоты
), что в конечном итоге приводило к апоптозу плюрипотентных стволовых клеток. С помощью этой молекулы удаётся выборочно удалить из культуры недифференцированные клетки.
. Ещё одной молекулой избирательно удаляющей недифференцированные клетки является STF-31,
являющийся ингибитором
GLUT1
.
Эффективной стратегией избирательного устранения плюрипотентных клеток, которые способны дать начало тератоме является ингибирование характерных для этих клеток антиапоптотических факторов, таких как
сурвивин
или
. Обработкой малыми молекулами, которые могут ингибировать эти антиапоптотические факторы, можно добиться селективного удаления подобных клеток вызвав их
апоптоз
. В частности, одной обработки смешанной популяции химическими ингибиторами сурвивина (такими как, например,
кверцетин
или YM155) достаточно чтобы вызвать избирательную и полную гибель недифференцированных клеток, вызванную накоплением р53 в митохондриях. Этого, по мнению авторов исследования, достаточно, чтобы предотвратить образование тератомы после трансплантации клеток полученных из ИПСК
.
.
Тем не менее, маловероятно, что какая либо, пусть даже самая изощрённая, предварительная очистка
, способна обезопасить подсадку ИПСК или ЭСК, так как при избирательном удалении плюрипотентных клеток, они вновь довольно быстро возникают путём превращения дифференцированных клеток обратно в стволовые (к обратному переходу может в частности подтолкнуть гипоксия
), что приводит к образованию опухоли
. Это может быть связано с нарушением регуляции осуществляемой микро РНК let-7 по отношению к её мишени — белку Nr6a1 (известному также как ядерный фактор зародышевых клеток — GCNF), являющемуся эмбриональным репрессором транскрипции генов плюрипотентности, который необходим для правильной дифференцировки индуцированных плюрипотентных клеток.
Обнаружена также малая молекула названная Дисплюригеном (Displurigen), которая воздействуя на
белок теплового шока
(Heat shock 70 kDa protein 8), необходимый для связывания
с ДНК, способна вывести клетку из состояния плюрипотентности
. Ещё один способ предотвратить образование тератомы — это вызвать в пересаживаемой клетке ИПСК гиперэкспрессию гена
Использование ИПСК для клеточной терапии пока ограничено.
Тем не менее, они могут быть использованы для целого ряда иных целей — включая моделирование болезней, скрининг (селективный отбор) лекарств, проверку токсичности различных препаратов
.
Важными факторами для получения высококачественных ИПСК являются определённые небольшие молекулы, способствующие сохранению геномной целостности, образующихся при перепрограммировании ИПСК, путём ингибирования двухцепочечных разрывов ДНК и активации гена Zscan4, содействующего процессам репарации ДНК
. Перепрограммирование вызывает
репликативный
стресс
, который можно снизить повысив уровень чекпоинт киназы 1 (
), благодаря чему повышается качество и эффективность образования ИПСК. Кроме того добавление во время перепрограммирования
нуклеозидов
позволяет снизить повреждения ДНК и число геномных перестроек в получаемых ИПСК
Ткани, выращенные из ИПСК, помещённых в «химерные» эмбрионы на ранних стадиях развития мыши, практически не вызывают иммунного ответа (после того, как эмбрионы выросли во взрослых мышей) и пригодны для
аутологичной трансплантации
,
причём даже в том случае когда ИПСК получены от очень старых животных
.
Перепрограммирование с омоложением
in vivo
В ранних работах полное перепрограммирование взрослых клеток в тканях у мышей
in vivo
путём временной активации факторов Oct4, Sox2, Klf4 и с-Мус, приводило к образованию в различных органах множества тератом
. Более того, частичное перепрограммирование клеток в ИПСК
in vivo
показало, что неполное перепрограммирование приводит к эпигенетическим изменениям (нарушению репрессии
Поликомб
целей и изменению
метилирования ДНК
) в клетках, которые ведут к развитию рака
Способы омолаживающего
без достижения плюрипотентности
Изменив продолжительность и дозировку удалось провести без последующего канцерогенеза циклическое частичное перепрограммирование
in vivo
путём экспрессии факторов Яманака в течение короткого периода времени (с их экспрессией в течение 2 дней и интервалом без экспрессии в течение 5 дней). Такими, циклически повторяемыми активациями факторов Яманака, удалось частично омолодить, и, таким образом, продлить продолжительность жизни прогероидных мышей.
Используя мышиную модель, которая обеспечивает индуцируемую экспрессию 4F - четырёх факторов Яманаки (Oct-3/4, Sox2, Klf4 и c-Myc) удалось путём их временной экспрессией
in vivo
индуцировать частичное перепрограммирование взрослых гепатоцитов в состояние предшественников и увеличить пролиферацию клеток, что по мнению авторов статьи может противодействовать печёночной недостаточности
Частичное эпигенетическое перепрограммирование 4F у старых мышей позволило обратить вспять некоторые маркеры старения печени, но только в сочетании с истощением богатых маркером старения клеток p16
High
, которые ограничивали пластичность клеток
.
В опытах
in vitro
, при использовании несколько более длительных периодов перепрограммирования (чтобы достичь более существенного омоложения) клетки теряли свою клеточную идентичность
, но затем восстанавливали свою первоначальную соматическую идентичность, при удалении факторов репрограммирования
.
Длительное частичное перепрограммирование
in vivo
приводит к омолаживающим эффектам в различных тканях, таких как почки и кожа, а также в целом на уровне организма. Многократность лечения определяла степень положительного эффекта. Омолаживающие эффекты были связаны со снижением возраста по данным
эпигенетических часов
, а также по данным метаболических и транскриптомных изменений, включая снижение экспрессии генов, участвующих в путях
воспаления
,
клеточного старения
и реакции на
стресс
Эффективность методов частичного перепрограммирования как in vitro, так и in vivo пока очень низкая, поскольку клетки в процессе частичного перепрограммирования усиливают экспрессию NK-активирующих
лигандов
, таких как
и
ICAM-1
, в результате чего
NK-клетки
распознают и убивают частично перепрограммированные клетки. Поэтому повысить эффективность частичного перепрограммирования
in vivo
помогает истощение пула NK-клеток
Алгоритм для предсказания набора транскрипционных факторов необходимых для преобразования клеток
Определение уникального набора транскрипционных факторов, которые необходимы для репрограммирования клеток представляет собой длительный и дорогостоящий процесс. Международная группа исследователей разработала алгоритм, называемый Магрифи (Mogrify), который помогает предсказать оптимальный набор клеточных факторов, необходимых для преобразования одного типа клеток человека в другой
. Появился также алгоритм, который предсказывает не только транскрипционные факторы необходимые для перепрограммирования, но также и идеальный выбор времени для применения этих факторов.
Поскольку выяснилось что
нуклеосомы
с меткой на
гистоне
H3 называемой
обычно сидят на последовательностях ДНК предшествующих генам, которые важны для определения судьбы клетки, предопределения её типа, стало возможным с помощью алгоритма эпиМагрифи (EpiMOGRIFY) находить подобные гены чтобы влиять на дифференцировку культивируемых клеток
Спрогнозировать результаты целенаправленных вмешательств и помочь разработке процессов перепрограммирования может программа cSTAR (cell state transition assessment and regulation) классифицирующая состояния клеток на основании данных
протеогеномики
Атлас транскрипционных факторов для направленной дифференцировки
Факторы транскрипции (TFs) регулируют генные программы, тем самым контролируя различные клеточные процессы и состояния клеток.
Создан Атлас транскрипционных факторов для направленной дифференцировки и библиотека
MORF
(multiplexed overexpression of regulatory factors) мультиплексной гиперэкспрессии регуляторных факторов, которые помогут идентифицировать комбинации факторов транскрипции, которые управляют определенными фенотипами.
Стратегии получения ИПСК для клинических испытаний
Разработаны критерии качества и стратегия производства ИПСК для клинических испытаний, так называемая cGMP (
англ.
current Good Manufacturing Practice
)
.
Стратегия получения универсальных ИПСК
Чтобы сделать доступными технологии регенеративной медицине на основе ИПСК большему числу пациентов, необходимо создавать универсальные ИПСК, которые можно трансплантировать независимо от
гаплотипов
HLA
. Текущая стратегия создания универсальных ИПСК преследует две основные цели: удаление экспрессии HLA и предотвращение атак со стороны
NK-клеток
, которые вызваны
делецией
HLA. Сообщалось, что делеция генов
B2M
и
с использованием системы
CRISPR/Cas9
подавляет экспрессию HLA класса I и класса II соответственно. Для предотвращения атак NK-клеток использовалась
трансдукция
лигандов
, ингибирующих NK-клетки, таких как
и
CD47
.
при этом оставляют без изменения, поскольку 12 часто встречающихся аллелей HLA-C достаточно для охвата 95 % населения мира.
Система индуцируемого апоптоза для безопасности
Чтобы обезопасить применение ИПСК в клинике, было предложено одновременно с перепрограммированием клеток пациента в ИПСК, вводить в эти клетки индуцируемый малой молекулой ген
каспазы-9
(IC9) для запуска каскадов
апоптоза
для самоубийства клеток образованных из этих ИПСК
. Такой «предохранитель» позволит избавляться от омоложённых клеток после того как они выполнили свою терапевтическую функцию или в случае образования опухоли из этих клеток
.
Устойчивость к онкогенезу у ИПСК Голого землекопа
У
голых землекопов
уровень заболеваемости раком крайне низок по сравнению с другими млекопитающими. Обнаружено, что у ИПСК этого животного ослаблена способность к образованию тератом при трансплантации, что может быть связано
:
с видоспецифической активацией
супрессора опухоли
(
англ.
alternative reading frame
), который является продуктом альтернативной рамки считывания гена
(другой продукт этого гена — маркер старения белок
), а также
с мутацией, приводящей к разрушению онкогена ERAS, являющегося аналогом
Ras
и отвечающего за онкогенность ЭСК.
Более того, удалось найти сигнальный путь ASIS (
англ.
ARF suppression-induced senescence
), с помощью которого вероятно удастся защитить ИПСК от возникновения из них опухолей.
Эффективность перепрограммирования в ИПСК
До настоящего времени недостаточно понятно, почему эффективность перепрограммирования с помощью
факторов транскрипции
значительно ниже, чем при пересадке ядра в ооцит. Показано, что большинство фибробластов кожи взрослого человека начинают процесс перепрограммирования сразу после обработки трансгенами Яманаки (
Oct4
,
,
, и
).
Помимо этих факторов в «коктейль» для репрограммирования можно также добавлять фактор CECR2 необходимый для преодоления эпигенетических барьеров во время репрограммирования.
Тем не менее, только небольшая часть (~ 1 %) из этих «новоиспечённых» ИПСК образуют впоследствии колонии ИПСК
. Причиной, понижающего эффективность перепрограммирования, возврата большинства клеток к состоянию дифференцировки может быть:
недостаточная деятельность активируемой цитидиндезаминазы (AID) из-за чего клетки не могут стабилизироваться и долго поддерживать состояние плюрипотенции
.
недостаточная активность гена
кодирующего один из белков
когезина
(необходимого для образования внутрихромосомной петли сближающей промоутер гена с последующим энхансером, что необходимо для активации эндогенных генов плюрипотентности), делает невозможным достижение плюрипотентности
важную роль на поздних этапах перепрограммирования играют и ферментативные модификации
гистонов
. В частности
-зависимая модификация
является барьером препятствующим репрограммированию клеток млекопитающих
. Показано, что необходимым условием эффективного перепрограммирования является подавление переносчика гистонов
CAF-1
и белкового комплекса ремоделирования нуклеосом и деацетилирования (nucleosome remodeling and deacetylation —
. Избыточная экспрессия субъединицы NuRD, называемой
, ингибирует индукцию ИПСК. Причиной этого является деацетилирование комплексом NuRD
лизина
27 в молекуле
гистона
Н3К27ac, что позволяет
Поликомб Репрессорному комплексу 2 (PRC2)
осуществить триметилирование лизина 27 в гистоне H3, приводящее, в конечном счёте, к ингибированию ряда генов-маркеров плюрипотентности
, в том числе генов Oct4 и Nanog. Ингибирование Mbd3, с другой стороны, повышает эффективность перепрограммирования и способствует образованию плюрипотентных стволовых клеток, которые способны генерировать жизнеспособных химерных мышей, даже в случае отсутствия с-Мус или Sox2
. Очевидно, Mbd3/NuRD исполняет роль эпигенетического регулятора, который ограничивает экспрессию ключевых генов плюрипотентности. Поэтому подавление Mbd3/NuRD (например, с помощью бутирата, вальпроевой кислоты, субероиланилидгидроксамовой кислоты или трихостатина А) может стать мощным средством для повышения эффективности и точности перепрограммирования. Действительно, подавив Mbd3 удалось впервые осуществить детерминированное и синхронизированное перепрограммирование клеток кожи мыши и человека в ИПСК в течение всего семи дней и с невиданной ранее эффективностью — около 100 %
Найден фактор BRD3 (
), который опознаёт «коды»
ацетилированных гистонов
в хромосоме, а также активирует большой набор
митотических
генов, повышая таким образом митотическую активность клетки. Этот фактор позволил более чем в 20 раз повысить эффективность выхода ИПСК, сократить длительность перепрограммирования до нескольких дней и повысить качество перепрограммирования
. Как отмечено выше повысить эффективность репрограммирования позволяет также замена с-Мус на H1foo
. В случае когда требуется репрограммировать клетки пожилых пациентов повысить эффективность позволяет ингибирование H3K79 гистон метилтрансферазы называемой
(Disruptor of telomeric silencing 1-like)
Сконструирован супер-SOX2-17 транскрипционный фактор состоящий из факторов
и
, который, будучи внесён в состав коктейлей усиливает перепрограммирование в сотни раз и позволяет перепрограммировать в ИПСК клетки пожилых людей, не поддававшиеся перепрограммированию
.
Предлагается также для повышения генетической стабильности ИПСК помимо факторов Яманаки во время перепрограммирования клеток использовать также трансфекцию
циклином D1
с целью повышения процессов
репарации ДНК
и уменьшения клеточного стресса
.
Элитные клетки
В первичных после биопсии культурах клеток при перепрограммировании лишь очень немногие клетки способны превратиться в ИПСК, и тех из них, которые такой способностью обладают, называют «элитными» клетками. Учёные нашли способ получения таких элитных клеток из соматических с помощью фактора
. В первичной культуре мышиных В-клеток непродолжительная экспрессия C/EBPα с последующим перепрограммированием факторами Яманаки позволила добиться 100-кратного увеличения эффективности перепрограммирования в плюрипотентные клетки, причём с участием 95 % клеточной популяции
. Такие искусственно созданные элитные клетки очень похожи на белые кровяные прогениторные клетки-предшественники
костного мозга
, известные как
миелобласты
. К сожалению, сверхэкспрессия C/EBPα может способствовать развитию острого миелоидного лейкоза и острого лимфобластного лейкоза делая клетки устойчивыми к лечению.
Замечено, что успешно репрограммируются те из фибробластов, которые имеют небольшой размер клетки и имеют более высокую способность к пролиферации. Их можно выявить и выделить по содержанию транскрипционного фактора
.
Роль витаминов в репрограммировании
Витамин С
Известно, что
аскорбиновая кислота (витамин С)
усиливает процесс перепрограммирования
с помощью нескольких механизмов, в первую очередь из-за ее кофакторной роли в Fe (II) и 2-оксоглутарат-зависимых диоксигеназах, включая
и
деметилазы гистонов
.
Витамин B
12
Обнаружено, что
витамин B
12
является ограничивающим фактором клеточного перепрограммирования как
in vitro
, так и
in vivo
.
Дифференцировка ИПСК в зрелые клетки
В тератоме
Тот факт, что ИПСК человека способны к образованию тератом не только в теле человека, но и в организме некоторых животных, в частности в организме мыши или свиньи, позволил разработать метод дифференцировки ИПСК в условиях in vivo. Для этого ИПСК вводят, вместе с клетками индуцирующими направленную дифференцировку,
генмодифицированной
свинье или мыши, у которой подавлена активация иммунной системы на клетки человека, а затем, вырезав образовавшуюся тератому, выделяют из неё необходимые дифференцированные клетки человека,
используя
моноклональные антитела
к тканеспецифичным маркерам на поверхности полученных клеток. Этот метод был успешно использован для получения функциональных
мышечных
, а также
миелоидных, лимфоидных и эритроидных
клеток человека пригодных для трансплантации (пока только мышам). Таким образом, доказана возможность производства in vivo из клеток пациента необходимых ему дифференцированных клеток для трансплантации, изготовления антител или скрининга лекарственных средств
. Используя перевиваемую генмодифицированную тератому с гиперэкспрессией факторов Gfi1b, c-Fos и Gata2 можно неоднократно трансплантировать мышкам тератому и на протяжении длительного времени стабильно получать полностью функциональные мышиные гематопоэтические стволовые клетки
Используя лектин rBC2LCN избирательно связывающий ИПСК
, или же MitoBloCK-6
и /или PluriSIn #1 можно очистить полученные прогениторные клетки от плюрипотентных клеток образующих тератому. Тот факт, что дифференцировка проходит в условиях тератомы позволяет надеяться, что полученные клетки достаточно устойчивы к стимулам способным запустить их обратный переход к дедифференцированному (плюрипотентному) состоянию, а значит безопасны.
Беспокойство, однако, вызывает тот факт, что «воспитанные» в тератоме у животных человеческие клетки за время своего «воспитания», по всей вероятности, поглощают значительное количество
экзосом
произведённых окружающими клетками организма носителя тератомы, а значит, попав в организм человека, могут повести себя неадекватно.
Методика основанная на обнаружении
ген-репортёр
-
GFP
-положительных клеток в тератоме, полученной из ИПСК, позволит идентифицировать и вырастить культуры ткани, используя индуцированные взрослые стволовые клетки различных типов, выделение которых ранее было затруднительно
.
В организме животных-биоинкубаторов
Весьма перспективной средой для первоначальной дифференцировки ИПСК in vivo могут оказаться куриные эмбрионы
. Есть доказательства того, что микросреда этих эмбрионов оказывает антионкогенное действие на человеческие клетки и намного лучше чем условия in vitro
Разработана технология «дозревания», полученных из ИПСК в условиях
in vitro
человеческих прогениторных клеток кардиомиоцитов, путём
ксенотрансплантации
их в организм новорождённых крыс, используемых в качестве
in vivo
биоинкубатора. Такое «дозревание» занимает ~6 недель
См. также:
Robert Lanza
, Michael West (2013)
. Patent US 20130058900 A1
Получение клеток хрусталика и сетчатки глаза из ИПСК
В ближайшее время предполагается приступить к клиническим испытаниям, призванным продемонстрировать безопасность использования ИПСК для клеточной терапии людей с
катарактой
а также с возрастной дегенерацией жёлтого пятна — заболевания, которое повреждая
сетчатку
, может привести к слепоте
. Описаны методы получения из ИПСК клеток
хрусталика
и сетчатки
и способы их использования для клеточной терапии
, которая по крайней мере на 6 недель улучшала зрение у подопытных животных
.
Получение из ИПСК лёгочных эпителиальных клеток
Хронические заболевания лёгких, такие как
идиопатический фиброзирующий альвеолит
,
Силикоз
,
хроническая обструктивная болезнь лёгких
и
бронхиальная астма
входят в число основных причин инвалидности и смертности. Поэтому исследователи ищут пути эффективной клеточной терапии и тканевой инженерии лёгких, которые бы позволили бороться с этими заболеваниями
.
Были разработаны способы получения различных типов лёгочных клеток из ИПСК, которые могут быть взяты за основу для получения терапевтических клеток из материала, полученного от пациента.
Получение нервных стволовых клеток человека из ИПСК
Юань и коллеги сообщили, что нервные стволовые клетки человека, индуцированные из ИПСК с помощью ретиноевой кислоты в бессывороточной среде имеют стабильный нейронный фенотип. После трансплантации крысам со смоделированным ишемическим инсультом, эти клетки не только выжили, но и мигрировали в зону ишемии мозга, где дифференцировались в зрелые нервные клетки, что оказало благотворное влияние на функциональное восстановление утраченных от повреждения в результате инсульта неврологических функций
.
Получение стволовых клеток почки из ИПСК
Разработана система для быстрого (за 3 дня) и эффективного (70 %-80 % популяции) превращения ИПСК в клоны характерные для клеток почки с помощью ингибитора CHIR99021 и некоторых ростовых факторов
. Более того, удалось излечивать в опытах на мышах острые поражения почек, используя стволовые клетки почки, полученные из ИПСК
.
Получение остеобластов из ИПСК
Известно что
аденозин
и его рецепторы, в частности
играют важную роль в регенерации
костных
переломов
. Простое добавление в культуральную среду аденозина позволило превратить человеческие ИПСК в остеобласты. При трансплантации этих остеобластов мышке, с использованием макропористой синтетической матрицы, остеобласты полученные из ИПСК, участвовали в регенерации повреждений кости образуя новые ткани и стимулируя кальцификацию. При этом не наблюдалось образования тератом, что очевидно свидетельствует о 100 % дифференцировке клеток ИПСК в остеобласты
.
Наивные плюрипотентные стволовые клетки (нПСК)
Человеческие плюрипотентные стволовые клетки, независимо от того, получены ли они из бластоцисты или являются результатом перепрограммирования соматических клеток, существенно отличаются от классических мышиных эмбриональных стволовых клеток и по мнению ряда исследователей представляют более позднюю стадию развития
эпибласта
. Удалось получить нПСК у которых утеряна эпигенетическая «память»
метилирования ДНК
как
гаметы
(
ооцита
), так и человеческой
бластоцисты
. Такие клетки в отличие от ИПСК не имеют антиген SSEA4 (Stage Specific Embryonic Antigen 4)
.
Перевести ЭСК и ИПСК человека в наивное состояние позволяет гиперэкспрессия фактора YAP (Yes-associated protein). Гиперэкспрессию YAP с получением наивного состояния можно также имитировать путём добавления к культуральной среде (
лизофосфатидной кислоты
(LPA), являющейся активатором YAP
.
Репрограммирование человеческих ЭСК и ИПСК с помощью рекомбинантного, усечённого человеческого NME7 (найденного в семенниках фактора, содержащего два
домена
нуклеозид
дифосфат киназы (
) и способного связываться с расщеплённой формой трансмембранного рецептора
MUC1
, называемой MUC1*
) позволило получить стабильно наивные клетки, которые более пригодны для широкомасштабного клонирования и имеют расширенный потенциал дифференциации
. На основе таких клеток можно создать «фабрики клеток» для промышленного производства продукции необходимой для нужд
клеточной терапии
.
Регион-селективные плюрипотентные стволовые клетки (рсПСК)
Ву и его коллеги обнаружили, что комбинация свободной от сыворотки среды,
фактора роста фибробластов 2 (FGF2)
и ингибитора
сигнальных путей Wnt
позволяет получить в результате устойчивую линию рсПСК (регион-селективных плюрипотентных стволовых клеток,
по англ
rsPSCs) клеток человека.
По
транскриптому
эти клетки напоминали таковые из задних клеток раннего
эмбриона
мыши. Трансплантация этих клеток в 7,5-дневные эмбрионы мыши привела к их эффективному включению в задний, но не в другие части эмбриона. После 36 часов культивирования этих
химерных
эмбрионов, клетки рсПСК проявили способность к пролиферации и способность к дифференцировке в ткани трёх
зародышевых листков
. Хотя исследователи остановили дальнейшую дифференцировку этих клеток, предполагается, что каждый из образованных этими клетками зародышевых листков способен дать начало определённым тканям и органам
.
В отличие от других человеческих стволовых клеток, которые, как правило, не удаётся интегрировать в эмбрион мыши, человеческие rsPSCs способны к такой интеграции и к развитию в ранние стадии тканей человека
.
Клетки F класса
Клетки F класса в отличие от ИПСК не способны включаться в ткани организма и участвовать в построении
химерного
организма. Тем не менее они удовлетворяют другому тесту на плюрипотентность — способны образовывать
тератомы
. По сравнению с обычными стволовыми клетками подобными эмбриональным и ИПСК,клетки F-типа растут в лаборатории быстрее и их выращивать проще и дешевле — их можно просто поместить в большой сосуд с питательной средой и вырастить за несколько дней или часов, а не за несколько недель как обычные ИПСК
.
Индуцированные прогениторные стволовые клетки
Методы прямой трансдифференцировки
В связи с тем, что использование ИПСК для клеточной терапии сопряжено с большим риском опухолей и рака, необходима разработка методов получения более безопасных клеточных линий, пригодных для применения в клинике. Альтернативой методам ИПСК стала техника так называемого «прямого репрограммирования», то есть индуцируемой определёнными факторами прямой трансдифференцировки, без предварительного прохождения клеток через стадии плюрипотентного состояния
. Основу для такого подхода заложили исследования Тейлор и Джонса (Taylor and Jones), показавших, что воздействие
5-азацитидина
— реактива, вызывающего деметилирование ДНК — на бессмертную линию клеток мышиных эмбриональных фибробластов способно вызвать образование миогенных, хондрогенных и адипогенных клонов
и Вейнтрауба с соавторами, обнаруживших, что для репрограммирования достаточно активации всего одного гена, позднее названного MyoD1
. По сравнению с ИПСК, для получения которых требуются не менее двух недель, образование индуцированных прогениторных клеток происходит сравнительно быстро — иногда за несколько дней. Эффективность перепрограммирования также обычно во много раз выше. Для этого перепрограммирования не всегда требуется деление клетки
. Но главное, это то, что получаемые в результате перепрограммирования мультипотентные соматические стволовые клетки более пригодны для клеточной терапии, так как не образуют тератомы
. Это очевидно связано с тем что при прямой трансдифференцировке в получаемых клетках сохраняются признаки старения исходных клеток.
См. также обзоры
Трансдифференцировка с помощью 5-азацитидина и тромбоцитарного фактора роста
Разработан метод получения, так называемых, индуцированных мультипотентных стволовых клеток (ИМПСК) путём непродолжительной обработки
постнатальных стволовых клеток
костного мозга и жировых клеток комбинацией фактора роста (
тромбоцитарный фактор роста
— АВ (PDGF-AB)) и
5-азацитидина
. Авторы этого исследования утверждают, что: «В отличие от первичных мезенхимальных стволовых клеток, которые хотя и используются в клинической практике для содействия восстановлению тканей, но не способны сами включаться в эту ткань, ИМПСК способны к непосредственному участию процессах регенерации тканей и при этом не образуют опухолей», в связи с чем «могут быть использованы для регенерации различных тканей»
Трансдифференцировка зрелых клеток всего одним фактором транскрипции
Особенностью
нематоды
Caenorhabditis elegans
является настолько жёсткая программа развития, что соматическая клетка, находящаяся в определённом участке организма, как правило, имеет одинаковую родословную у всех особей.
При этом зрелые клетки, в отличие от ранних эмбриональных клеток, обычно очень устойчивы к изменению их фенотипа. Тем не менее, обнаружено, что как у интактных личинок, так и у неповреждённых взрослых нематод краткосрочный синтез всего одного фактора транскрипции, а именно фактора ELT-7
может превратить фенотип полностью дифференцированной, высокоспециализированной не-энтодермальной клетки фаринкса (глотки) в фенотип полностью дифференцированной энтодермальной клетки кишечника. Это превращение происходит «в одну стадию» — путём прямой трансдифференцировки, без каких-либо промежуточных стадий дедифференцировки
.
Трансдифференцировка с помощью CRISPR-опосредованного активатора
Фенотип клетки можно изменять с помощью
. Например, путём активации определённых эндогенных генов, используя для этого
CRISPR
— опосредованный активатор. Если соединить домен d
Cas9
(который изменён таким образом, что он больше не режет ДНК, но все ещё может находить и связываться с конкретными последовательностями ДНК) с активатором транскрипции (таким как p65HSF1
), то можно с большой точностью изменять эндогенную экспрессию конкретных генов.
Например, активируя только один эндогенный ген Sox2 или Oct4 удалось получить из фибробластов мыши ИПСК с выходом в 0,1 %
. Пользуясь подобным методом, Вэй и др. усиливали экспрессию эндогенных генов
и
, воздействуя на них с помощью CRISPR-опосредованных активаторов, и таким образом, сумели осуществить прямое репрограммирование мышиных эмбриональных стволовых клеток в две внезародышевые линии, а именно в типичные
трофобласты
и клетки внеэмбриональной
энтодермы
. Аналогичным образом активация эндогенных генов Brn2, Ascl1, и Myt1l позволила преобразовать эмбриональные фибробласты в индуцированные нервные клетки
.
Перепрограммирование путём поэтапного моделирования процессов регенерации
Ещё один способ перепрограммирования заключается в поэтапном моделировании на скелетной мышце млекопитающих процессов, которые происходят у амфибий при регенерации конечности. Таким образом, с помощью химических веществ: миосеверина (myoseverin), реверсина (2-(4-морфолиноанилино)-6-циклогексиламинопурина) и некоторых других веществ, в условиях культуры мышечных клеток млекопитающих, которые, как известно, не способны регенерировать конечности, удалось индуцировать процессы аналогичные тем которые протекают при регенерации конечностей у амфибий и получить предшественники мышечных, костных, жировых и нервных клеток
.
Получение ИПСК и трансдифференцировка с помощью антител
Обнаружены моноклональные антитела, способные преобразовывать стволовые клетки костного мозга непосредственно в прогениторные клетки нейронов головного мозга
.
Для этой трансдифференцировки как оказалось достаточно всего одного белка — антитела имитирующего фактор
. Для поиска подобных антител используется специальный метод селекции антител
.
Идентифицированы антитела, которые могут во время перепрограммирования эмбриональных фибробластов мыши в ИПСК соответственно заменить три из четырёх факторов перепрограммирования Sox2, c-Myc или Oct4. Найти замену четвёртому фактору Klf4 пока не удалось. Более того Sox2-замещающее антитело действуя как антагонист к связанному с мембраной белку
, тем самым активирует подавленные им ядерные факторы
,
и
Lin28a
(Lin28) независимо от Sox2
.
Перепрограммирование бактериями
Желудочно-кишечный тракт человека колонизирован обширным сообществом бактерий
симбионтов
и
комменсалов
. Исследователи продемонстрировали феномен перепрограммирования соматических клеток бактериями и выработку мультипотентных клеток из клеток кожи человека, под воздействием
молочнокислых бактерий
. Выяснилось, что подобная клеточная трансдифференцировка вызвана
рибосомами
и «может произойти под воздействием бактерий, которые проглатываются и перевариваются клетками-хозяевами, что приводит к стрессу от попадания чужеродных рибосом и стимулирует клеточную пластичность».
Условно перепрограммированные клетки (УПК)
Ричард Шлегель и его исследовательская группа разработали метод
, который позволяет размножать in vitro культуру клеток похожих на взрослые стволовые клетки, без каких-либо генетических манипуляций. Они показали, что под воздействием облучённых фибробластов (см. обзоры
и
) и ингибиторов Rho киназы, таких как:
или
фасудил
, первичные эпителиальные клетки млекопитающих переходят к состоянию неограниченной пролиферации
(что, по мнению авторов, связано с ростом концентрации
β-катенина
в ядре и снижением Notch сигнализации). Индукция УПК происходит довольно быстро (в течение 2 дней) и является результатом «перепрограммирования» всей клеточной популяции, а не одной из её субпопуляций. При этом в УПК не наблюдалась характерная для ИПСК или эмбриональных стволовых клеток (ЭСК) активация синтеза Sox2, Oct4, Nanog, и Klf4. Эта индукция УПК обратима — достаточно удалить Y-27632 и облучённые фибробласты, чтобы клетки перешли к обычной дифференцировке
.
Обнаружено, что факторы, вызывающие индукцию условно перепрограммированных клеток, переходят из «питающих» клеток подложки в культуральную среду в результате обусловленного радиацией
апоптоза
этих клеток.
Этот метод может иметь большое будущее в
регенеративной медицине
, так как эти клетки в отличие от ИПСК не образуют опухоли
. Так, например, используя технологию условно-перепрограммированных клеток, исследователи смогли найти эффективную терапию для пациента с редким типом опухоли лёгких
.
Иной подход к получению условно перепрограммированных клеток заключается в ингибировании мембранного белка
CD47
, являющегося рецептором
тромбоспондина-1
. Показано, что потеря CD47 снимает запрет на устойчивую пролиферацию первичных мышиных эндотелиальных клеток, повышая частоту их асимметричного деления, а также позволяет этим клеткам спонтанно перепрограммироваться в мультипотентные клетки формирующие
.
Нокдаун гена
CD47 резко увеличивает в клетках уровни мРНК с-Мус и других факторов перепрограммирования Яманаки как in vitro, так и in vivo. Очевидно, тромбоспондин-1 является ключевым сигналом
нишы
, который подавляет способность стволовых клеток к самообновлению влияя на них через CD47. Поэтому антагонисты CD47 могут активировать самообновление и перепрограммирование клеток выключая механизмы негативной регуляции с-Мус и других факторов транскрипции стволовых клеток
По мнению авторов исследования, образующиеся при этом мультипотентные клетки не образуют тератом.
In vivo блокада CD47 с помощью антисмыслового
морфолино
повышает выживаемость мышей, тело которых подверглось воздействию летальной дозы облучения. Эта устойчивость к радиации обусловлена увеличением пролиферативной способности клеток крови, образующихся из костного мозга, и активации защитной
аутофагии
радиочувствительных желудочно-кишечных тканей.
Косвенное перепрограммирование клеток (ILC)
Разработан метод при котором соматические клетки переходят в промежуточное пластическое состояние- частично перепрограммированные ИПСК (pre-iPSC), индуцированное кратковременным воздействием перепрограммирующих факторов, а затем дифференцируются с помощью специально разработанной химической среды (искусственной ниши).
Предполагается, что этот новый метод может быть более эффективным и безопасным, так как он, по мнению его авторов, не вызывает опухоли или другие нежелательные генетические изменения, и при этом позволяет получать требуемые клетки быстрее и c гораздо большим выходом по сравнению с другими методами. Тем не менее, безопасность этих клеток все же сомнительна — учитывая то, что преобразование из пре-ИПСК опирается на использование условий перепрограммирования в ИПСК, и нельзя исключить что часть клеток может все же приобрести плюрипотентные свойства(если они не прекратят процесса де-дифференцировки in vitro или в связи с дальнейшей де-дифференцировкой in vivo).
Перепрограммирование воздействием на гликопротеин наружней мембраны
Общей особенностью, взятых из разных источников, плюрипотентных стволовых клеток, отличающей их от большинства (исключение составляют лейкоциты) неплюрипотентных клеток, является особый характер гликозилирования белков их наружней мембраны
.
Расположенные на поверхности стволовых клеток
гликаны
быстро реагируют на изменения в состоянии клетки и поэтому идеально подходят в качестве маркеров для выявления изменений в клеточных популяциях. Многие широко используемые
(в том числе
, SSEA-4, TRA 1-60, и Тра 1-81.) являются гликанами клеточной поверхности
. Так, например, гликопротеин подокаликсин (podocalyxin) расположен исключительно только на недифференцированных клетках человека (ИПСК и ЭСК), но не на поверхности дифференцированных соматических клеток, что позволяет отделить эти клетки с помощью лектина BC2L-C из Burkholderia cenocepacia (rBC2LCN).
Suila Хели и соавторы
полагают, что у стволовых клеток человека внеклеточные о-GlcNAc и о-LacNAc, выполняют решающую роль в тонкой настройке
— высококонсервативной системы
клеточной сигнализации
, от которой зависят судьбы стволовых клеток, их дифференцировка, лево- и правосторонняя асимметрия, апоптоз, и пролиферация (см. обзоры:
)
Очевидно, изменения характера
гликозилирования
белков наружней мембраны являются маркерами состояния клетки каким-то образом связанными с плюрипотенцией и дифференцировкой
. Причём «сдвиг» в характере гликозилирования, по всей видимости, это не просто результат инициализации экспрессии генов, а механизм играющий роль важного регулятора группы генов, вовлечённых в приобретении и поддержании недифференцированного состояния
. Так, показано, что активация
гликопротеина
ACA
, связывающего
гликозилфосфатидилинозитол
на поверхности прогениторных клеток периферической крови человека, посредством сигнального каскада
PI3K/Akt/mTor/PTEN
индуцирует повышение экспрессии генов
Wnt
,
Notch1
,
Bmi-1
и
HoxB4
, а также способствует образованию и самообновлению популяции гемопоэтических стволовых клеток
. Более того показано, что индуцируемая посредством ACA-зависимого сигнального пути дедифференцировка прогениторных клеток, приводит к образованию ACA-индуцированных плюрипотентных стволовых клеток, способных дифференцироваться in vitro в клетки всех трёх
зародышевых листков
.
. Изучение избирательно связывающих гликопротеины
лектинов
, на предмет их способности поддерживать культуру плюрипотентных стволовых клеток человека, привело к открытию лектина эритрина кристагалли (Erythrina Cristagalli — ЕСА) способного служить в качестве простой и высокоэффективной матрицы для культивации человеческих плюрипотентных стволовых клеток
Перепрограммирование с помощью протеогликана
Альтернативной стратегией превращения соматических клеток в плюрипотентные состояния может быть непрерывная стимуляция фибробластов одним из
протеогликанов
ECM
, а именно
. Такие клетки проявляют способность к регенерации скелетных мышц с заметно меньшим онкогенным риском по сравнению с ИПСК
. Пониженная онкогенность таких клеток связана с активацией
(ингибитора
циклин-зависимой киназы
2B) во время процесса перепрограммирования рекомбинантным человеческим фибромодулином
.
Индуцированные стрессом стволовые клетки (ИССК)
Клетки STAP (Stimulus-triggered acquisition of pluripotency)
В 2014 году группа японских исследователей опубликовала в статью в журнале Nature
, где было заявлено открытие нового способа быстрого перепрограммирования соматических клеток млекопитающих в плюрипотентные клетки — так называемые клетки
в ответ на действие сильных внешних раздражителей, таких как временное повышение кислотности окружающей среды. Однако другим исследователям не удалось воспроизвести эти результаты. Впоследствии материал о клетках STAP был отозван журналом Nature как ошибочный
, один из соавторов работы покончил жизнь самоубийством
, а сами работы по этому направлению были прекращены
.
Плюрипотентные клетки содержат
E-кадгерин
, который при дифференцировке заменяется на
N-кадгерин
. Уникальной особенностью Е-кадгерина, помимо того, что он ответственен за межклеточную адгезию, является способность регулировать сигнальные пути клетки и заменять фактор Oct4 при индукции плюрипотентности
. Фибробласты в которых подавлен синтез E-кадгерина не могут перепрограммироваться. Во время перепрограммирования, N-кадгерин может заменять функции E-кадгерина, что предположительно указывает на необходимость адгезии для перепрограммирования
. Однако, согласно Гуаньнань Су с соавт., формирование в культуре клеток 3D сфер, в связи с вынужденным ростом клеток на поверхности с низкой связывающей способностью, иногда приводит к репрограммированию клеток. В качестве примера они показали, что нервные клетки-предшественники могут быть получены непосредственно из фибробластов путём физического воздействия, без введения экзогенных перепрограммирующих факторов.
Ранее подобные сферы были получены в опытах с фибробластами мыши с мутацией инактивирующей
ген-супрессор опухолей
ретинобластомы — RB1
, белка без которого клетки теряют способность к старым контактам и контактному ингибированию пролиферации в результате чего выходят за пределы колонии и образуют сферы где доминируют новые межклеточные контакты, по всей видимости, и вызывающие самопроизвольное перепрограммирование в тератомоподобные стволовые клетки
.
В
биореакторной
культуре сдвиг жидкости при перемешивании индуцирует повышенную экспрессию генов маркеров плюрипотентности, которую можно подавить ингибируя
β-катенин
или
.
Физические сигналы, в виде параллельных микродорожек на поверхности подложки для культивации клеток, могут заменить действие низкомолекулярных эпигенетических модификаторов и значительно повысить эффективность перепрограммирования. Механизм основан на механомодуляции изменяющей морфологию и эпигенетическое состояние клеток. В частности, по мнению авторов исследования: «снижение активности гистоновой дезацетилазы и повышение экспрессии WD повторяющего домена 5 (WDR5)-субъединицы H3 метилтрансферазы вызванное поверхностью с микродорожками приводит к увеличению ацетилирования и метилирования гистона Н3». Аналогичное действие на клетки оказывали нановолоконные подложки с выровненной ориентацией волокон
.
Схема адгезии клеток
Важным биофизическим фактором влияющим на дифференцировку клеток является жёсткость подложки. Например, мягкие субстраты способствуют образованию из ЭСК, по
-зависимому пути, нейроэпителиальных клеток, в то же время предотвращают дифференцировку в клетки
нервного гребня
. Исследования показали, что в этом механизме задействованы механочувствительное
Smad
фосфорилирование
и
ядерно-цитоплазматические
перемещения, зависящие от регулируемой жёсткостью подложки активности
/
и сократительной способности комплекса
актомиозин
-
цитоскелет
.
Помогает клетке преобразовывать механические раздражители в электрические и биохимические сигналы белок регулирующий открытие ионного канала Са++ названный Пьезо1 (
), который активируется натяжением мембраны. В зависимости от липидного состава мембран придающего ей жёсткость или мягкость меняется и способность Пьезо реагировать на механические стимулы
Механизмы механомодуляции см. в обзорах:
Разработан метод, который превращает соматические клетки в стволовые клетки «сжимая» их с помощью 3D микроокружения состоящего из специально подобранного геля, что открывает путь для крупномасштабного производства стволовых клеток для медицинских целей
.
Как отмечено выше, в процессе перепрограммирования клетки морфологически изменяются, что приводит к изменению их способности к адгезии. Эти характерные различия в адгезии позволили разработать процесс выделения плюрипотентных стволовых клеток с помощью
микрожидкостных
устройств
. Преимуществом этого метода является то что: разделение занимает менее 10 минут, при этом удаётся получить более чем на 95 % чистую культуру ИПСК клеток, причём выживаемость клеток больше 80 % и полученные клетки сохраняют нормальные транскрипционный профиль, потенциал дифференцировки и кариотип.
Индуцированные нервные стволовые клетки (иНСК)
Центральная нервная система млекопитающих имеет крайне ограниченные возможности для регенерации. Поэтому для лечения многих нервных расстройств (таких как: инсульт, болезнь Паркинсона, болезнь Альцгеймера, боковой амиотрофический склероз и т. д.) требуются нервные стволовые клетки, автологичным источником которых могут стать иНСК пациента. В ряде новейших публикаций описано прямое преобразование соматических клеток в индуцированные нервные стволовые клетки
.
Так, например, предшественники нервных клеток можно получить прямым преобразованием и без введения экзогенных транскрипционных факторов, пользуясь только химическим коктейлем
. Эти клетки, называемые ciNPCs (chemical-induced neural progenitor cells) можно к примеру получить из фибробластов кончика хвоста мыши или мочевыводящих соматических клеток человека, используя для этого коктейль состоящий из:
Аналогичным образом без введения экзогенных транскрипционных факторов, пользуясь только химическим коктейлем можно получить
Шванновские клетки
.
По некоторым данным, в принципе, возможно, преобразовать трансплантированные в мозг мыши фибробласты и астроциты человека, спроектированные методами генной инженерии на выработку факторов (Ascl11, Brn2a и Myt1l) индуцирующих их перепрограммирование в нейроны, активируя после трансплантации соответствующие гены с помощью активатора добавленного к питьевой воде животных.
Было также показано, что in situ эндогенные
астроциты
мыши могут быть напрямую преобразованы в функциональные нейроны
, способные участвовать в формировании
нейросетей
. В отличие от ИПСК, полученные таким образом клетки не пролиферируют, а значит более безопасны. Наблюдения за подвергшимися этой процедуре мышами в течение года, не выявили у них признаков образования опухоли. Те же исследователи превратили астроциты спинного мозга в прогениторные клетки, называемые
нейробластами
, которые способны дифференцироваться в нейроны при поврежденнии спинного мозга
.
В то время как нейроны взрослого человека обычно не в состоянии регенерировать после травмы спинного мозга, нейроны, полученные из человеческих ИПСК, после трансплантации крысам с травмами спинного мозга продемонстрировали значительный рост по всей длине центральной нервной системы животных. В эксперименте были использованы ИПСК полученные из клеток кожи, взятых от 86-летнего мужчины. Авторы исследования продемонстрировали, что полученные из ИПСК омоложённые нейроны способны прожить в костном мозге крысы не менее трёх месяцев и в течение этого срока не образовывали опухолей. Однако, такая
клеточная терапия
не привела к излечению крысы от паралича.
Inoue и его коллеги трансплантировали
глиальные нервные клетки
-предшественники, полученные из человеческих иПСК в поясничный отдел спинного мозга мышей с моделью
бокового амиотрофического склероза (БАС)
. Трансплантированные клетки дифференцировались в
астроциты
и продлевали жизнь мышей с БАС. Очевидно ИПСК могут стать перспективным ресурсом для трансплантационной терапии БАС.
Разработана технология для прямого преобразование фибробластов в функциональные астроциты с помощью транскрипционных факторов
,
и
Как показано в обзоре Бельмонто с соавт. способы прямого преобразования соматических клеток в индуцированные нервные стволовые клетки отличаются по своим методическим подходам
. Какой из этих подходов окажется наиболее приемлемым для клиники покажут исследования.
Прогениторные клетки олигодендроцитов (ПКОД)
Без
миелиновой
оболочки, выполняющей роль изоляции волокон
нейронной сети
, сигналы посланные по нервам быстро затухают. Поэтому при заболеваниях, связанных с потерей миелиновой оболочки, таких как
рассеянный склероз
, наблюдается снижение
интеллекта
,
парез
,
атаксия
туловища и конечностей, нарушения зрения, потеря чувствительности и ряд других неврологических симптомов. Перспективным подходом к лечению подобных заболеваний является трансплантация клеток-предшественников
олигодендроцитов
(ПКОД), способных заново создать миелиновую оболочку вокруг поражённых нервных клеток. Для такой терапии необходимо иметь доступный источник этих клеток.
Основу для решения этой проблемы заложил метод прямого преобразования фибробластов мышей и крыс в олигодендроглиальные стволовые клетки индуцированного путём принудительной гиперэкспрессии восьми
или всего трёх транскрипционных факторов Sox10, Olig2 и Zfp536.
Показано, что аутологичная клеточная терапия с использованием полученных
in vitro
из ИПСК пациента клеток-предшественниц олигодендроцитов приводит к миелинизации
in vivo
, что свидетельствует о функциональности этих человеческих клеток в организме мыши и об открывшейся перспективе их использования в клинике.
Индуцированные кардиомиоциты (иКМ)
Одной из наиболее актуальных задач клинической науки нынешнего столетия является развитие терапевтических стратегий, способных обратить вспять прогрессирование сердечной недостаточности — основной причины инвалидности и смертности населения. Большие надежды в этом плане возлагаются на методы клеточной терапии, которые могли бы предотвратить образование соединительной рубцовой ткани (фиброз) вместо мышечной. Простейшим подходом к решению этой задачи могло бы быть перепрограммирование сердечных фибробластов непосредственно в организме путём доставки в сердце факторов транскрипции
или микроРНК
. Была предпринята попытка перепрограммировать сердечные фибробласты в кардиомиоцит-подобные клетки in vivo путём гиперэкспрессии в них факторов транскрипции Gata4, Mef2c и Tbx5 (GMT)
. В случае удачи, такой поход позволил бы превращать рубцовую ткань в мышечную непосредственно в сердце, без необходимости клеточной трансплантации. Эффективность такого перепрограммирования оказалась очень низкой, а фенотип полученных кардиомиоцитов существенно отличался от фенотипа нормальных зрелых кардиомиоцитов. Результатом чего явилась низкая выживаемость перепрограммированных клеток
. Позднее в опытах in vitro фенотип удалось несколько исправить (добавлением ESRRG, MESP1, Myocardin, ZFPM2 и TGF-β), но эффективность перепрограммирования осталась низкой
. Поднять эффективность перепрограммирования
in vivo
позволяют неинтегрирующиеся векторы вируса Сендай, с вектором факторов перепрограммирования Gata4, Mef2c, и Tbx5
Более того предпринята успешная попытка лечения
инфаркта миокада
in vivo
путём улучшения регенерации сердца и улучшения работы миокарда после инфаркта, опосредованной полицистронным вектором Mef2c/Gata4/Tbx5 (MGT), эффективность которого подняли с помощью деградатора
BRD4
BETd-246
.
Определённые успехи наметились в методах получения и выращивания большого количества кардиомиоцитов in vitro
. Так, например, удалось с высокой степенью эффективности получить из ИПСК человека прогениторные сердечные клетки способные, при трансплантации их в сердечную мышцу, снизить её перерождение в рубцовую ткань после инфаркта
. С помощью малых молекул и путём активации синтеза
β-катенина
или же ингибирования синтеза
Wnt
в ИПСК человека in vitro, удалось повысить эффективность получения кардиомиоцитов до 80 %
.
Возможно, что в будущем удастся заменить искусственные
электрокардиостимуляторы
, необходимые людям с медленным или
нерегулярным сердцебиением
, на биологический кардиостимулятор (
пейсмекер
) из индуцированных стволовых клеток. Надежду на это вселяют эксперименты в которых поросятам делали инъекцию индуцированных сердечных клеток, способных синхронизировать ритм сердцебиения
. Более того, при ишемической кардиомиопатии, вызванной смоделированным на мышах инфарктом миокарда, трансплантация ИПСК способствовала синхронизации повреждённых желудочков сердца, улучшая их проводимость и сократимость за счёт активации процессов восстановления
.
Перепрограммированием соматических клеток
in vivo
с помощью эмбрионального фактора транскрипции
можно преобразовать кардиомиоциты в клетки пейсмекера. Это открытие открывает возможность легко и быстро вылечить пациентов зависящих от кардиостимулятора. Перенос гена TBX18
In situ
с помощью инъекции его аденовирусного носителя позволяет создать в месте инъекции естественный источник биологического водителя ритма уже через 2-3 дня после введения. При этом пока не наблюдалось возникновения опухолей или каких-либо нарушений в деятельности сердца. Таким образом, минимально инвазивный перенос генов TBX18 может рассматриваться как перспективный метод лечения больных с блокадой сердца, который в будущем, очевидно, заменит лечение искусственными кардиостимуляторами.
Создан коктейль для прямой (без прохождения через плюрипотентное состояние) трансдифференцировки, состоящий из четырёх низкомолекулярных компонентов (SB431542 (ингибитора ALK4/5/7), CHIR99021 (ингибитора
GSK3
), парната (ингибитора LSD1/KDM1, называемого также
транилципромином
), и форсколина (активатора
аденилатциклазы
)). Этот коктейль позволил с высокой эффективностью превратить фибробласты мыши в клетки сердечной мышцы с помощью всего одного фактора транскрипции — Oct4. Полученные таким способом индуцированные кардиомиоциты спонтанно сокращались
. Методом прямой трансдифференциации без использования генетических векторов, то есть чисто фармакологически, с помощью коктейля из девяти компонентов, удалось получить с выходом 97 % из фибробластов кожи бьющиеся химически индуцированные кардиомиоцит-подобные клетки (ciCMs), которые почти не отличались от человеческих кардиомиоцитов по данным исследования их транскриптома, эпигенетически и по электрофизиологическим параметрам. Более того, при трансплантации в сердце мыши с инфарктом, обработанные этим коктейлем фибробласты превращались в выглядевшие здоровыми клетки сердечной мышцы
. Предпринята успешная попытка противостоять постинфарктному
фиброзу
(перерождению сердечной мышцы в соединительную ткань с образованием
рубца
) с помощью химического перепрограммирования in vivo сердечных фибробластов в кардиомиоциты.
Лу с соавторами
создали биоинженерную конструкцию сердца путём заселения очищенного от клеток (децеллюларизованного) сердца мыши мультипотентными сердечно-сосудистыми прогениторными клетками, полученными из ИПСК человека. Они обнаружили, что мультипотентные сердечно-сосудистые прогениторные клетки направленно мигрируют в соответствии с архитектурой сердца, а прибыв на место, размножаются и дифференцируются в кардиомиоциты, клетки гладких мышц и эндотелиальные клетки, как это необходимо для восстановления утраченной структуры сердца. Очевидно, что внеклеточный матрикс сердца мыши (оставшаяся после удаления клеток мыши подложка сердца) может посылать сигналы мультипотентным сердечно-сосудистым прогениторным клеткам человека, необходимые для их навигации и превращения в специализированные клетки, обеспечивающие нормальную работу сердца. Через 20 дней после перфузии сердца средой содержащей факторы роста, оно, после электростимуляции, начинало биться с темпом 40-50 ударов в минуту и реагировало на медикаменты.
Подробнее в обзорах:
Созревание кардиомиоцитов in vivo
Кардиомиоциты получаемые из ИПСК отличаются от взрослых соматических клеток и остаются незрелыми при культивировании в чашках Петри. Японским учёным удалось добиться созревания этих клеток. Для этого они на месяц поместили незрелылые клетки кардиомиоцитов в сердце новорождённой мыши для дозревания
.
Омоложение мышечных стволовых клеток
Пожилые люди нередко страдают от прогрессирующей
дистрофии
и слабости мышц, что отчасти связано с повышенной активностью сигнальных путей
p38α и p38β митоген-активированных протеин киназ
в стареющих мышечных стволовых клетках. Подвергнув такие стволовые клетки непродолжительному воздействию SB202190 — ингибитора p38α и p38β — в сочетании с культивированием на мягкой подложке из
гидрогеля
, можно быстро омолодить их и размножить. Более того, после имплантации в организм такие омоложённые клетки способны повысить силу старых мышц
.
Восстановить способность сателлитных стволовых клеток к регенерации можно и подавив синтез на гене
(называемом также Cdkn2a)
.
Миогенные предшественники, которые могут быть использованы для моделирования болезней или клеточной терапии скелетных мышц, могут быть также получены непосредственно из ИПСК с помощью свободно-плавающей шаровой
культуры
(EZ сфер) в культуральной среде, содержащей высокие концентрации (100 нг / мл)
фактора роста фибробластов-2 (FGF-2)
и
эпидермального фактора роста
.
Индуцированные гепатоциты
Получение клеток печени из ИПСК
Гепатоциты
человека имеют очень ограниченную способность к восстановлению после повреждений печени. Поэтому трансплантация печени нередко является единственным способом лечения таких болезней как цирроз. Клеточная терапия печени затрудняется тем, что культура гепатоцитов плохо размножается in vitro.
Поэтому удобнее размножить клетки в виде ИПСК, и только затем превратить их в гепатоциты.
Разработано несколько способов получения гепатоцитов из ИПСК
Так, например для очистки и размножения самообновляющихся гепатобласт-подобных клеток из человеческих плюрипотентных стволовых клеток (ЭСК/ИПСК), их культивировали на чашках покрытых человеческим ламинином-111 в течение более 3 месяцев, после чего они подобно овальным клеткам печени были способны дифференцироваться в гепатоцит-подобные клетки, а также в клетки жёлчных путей —
холангиоцит
-подобные клетки. Было показано, что такие гепатобласт-подобные клетки могут интегрироваться в паренхиму печени мыши. Предполагается, что, благодаря подавлению неблагоприятных генных регуляторных сетей при культивировании на поверхности покрытой ламинином, гепатоциты имеют большое сходстве со взрослыми гепатоцитами и могут быть использованы для скрининга лекарственных средств, а также в качестве источника клеток для регенеративной терапии печени
.
В 2010 году была продемонстрирована возможность индуцировать полученные из жировой ткани стромальные клетки (ASC) в клетки похожие по ряду функций на гепатоциты человека, способные прижиться в повреждённой токсинами печени мыши
. Позднее был разработан быстрый (до десяти дней) и эффективный (с выходом более 50 процентов) способ превращать клетки полученные путём липосакции в клетки печени. Клетки, полученные из собственных клеток человека с помощью этой новой методики, превращаются в клетки печени без промежуточной фазы плюрипотентных клеток и, очевидно, не образуют опухоли. В печени они формируют многоклеточные структуры необходимые для образование жёлчных протоков. Особенностью этой методики является культивирование адипоцитов в жидкой суспензии, в которой они образуют сфероиды
Обнаруженная у совместной культуры гепатоцитов, полученных из ИПСК, с эндотелиальными (для образования сосудов) и мезенхимальными (для образования поддерживающего
внеклеточного матрикса
) клетками, способность к самоорганизации (
самосборке
) в трёхмерные шарообразные структуры, представляющие собой зачаток печени
позволяет надеяться, что в будущем трансплантологам: не надо будет искать и ждать донора, больному будут пересаживать зачаток нужного органа, полученный из его же собственных клеток, и этот зачаток будет уже на месте дорастать до нужных размеров.
Эта методика позволяет использовать клетки всего одной мыши для предварительной проверки 1.000 лекарственных препаратов на их пригодность для лечения болезней печени, что открывает новые возможности для медицинских исследований и проверки безопасности лекарств
.
Методы получения гепатоцитов без использования ИПСК
Для получения
гепатоцитов
из человеческих фибробластов не обязательно вначале получить ИПСК. Используя небольшие молекулы можно добиться прямого перехода фибробластов в индуцированные мультипотентные прогениторные клетки (iMPC) из которых затем образуются сначала прогениторные клетки эндодермы, а затем гепатоциты. После трансплантации мышкам с иммунодефицитом и смоделированным поражением печени, клетки iMPC интенсивно размножаются и приобретают функциональные способности характерные для взрослых гепатоцитов. При этом не наблюдалось образование опухолей, потому что клетки не проходили через стадию плюрипотентного состояния
. С помощью инфицирования
лентивирусами
, вызывающими экспрессию генов
,
и
HNF4A
, удалось осуществить прямое преобразование фибробластов человека во взрослые гепатоцито-подобные клетки, которые могут быть размножены в культуре, а затем использованы для лечения острой печёночной недостаточности и метаболической болезни печени.
.
Инактивация сигнального пути Hippo in vivo с высокой эффективностью приводит к дедифференцировке взрослых гепатоцитов в клетки, несущие характеристики прогениторных клеток. Эти клетки-предшественники продемонстрировали способность к самообновлению и смогли прижиться в печени. Эти данные продемонстрировали беспрецедентный уровень фенотипической пластичности зрелых гепатоцитов
Коктейль из малых молекул, Y-27632, A-83-01 и CHIR99021, может превратить зрелые гепатоциты крысы и мыши in vitro в пролиферативные бипотентные клетки — CLiPs (chemically induced liver progenitors — химически индуцированные клетки-предшественники печени). CLIPS могут дифференцироваться как в зрелые гепатоциты, так и в эпителиальные клетки жёлчных протоков, которые могут образовывать функциональные структуры протоков. При длительном культивировании CLIPS не теряют свою пролиферативную активность и способность дифференциации в клетки печени, и могут заселять хронически поражённые ткани печени
.
Подробнее см. обзор:
Крипта кишечника
. На дне крипт располагаются недифференцированные
бескаёмчатые энтероциты
являющиеся наиболее подходящим и доступным источником для перепрограммирования в инсулин-продуцирующие клетки
Избыточная экспрессия комбинации трёх
транскрипционных факторов
(
,
и
) называемой PNM, способна привести к трансформации некоторых видов клеток в состояние подобное β-клеткам.
Оказалось, что наиболее подходящим и доступным источником для перепрограммирования в инсулин-продуцирующие клетки, является
эпителий
кишечника
. Под действием PNM трёхмерная культура зачатков органа (так называемые
органоиды
) стимулирует превращение эпителиальных клеток кишечника в β-подобные клетки, которые можно использовать для трансплантации
.
Биоинженерия клеток кровеносных сосудов
Кровеносные сосуды образуют обширные сети, которые в течение всей жизни обеспечивают клетки организма питательными веществами и кислородом. Когда кровеносные сосуды становятся старше, их структура и функции, нередко, отклоняются от нормы, способствуя тем самым многочисленным возрастным заболеваниям, таким как: инфаркт миокарда, ишемический инсульт и атеросклероз артерий, питающих сердце, мозг и нижние конечности. Поэтому, важной задачей является стимулирование роста сосудов для обеспечения циркуляции, чтобы предотвратить обострение этих заболеваний. Одним из способов стимулирования роста сосудов является имплантация индуцированных прогениторных клеток
эндотелия
(иПЭк).
Так, например, с помощью иПЭк, полученных путём частичного репрограммирования клеток эндотелия, удалось добиться увеличения коронарного кровотока и по данным эхокардиографии улучшить функционирование сердца
. Стволовые клетки, извлечённые из жировой ткани после
липосакции
можно превратить в прогениторные
гладкие мышечные клетки
(иПГМк), участвующие в формировании артерий и вен. Это клетки могут быть использованы для создания кровеносных сосудов, необходимых для замены неисправных артерий сердца
.
Так, например, обнаружено, что с помощью культуры ИПСК человека в сочетании с селекцией с помощью трёх маркеров: CD34 (поверхностного гликофосфопротеина ранних эмбриональных фибробластов), NP1 (рецептора — нейрофилин1) и KDR (киназы содержащей домен рецептора), удалось получить эндотелиальные клетки, которые после трансплантации мышам образовали in vivo стабильные функциональные кровеносные сосуды, работавшие на протяжении по меньшей мере 280 дней.
.
При лечении
инфаркта миокарда
важно предотвратить образование
фиброзных тканей шрама
и стимулировать регенерацию. Достичь этого in vivo можно применив паракринные факторы способные изменить направление дифференцировки сердечных стволовых клеток предшественников от специализации в фиброзную рубцовую ткань в сторону образования сердечно-сосудистой ткани. Например, на мышиной модели инфаркта миокарда, было показано, что однократная интрамиокардиальная инъекция мРНК
фактора роста эндотелия сосудов
(VEGF-A modRNA), синтетически модифицированной так чтобы предотвратить её деградацию организмом, приводит к длительному улучшению функции
сердца
, обусловленному перенаправлением дифференцировки эпикардиальных клеток-предшественников в сердечно-сосудистый тип клеток
.
Мервин Иодер с соавт., описали метод для преобразования ИПСК человека в клетки, подобные эндотелиальным колониеобразующим клеткам
пуповинной крови
(CB-ECFCs). Полученные ими CB-ECFC-подобные клетки имели стабильный эндотелиальный фенотип, высокий пролиферативный потенциал и способность, при трансплантации мышкам, образовывать человеческие кровеносные сосуды, а также участвовать в регенерации сетчатки и конечностей мыши после ишемии. Индуцированные CB-ECFC-подобные клетки практически не образуют тератомы
.
Прямое перепрограммирование клеток взрослого организма в прогениторные нефроны (ИПН)
Взрослые клетки проксимальных канальцев
почки
могут быть непосредственно перепрограммированы в прогениторные
нефроны
эмбриональной почки, с использованием пула из шести генов кодирующих «инструктирующие» факторы транскрипции (SIX1, SIX2, OSR1, Eyes absent homolog 1(EYA1), Homeobox A11 (HOXA11) и Snail homolog 2 (SNAI2)).
Возможность получения таких клеток позволит в будущем приступить к разработке методов клеточной терапии почечных заболеваний. Первые успехи на этом пути уже есть. Так, недавно было показано, что эмбриональные органоиды почки, сформированные путём самоорганизации из клеточной суспензии, после трансплантации их во взрослую почку крысы могут в ней прижиться.
Биоинженерия стволовых клеток крови
Одной из самых востребованных целей
регенеративной медицины
является возможность получения в неограниченном количестве
гемопоэтических стволовых клеток
, пригодных для трансплантации, из более зрелых или дифференцированных клеток крови, для того чтобы покрыть дефицит
трансплантатов
костного мозга
. Чтобы запустить в фибробластах процессы
гемопоэза
в условиях
in vitro
достаточно всего четырёх
транскрипционных факторов
: Gata2, Gfi1b, cFos, и Etv6 . Их воздействие приводит к образованию клеток подобных
эндотелиальным
—
прогениторным клеткам
с последующим возникновением из них кроветворных клеток
. Аналогичным образом, используя 6 транскрипционных факторов: Run1t1, Hlf, Lmo2, Prdm5, Pbx1, и Zfp37, а также ещё два фактора Mycn и Meis1 для повышения эффективности перепрограммирования, удалось получить
гемопоэтические стволовые клетки
из зрелых дифференцированных клеток крови
.
См. также обзоры:
Эритроциты
Переливание
эритроцитов
необходимо для многих пациентов с травмами или гематологическими заболеваниями. Однако, на сегодняшний день, поставка эритроцитов зависит от добровольных доноров число которых недостаточно. Кроме того, переливание крови от доноров сопряжено с определённым риском из-за возможности передачи ряда инфекций. Решить эту проблему могло бы производство необходимых количеств эритроцитов вне организма
. В принципе уже доказано, что эритроциты, полученные вне организма из мобилизованных
CD34
-позитивных клеток (CD это на англ. сокращённо
кластер дифференцировки
), способны выжить при переливании
аутологичному
реципиенту
. Эритроциты, получаемые in vitro, как правило, содержат исключительно
зародышевый гемоглобин
(HbF), который непригоден для нормального функционирования эритроцитов во взрослом организме.
Тем не менее, in vivo, после трансфузии полученных из ИПСК эритроидных прогениторных клеток содержащих ядро, наблюдалось переключение на синтез
взрослой изоформы гемоглобина
. Однако в этом случае возникает другая проблема: несмотря на то, что эритроциты не имеют ядер, и, следовательно, не могут образовывать опухоли, их непосредственные предшественники эритроидные прогениторные клетки ядром обладают и следовательно потенциально опасны. Созревание
эритробластов в функционально зрелые эритроциты
требует сложного процесса реорганизации, который заканчивается удалением ядра с образованием безъядерных эритроцитов
. Увы, методы перепрограммирования клеток в настоящее время часто приводят к нарушению этих процессов энуклеации и поэтому использование эритроцитов или их непосредственных предшественников эритробластов для переливания ещё недостаточно защищено от возможности образования опухолей. Тем не менее Bouhassira и его коллеги обнаружили, что кратковременное воздействие цитокинов, благоприятствующих дифференцировке стволовых клеток в эритроидные, на CD34 позитивные клетки, до их размножения с последующей
пролиферацией
полученных предшественников, позволяет получать на порядок больший выход эритроидных клеток, чем наблюдалось ранее. И что самое главное: эти красные кровяные клетки имели те же изоформы глобина что и использованные в качестве источника CD34 позитивные клетки
. Значительно повысить выход эритроидных клеток из ИПСК или же эритроцитов из человеческих гемопоэтических стволовых клеток позволяет подавление гена
SH2B3
или его инактивация генным редактированием с помощью
CRISPR
/
Cas9
Важную роль в развитии нормальных клеток крови играет сигнальный путь рецептора арил-углеводородов (AhR) (который как было установлено, содействует и образованию раковых клеток
). Активация AhR в человеческих гемопоэтических клетках-предшественницах (HPS) приводит к беспрецедентной пролиферации HPS, мегакариоцитов и клеток эритроидных линий.
.
Подробный обзор методов получения эритроцитов см. в
Тромбоциты
Тромбоциты
играют важную роль в предотвращении кровоизлияния у больных с тромбоцитопенией или с тромбоцитемией. Серьёзной проблемой для пациентов после повторных переливаний тромбоцитов является развитие иммунных реакций. Поэтому для клиники большое значение имеет возможность получения тромбоцитов, не содержащих
HLA-антигены
, вне организма и на средах, не содержащих сыворотки. Некоторых успехов в этом направлении добились Figueiredo с соавторами. Используя
РНК-интерференцию
для подавления синтеза β2-микроглобулина в
CD34
-положительных клетках, они сумели получить тромбоциты, в которых на 85 % было снижено содержание антигенов HLA
. Позднее удалось получить неиммуногенные по HLA класса I тромбоциты, которые кроме того не активируют
NK-клетки
Разработан метод получения тромбоцитов, который заключается в создании из ИПСК человека устойчивых иммортализованных линий клеток-предшественников
мегакариоцитов
(imMKCLs) путём
доксициклин
-зависимой гиперэкспрессии
Bmi1
и
.
Полученные imMKCLs можно размножать и культивировать в течение длительного периода (4—5 месяцев), причём даже после
криоконсервации
. Прекращение сверхэкспрессии c-MYC, Bmi1 и Bcl-X L (путём удаления доксициклина из среды) заставляло эти клетки производить тромбоциты
CD42b
+, которые по большинству параметров не отличались от тромбоцитов крови
.
Альтернативный подход получения мегакариоцитов, с высоким выходом (3 единицы (2.4 × 10
11
тромбоцитов на единицу) тромбоцитов для переливания с одного миллиона клеток ИПСК) и с чистотой более 90 %, позволяет культивирование на среде без продуктов животного происхождения (а потому с достаточно определёнными, предсказуемыми условиями, что важно для получения надёжно воспроизводимых результатов). Для перепрограммирования использовалась лентивирусная трансдукция, приводящая к одновременной экзогенной экспрессии трёх факторов транскрипции:
GATA1
,
и
.
Обзор по проблемам связанным с производством тромбоцитов см.
Иммунные клетки
Вырабатываемый иммунной системой специализированный тип
белых кровяных клеток
, известный как
цитотоксические Т-лимфоциты
(CTL)
, способен распознать специфические маркеры на поверхности различных инфекционных или опухолевых клеток и уничтожить эти вредоносные клетки. Поэтому
иммунотерапия
с использованием антиген-специфических Т-клеток в будущем может быть использована для борьбы со многими видами рака и вирусных инфекций
. Организм производит очень мало таких клеток и выделить их в количестве необходимом для терапии очень сложно. Потенциально эффективным подходом получения этих клеток для терапии может быть технология заключающаяся в том чтобы превратить зрелые CTL в ИПСК, которые обладают способностью к неограниченной пролиферации in vitro, размножить эти ИПСК до необходимого количества и затем провести их дифференцировку обратно в зрелые CTL
.
Ещё большие возможности обещает метод, который сочетает две технологии — 1. получение ИПСК и превращение их в Т-клетки, и 2. последующую их генетическую модификацию, с помощью технологии конструирования химерных рецепторов антигенов (
CAR
)
, позволяющую им распознавать раковые клетки-мишени по антигенам, в частности по CD19 — антигену, синтезируемому злокачественными В-клетками
. По аналогичной технологии можно было бы создать распознающие белок
Т-клетки направленные против
устойчивых к антибиотикам бактерий
таких как
метициллин-резистентный золотистый стафилококк
.
Большой клинический потенциал в качестве
адъюванта
для иммунотерапии рака имеют
инвариантные
естественные киллеры
T (INKT)
— клетки, которые могут служить в качестве моста между
врождённой
и
приобретённой
иммунной системами. Они повышают противоопухолевую активность организма, производя гамма-
интерферон
(ИФН-γ)
. Предложен концептуальный метод использования INKT клеток, полученных из ИПСК, для терапии рака, который состоит из четырёх ступеней: (1) выделение минимального количества INKT клеток, (2) перепрограммирования этих INKT клеток в ИПСК, (3) размножение этих ИПСК в культуре и дифференцировка их обратно в INKT клетки и (4) инъекция, полученных INKT клеток, подопытным животным для терапии рака
.
Сконструирована клональная линия ИПСК с тремя генами отредактированными так, чтобы они экспрессировали: высокоаффинную, нерасщепляемую версию
Fc-рецептора
; белок
интерлейкина (IL)-15
связанный с его мембранным рецептором
и при этом у них был бы подавлен нокаутом фермент
CD38
, который гидролизует НАД+. Природные киллеры (NK), полученные из этих сконструированных ИПСК, называемых
iADAPT
, активны в условиях, когда клетки обычных естественных киллеров уже не работают и поэтому могут быть использованы для эффективного лечения пациентов с запущенным раком
.
Для терапии могут быть использованы также
дендритные клетки
, которые участвуют в контроле Т-клеточного ответа. После инъекции они могут выжить достаточно долго, чтобы стимулировать антиген-специфические CTL, после чего могут быть полностью устранены. Неисчерпаемым источником для терапии вакцинами могут служить антиген-представляющие дендритные клетки, полученные из человеческих ИПСК
или прямым перепрограммированием из фибробластов
.
В-клетки
способны к быстрой (за 2-3 дня) трансдифференцировке в макрофаги под воздействием транскрипционного фактора
.
Кроме того эффективность перепрограммирования В-клеток в ИПСК с помощью транскрипционных факторов Oct4, Sox2, Klf4 и Myc под воздействием C / EBPα возрастает 100-кратно и охватывает порядка 95 % популяции клеток.
С помощью C / EBPα можно преобразовать некоторые линии в-клеток лимфомы человека и лейкоза в макрофаг-подобные клетки уже не способные к дальнейшему онкогенезу.
Омоложение эпителиальных клеток тимуса
Тимус
является органом, размеры которого существенно сокращаются с возрастом. Это сокращение является одной из основных причин того, что
иммунная система
с возрастом становится менее эффективной. Одним из центральных звеньев механизма возрастной инволюции тимуса является снижение синтеза транскрипционного фактора
.
Клэр Блэкберн и её коллеги показали, что даже далеко зашедшая возрастная инволюция тимуса может быть обращена вспять путём принудительного усиления активности в эпителиальных клетках тимуса всего одного фактора транскрипции — FOXN1 с целью содействия омоложению, пролиферации и дифференцировки этих клеток в полностью функциональный эпителий
.
Более того они показали, что принудительная экспрессия Foxn1 позволяет перепрограммировать клетки кожи — фибробласты в функциональные эпителиальные клетки тимуса. Эти FOXN1-индуцированные эпителиальные клетки тимуса (iTECs) поддерживали эффективное развитие
in vitro
линий клеток
CD4
+ и
CD8
+ тимуса. Но, что самое главное, после трансплантации в почку мыши, iTECs собирались и образовывали полностью организованный и функциональный тимус, который содержал все подтипы эпителиальных клеток тимуса, необходимых для поддержки дифференцировки Т-клеток, в результате чего иммунная система реципиента пополнялась новыми Т-клетками.
Это открытие можно считать
первым примером
выращивания органов
из
трансплантированных индуцированных стволовых клеток
. В будущем этот метод может быть широко использован для повышения иммунной функции и борьбы с
у пациентов путём омоложения тимуса
in situ
.
Индуцированные стволовые/стромальные клетки мезенхимы (ИМСК)
Благодаря своим способностям вызывать иммуносупрессию и способности к дифференцировке во многие типы мезенхимальных тканей,
стволовые/стромальные клетки
мезенхимы
(МСК) интенсивно исследуются на предмет их применения для лечения сердца, почек, нервной ткани, суставов и регенерации костей, а также терапии воспалительных заболеваний и подавления реакции отторжения при трансплантации
. МСК, как правило, получают путём болезненных, инвазивных процедур из взрослого костного мозга или жира, при этом выход очищенных МСК составляет всего лишь 0,001 % — 0,01 % от клеток костного мозга и 0,05 % от аспирата липосакции
. На практике удобнее всего получать МСК из аспирата липосакции, при этом удаляют взрослые адипоциты, которые успели потерять способность к пролиферации. Между тем взрослые адипоциты легко можно выделить и подвергнуть дедифференцировке в так называемые дедифференцированные жировые клетки (ДДЖК), которые возвращают способность к пролиферации и мультипотентность
. При соответствующих условиях культивирования in vitro или окружения in vivo ДДЖК могут дать начало адипогенным, остеогенным, хондрогенным или миогенным прогениторным клеткам, а также стимулировать неоваскуляризацию то есть проявляют те же свойства, что и МСК костного мозга
. У пожилых пациентов, которые больше всего нуждаются в восстановлении тканей путём аутологичной клеточной терапии, с возрастом наблюдается резкое возрастное снижение количества и качества МСК и адипоцитов
. Вместе с тем, известно, что ИПСК могут быть получены путём омоложения клеток даже от столетних людей
. Поэтому ИПСК, которые можно получить перепрограммированием клеток из тканей пациента и затем практически неограниченно размножать in vitro, может стать удобным источником омоложённых МСК.
Как показали опыты на мышах с моделью
таких как
Болезнь Крона
и
Язвенный колит
, молоденькие ИМСК могут быть успешно использованы для лечения даже лекарственно-рефрактерных форм подобных воспалительных заболеваний
.
Chen с соавт. обнаружили, что воздействуя на ИПСК человека препаратом
можно достаточно быстро получить однородную культуру клеток ИМСК, которые по свойствам мало чем отличаются от молодых МСК. По мнению авторов статьи такие ИМСК не обладают способностью к образованию тератом и имеют стабильный кариотип, а поэтому могут быть использованы для терапии
В настоящее время пока мало данных об эффективности и долгосрочной безопасности полученных этим методом ИМСК in vivo. Известно только что ИМСК могут быть использованы в клинике для лечении
периодонтита
и разработки методов
ортопедии
Мезенхимальные стволовые клетки (МСК) присутствуют во всех тканях и, как известно, прямо или косвенно способствуют регенерации тканей. Однако при использовании иМСК для трансплантационной терапии существует риск образования тератом из остаточных недифференцированных иПС-клеток. Удаление таких нежелательных иПС-клеток перед трансплантацией имеет решающее значение для реализации безопасной клеточной терапии на основе иМСК. Обнаружено что обработка культуры иМСК в течение 7 дней
мощным ингибитором
избирательно удаляет недифференцированные иПС-клетки человека из иМСК, активируя остановку клеточного цикла и апоптоз, тогда как иМСК сохраняют свои свойства и потенциал дифференцировки.
Важную роль в инициировании и ускорении молекулярной программы, которая приводит к дифференциации ИМСК из ИПСК выполняет белок
.
Генетическая делеция MSX2 ухудшает дифференцировку ИМСК из ИПСК. При использовании коктейля растворимых молекул эктопическая экспрессия MSX2 способствует образованию почти однородной популяции полностью функциональных ИМСК
.
Разработан химический метод получения ИМСК из первичных фибробластов кожи человека с использованием шести химических ингибиторов (SP600125, SB202190, Go6983, Y-27632, PD0325901 и CHIR99021) с добавлением трёх факторов роста: трансформирующего фактора роста-β (TGF-β), основного фактора роста фибробластов (bFGF) и фактора подавления лейкемии (LIF). Этот химический коктейль преобразует человеческие фибробласты в ИМСК всего за 6 дней с эффективностью порядка 30-40 процентов
.
Культуры мезенхимальных стволовых клеток человека могут быть использованы in vitro для массового производства
экзосом
, которые, как выяснилось, идеально подходят в качестве средства для доставки лекарств
и для доставки в клетку-мишень факторов транскрипции или микроРНК индуцирующих перепрограммирование (дедифференцировку, дифференцировку или трансдифференцировку).
Обнаружены гены позволяющие успешно идентифицировать МСК во всех исследованных тканевых источниках, в связи с чем они могут использоваться наряду с раннее выработанными критериями
в качестве маркеров стволовых клеток.
Это шесть генов:
,
,
,
,
и
.
Все эти гены кодируют белки участвующие в работе комплекса
протеасомы
— многобелкового комплекса, разрушающего ненужные или дефектные белки при помощи протеолиза.
Индуцированные хондрогенные клетки (ИХОНК)
Хрящ соединительной ткани обеспечивает движение суставов без трения. Его дегенеративное перерождение в конечном счёте, приводит к полной потере функции сустава на поздних стадиях остеоартрита. Единственным типом клеток в хряще являются хондроциты окружённые выделяемым ими внеклеточным матриксом. В настоящее время исследователи используют два способа восстановления хрящевой ткани:
получением хондроцитов из плюрипотентных клеток (ЭСК/ИПСК)
.
получением хондроцитов прямым преобразованием фибробластов человека непосредственно в индуцированные хондрогенные клетки, минуя промежуточную стадию плюрипотентных клеток, с помощью трёх факторов репрограммирования (с-Мус, KLF4, и SOX9)
.
Преимуществом первого метода является быстрое размножение культуры исходных клеток. Преимуществом второго — отсутствие в культуре плюрипотентных клеток, которые могли бы вызвать тератому. Клетки, полученные прямым перепрограммированием, синтезировали коллаген типа II. После имплантации в область поражения они смогли выжить и в течение не менее четырёх недель участвовали в образовании хрящевой ткани у мышей.
Источники соматических клеток
Наиболее часто для перепрограммирования используют получаемые биопсией фибробласты кожи
и клетки крови
, однако удобнее получать соматические клетки из мочи
. Этот способ не требует
биопсии
или взятия образцов крови и поэтому безвреден для пациента. Стволовые клетки мочи имеют способность к мультипотентной дифференцировке. Они способны дифференцироваться в эндотелиальные, остеогенные, хондрогенные, адипогенные, скелетные миогенные и нейрогенные линии и вместе с тем не образуют тератомы.
. Поэтому их эпигенетическая память хорошо подходит для перепрограммирования в ИПСК. Вместе с тем, клеток в моче мало, эффективность их превращения в стволовые клетки низка, тогда как риск бактериального заражения выше, по сравнению с другими источниками клеток
.
Ещё одним перспективным источником клеток для перепрограммирования являются мезенхимальные стволовые клетки, полученные из фолликулов человеческого волоса.
и кератиноциты
Происхождение соматических клеток используемых для перепрограммирования может оказывать влияние на эффективность перепрограммирования
, функциональные свойства получаемых индуцированных стволовых клеток
и способность к образованию опухолей
.
ИПСК сохраняют эпигенетическую память о тканях из которых они произошли, и это влияет на их способность к направленной дифференцировке
Остаточная эпигенетическая память не обязательно проявляется на стадии плюрипотентности — ИПСК, полученные из разных тканей имеют надлежащую морфологию, в них активны гены характерные для плюрипотентности, и они способны дифференцироваться в ткани трёх эмбриональных слоёв как in vitro, так и in vivo
. Однако, эта эпигенетическая память может проявиться позже, во время повторной дифференцировки в специфические типы клеток, которая требует активации локусов, сохранивших элементы остаточной эпигенетической памяти.
Культуральная среда для плюрипотентных стволовых клеток свободная от питающих клеток и сыворотки
Для выращивания плюрипотентных стволовых клеток человека обычно используются так называемые питающие клетки (feeder cells) и сыворотка из эмбрионов быка (FBS). И то и другое является продуктами животного происхождения и может изменять свойства от партии к партии, что затрудняет стандартизацию условий. Кроме того выращивание стволовых клеток на клетках другого человека или животных создаёт риск загрязнения патогенными микроорганизмами, которые могут стать источником болезни для пациента после клеточной терапии.
. Поэтому компоненты животного происхождения требуют дорогостоящего контроля на качество, и их свободу от
патогенов
,
и
антигенов
.
Для замены питающих клеток используются различные подложки, такие как: Matrigel, CELLstart, рекомбинантные белки и синтетические полимеры такие как Synthemax (см. обзор к статье
.
Известно, что важную роль в адгезии клеток друг к другу и к внеклеточному матриксу играет тримерный белок
ламинин
. Было найдено что ламинин-511, названный так за то, что он содержит α5 , β1 и γ1 цепи
, если его нанести на дно
чашки Петри
способен поддерживать стабильную культуру ЭСК или ИПСК
. На основе этого открытия была разработана стандартная методика для длительного культивирования ЭСК и ИПСК человека в чашках покрытых rLN511E8 — рекомбинантным фрагментом ламинина −511 и с безсывороточной средой StemFit™
.
Аналогичная методика но с использованием ламинина-521 и
E-кадгерина
позволила клонировать in vitro эмбриональные стволовые клетки без необходимости использовать ингибиторы
(
англ.
)
. Интересно было бы применить её и для ИПСК.
В стадии разработки находится также очень дешёвая подложка из углеродных нанотрубок. Она позволит выращивать и проводить дифференцировку стволовых клеток в промышленных масштабах. По заверениям авторов изобретения, изменяя условия изготовления этой подложки, можно так изменить её свойства, что она будет влиять на способность выращиваемых клеток к адгезии, на их пролиферацию и морфологию образуемых клеточных колоний.
Разработана среда-коктейль CEPT, состоящая из четырёх малых молекул: chroman 1 (
)
, emricasan (ингибитор
каспазы
)
, транс-
ISRIB
.
и полиаминов таких как
спермин
, которая, улучшает жизнеспособность плюрипотентных стволовых клеток человека, защищает клетки во время культивирования и
криоконсервации
, а также способствует дифференцировке
in vitro
и образованию
органоидов
Способы доставки репрограммирующих факторов в ядро
Способы доставки можно подразделить на
вирусные
и невирусные, а также на те, что связаны с интеграцией
векторов
несущих репрограммирующие факторы в геном и действующие без интеграции
.
(Значками отмечены свойства соответствующего вектора: (+)- Геномная интеграция происходит; (±)- интеграция происходит, но очень редко; (-)- вектор не интегрируется; (тр)- после интеграции векторная конструкция должна быть удалена транспозазой.)
Доставка вирусами
Чаще всего для доставки используются вирусные
векторные
системы. Вирусы используют свой врождённый механизм заражения клетки, что позволяет использовать их для доставки и внедрения кассеты генов необходимых для экспрессии репрограммирующих факторов
В качестве вирусов для доставки генов обычно используют:
Ретровирусы
(+). Они в качестве генома содержат одноцепочечную молекулу РНК. С помощью обратной транскрипции на РНК вируса синтезируется линейная двухцепочечная ДНК которая затем интегрируется в двухцепочечную ДНК генома клетки-хозяина. Описан метод эффективного перепрограммирования клеток человека в ИПСК с помощью одного вектора содержащего четыре ТФ, в сочетании с коктейлем, содержащим три небольшие молекулы
. Приведены прописи аналогичных методов
.
Лентивирусы
(+). Они являются подклассом ретровирусов. В отличие от ретровирусных векторов, лентивирусные векторы могут заражать не только делящиеся клетки, но и находящиеся в покое терминально дифференцированные клетки
. Удаляемая полицистронная кассета STEMCCA представляющая собой вырезаемый
перепрограммирующий лентивирусный вектор позволяет осуществлять свободное от трансгенов перепрограммирование взрослых фибробластов кожи человека в ИПСК
.
Вирус Сендай (-) — вирус из семейства Paramyxoviridae, содержащий одноцепочечную РНК
. Вирус Сендай считается безопасным, потому что его генетический материал не включается в ДНК клетки, и от него достаточно легко избавиться инкубацией культуры клеток при повышенной температуре. Вирус от тепла погибает, тогда как трансформируемым клеткам такая обработка не вредит
. Для получения ИПСК этим методом можно воспользоваться готовыми наборами
. В отличие от ретровирусных и эписомных векторов, при перепрограммировании с использованием вируса Сендай пока не наблюдалось дефектных клонов, не способных к дифференцировке
. Описание метода см
.
Венесуэльский вирус лошадиного энцефалита (VEE) (-) у которого структурные белки удалены, но все ещё присутствуют неструктурные белки,
позволяет с помощью самореплицирующегося полицистронного репликона РНК внести в клетку однократной трансфекцией четыре перепрограммирующих фактора (OCT4, KLF4, SOX2 и либо c-MYC либо GLIS1) .
Не интегрирующиеся аденовирусы (±)
. По мнению некоторых авторов, векторную кассету, после того как перепрограммирование достигнуто, можно удалить с помощью трансфекции мРНК Cre
рекомбиназы
, что якобы позволяет сочетать высокую эффективность вирусной доставки с достоинствами перепрограммированных клеток, свободных от остатков трансгенов, которые могут вызывать злокачественную трансформацию.
Доставка c помощью аденоассоциированного вирусного вектора в экзосоме
Аденоассоциированный вирусный
вектор (AAV) может быть связан с
экзосомами
(exo-AAV), если вектор выделяют из культуральной среды клеток-продуцентов. Такой вектор более устойчив к нейтрализующим антителам по сравнению со стандартным AAV. Он более эффективен для трансфекции in vivo
Невирусная доставка векторов
По сравнению с вирусными векторами, не-вирусные векторы являются потенциально менее иммуногенными и их сравнительно легче использовать в условиях клиники.
Невирусным подходом является прямая доставка в клетку
синтетической мРНК
(-) кодирующей четыре канонических фактора Яманаки: KLF4, c-MYC, OCT4, и SOX2. Метод позволяет достичь высокой эффективности перепрограммирования, но технически сложен и сильно зависит от качества реактивов
. Недавно он был модифицирован,
что позволило сократить продолжительность процесса и число необходимых реагентов. Ещё более экономным в отношении стоимости перепрограммирования является метод получения ИПСК и последующей их дифференцировки с помощью микрожидкостного устройства в объёмах не превышающих микролитра и эффективностью в 50 раз большей чем при традиционном перепрограммировании путём доставки синтетических мРНК, кодирующих факторы транскрипции
.
Для перепрограммирования in vivo очевидно подойдёт доставка мРНК с помощью олиго (карбонат-b-α-амино эфиров) под общим названием CARTs (charge-altering releasable transporters), катионов, которые образуя комплекс с мРНК, защищают её и доставив в клетку высвобождают
Перепрограммирование мышечных клеток головастиков Xenopus в недифференцированные клетки может быть достигнуто In vivo с помощью
ДНК
(±) мыши кодирующей Oct4, Sox2, и Klf4 в условиях способствующих регенерации.
Привлекательным методом невирусной доставки генов, который позволяет эффективно встраивать желаемую ДНК в геном различных клеток является использование
транспозонов
— сложных структур содержащих дискретные куски ДНК, обладающие способностью изменять своё местоположение в геноме посредством механизма транспозиции (инсерции), вынуждающего его включиться или наоборот покинуть определённый участок генома. Транспозон состоит из инсерционных сегментов ДНК, которые могут перемещаться как единое целое, захватывая лежащие между ними гены. Описано несколько транспозонных систем пригодных для транспортировки генов в клетки млекопитающих. Это «Спящая красавица» — Sleeping Beauty (SB), SB100X, и Tol2 и PiggyBac (PB). Разработаны эффективные методики получения ИПСК мыши и человека введением в соматические клетки векторов на основе
PiggyBac
(тр)
В том числе с кассетой из ДНК кодирующей 6 факторов (Oct4, c-Myc, Klf4, Sox2 плюс
и
).
, что позволило сократить продолжительность перепрограммирования до 4-12 дней и поднять его эффективность до уровня сопоставимого с методами переноса ядер и слияния клеток.
Векторные системы на основе
эписомных
плазмид
(±). Перепрограммирование основанное на использовании эписомных плазмид считается наиболее эффективным и безопасным так как не требует интеграции
трансгенов
в геном
. Однако первоначально эффективность перепрограммирования этим методом была крайне низка (менее 0,0002 %.) . Значительно увеличить эффективность индукции ИПСК позволило использование комбинации плазмид, кодирующих OCT3/4, SOX2, KLF4, L-MYC, LIN28 и малой интерферирующей РНК для ингибирования гена TP53, кодирующего белок
p53
(супрессор опухолей, ингибирующий перепрограммирование) в сочетании с
Эпштейн-Барр ядерным антигеном 1
(EBNA1), который является белком
вируса Эпштейна-Барр
(ВЭБ), необходимым для амплификации эписомных векторов
Разработаны полицистронные свободные от вирусов плазмиды (±), содержащие последовательности самовыщепляющегося 2A пептида (пептида, который взаимодействуя с рибосомой инициирует отделение от растущего полипротеина, вызвав «пропуск» последней пептидной связи на С-конце 2А
) и постоянно активный промотор вируса энцефаломиокардита (CMV).
Рекомбинантные белки
(-) представляют собой проникающие в ядро белки, полученные путём
рекомбинации
(в данном случае, присоединения последовательности, кодирующей поли-аргининовый домен
трансдукции
, к генам четырёх перепрограммирующих факторов: Oct4, Sox2, Klf4 и C-Myc, на участке соответствующем их С-концевой последовательности, и, кроме того, постоянно активного промотора вируса энцефаломиокардита) с последующим синтезом этих белков в тельцах включения бактерии
E.coli
. Выделенные из E. coli и очищенные рекомбинантные белки используются для репрограммирования без интеграции. Эффективность репрограммирования с помощью рекомбинантных белков ничтожно мала, но может быть существенно увеличена при воспалении вызываемом поли I:C (синтетическим аналогом двухцепочечной РНК)
.
Химически индуцированные плюрипотентные клетки (ХИПСК)
Используя для перепрограммирования исключительно малые молекулы, китайские учёные Дэн Хонгкуй с коллегами показали, что для перепрограммирования клеток достаточно эндогенных «мастер генов». Они индуцировали плюрипотентное состояние у взрослых клеток мыши, используя семь низкомолекулярных соединений
.
Эффективность метода оказалась достаточно высокой: она была в состоянии преобразовать 0,2 % взрослых клеток ткани в ИПСК, что сопоставимо с результатами получаемыми при использовании экзогенных «мастер генов» доставляемых вирусными векторами.
Авторы отмечают, что полученные от мышей CiPSCs были на «100 % жизнеспособными и здоровы на протяжении, по меньшей мере, 6 месяцев наблюдения».
В число этих семи низкомолекулярных соединений входили:
ингибитор GSK3 — гликоген синтез киназы 3 (CHIR) (подробнее см.
). Ингибирование GSK3 имитирует активацию
сигнального пути Wnt
в результате чего
β-катенин
избежав деградации входит в ядро, где активирует синтез ферментативной субъединицы
теломеразы
(TERT)
;
ингибитор
трансформирующего ростового фактора бета
(E-616452). Низкомолекулярный ингибитор TGF-[бета] сигнализации в ходе перепрограммирования заменяет Sox2 активируя Nanog
;
ингибитор
моноаминоксидазы
антидепрессант
транилципромин
одновременно является ещё и ингибитором деметилазы которая избирательно удаляет
метильные группы
с
лизина
в положении Лиз4 или Лиз9
гистона H3
. Действуя на хроматин транилципромин вызывает глобальное увеличение H3K4 метилирования
и как следствие этого дерепрессию генов
. Ингибиторы деметилаз обладают ещё и способностью подавлять избыточную
пролиферацию
клеток и таким образом подавлять развитие раковых опухолей
ингибитор гидролазы S-аденозилгомоцистеина (под названием 3-деазанепланоцин (DZNep)), вещество вызывающее разрушение
поликомб репрессорного комплекса PRC2
и ингибирующее метилирование гистонов
.
PD-0325901 — высокоизбирательный и сильнодействующий синтетический ингибитор
митоген-активируемой протеинкиназы MEK
, избирательно фосфорилирующей серин / треонин и остатки тирозина в активационной петле её субстратов. PD-0325901 проходит испытания в качестве противоопухолевого препарата.
Методы и составы «коктейлей» из малых молекул для химического воздействия на мышечную ткань с целью активации процессов образования функциональных сердечных, скелетных и гладкомышечных клеток и регенерации повреждённых тканей in situ можно найти в обзоре Джанга и Виллиамса.
С механизмами действия малых молекул при перепрограммировании можно ознакомиться в обзорах
.
Прямое перепрограммирование фибробластов через универсальный XEN этап
В 2015-м году система химического перепрограммирования была усовершенствована — разбита на три этапа с использованием разных «коктейлей» на разных этапах. Были найдены новые малые молекулы. Это позволило поднять выход перепрограммированных клеток почти в 1000 раз
.
Выяснилось, что поскольку начальный этап химического перепрограммирования переводит клетки фибробластов в стабильное XEN (extra-embryonic endoderm)-подобное состояние одинаковое для прямого перепрограммирование фибробластов в различные клетки, эти XEN-подобные клетки (которые можно размножать) могут служить универсальной платформой для создания различных желаемых типов клеток
.
Обнаружено что основным барьером для химического перепрограммирования является путь
, ингибирование которого необходимо для подавления провоспалительных путей мешающих индукции клеточной пластичности и программы, подобной
регенерации
конечностей
аксолотля
. Не случайно поэтому эволюционно консервативный сигнальный
путь N-концевой киназы c-Jun (JNK)
является важным генетическим детерминантом контроля
долголетия
Краситель для выявления плюрипотентных клеток человека
Для безопасной терапии индуцированными стволовыми клетками необходимы несложные методы обнаружения и разрушения недифференцированных стволовых клеток. С этой целью широко используются антитела SSEA-4 и SSEA-5, а также антитела на антигены: антитело TRA-1-60
на трансмембранный гликопротеин подокаликсин (
), Oct3/Oct4 и Nanog. Эти вещества как и большинство белков стоят недёшево и быстро портятся. Гораздо более стабильным и дешёвым реагентом способным отличить плюрипотентные стволовые клетки от дифференцированных клеток оказался флуоресцентный краситель KP-1 (Kyoto probe 1), который избирательно красит в митохондриях альдегиддегидрогеназу 2 (ALDH2). Эта селективность KP-1 зависит от способности митохондрии удалить его с помощью транспортных белков множественной лекарственной устойчивости ABCB1 и ABCG2, экспрессия которых подавляется в плюрипотентных клетках человека и индуцируется после дифференцировки
.
Фотодинамическая технология удаления ИПСК
Способность плюрипотентных клеток, и в частности ИПСК, избирательно окрашиваться красным красителем CD1 может быть использована для их селективного удаления из инкубационной среды, с помощью фотодинамической обработки светом, оставшихся недифференцированными ИПСК после проведения их дифференцировки в соматические клетки. Эта простая технология предварительной очистки позволяет резко снизить риск образования опухолей типа тератомы при трансплантации клеток полученных из ИПСК
.
Разработана также разновидность этой технологии. Для этого флуоресцентную пробу KP-1 (Kyoto probe 1) (которая обычными клетками выводится наружу белками ABCB1 и ABCG2, синтез которых в ИПСК подавлен) соединили с анти раковым препаратом SN38. Полученный препарат, названный «конъюгат 17» за 72 часа полностью удаляет оставшиеся недифференцированными ИПСК
Химическая технология удаления ИПСК
in vitro
Чтобы избавиться от плюрипотентных клеток, которые могут вызвать образование терратомы, можно использовать синтетические фосфо-D-пептиды, поскольку известно что на поверхности ИПСК сверхэкспрессируются щелочные фосфатазы, которые вызывают дефосфорилирование фосфо-D-пептидов в гидрофобные пептиды, которые агрегируя вызывают гибель клеток
.
Можно также удалить плюрипотентные клетки с помощью препарата Брекинар (
, DuP-785), который действует как мощный и селективный ингибитор фермента
. Он блокирует синтез нуклеотидов на основе пиримидина в организме и таким образом подавляет рост клеток. В опытах с плюрипотентными стволовыми клетками мыши выяснилось что брекинар вызывает остановку клеточного цикла, гибель именно стволовых клеток, тогда как в отношении нормальных тканеспецифичных стволовых клеток и дифференцирующихся клеток он менее токсичен.
Описаны также способы устранения ИПСК, смешанных с кардиомиоцитами, нейронами и гепатоцитами, с использованием препарата от ожирения, называемого
орлистат
или аторвастатина
.
Перспективы изучения индуцированных стволовых клеток для медицины
Ярким свидетельством значения индуцированных стволовых клеток для медицины будущего и всего человечества, стало присуждение
Джону Гордону
и
Синъе Яманаке
Нобелевской премии 2012 года по медицине
. Бо́льшую часть от Нобелевской премии, а также от полученной им в 2013 году
Премии за прорыв в области медицины
в 3 млн долларов Синъя Яманака решил потратить на развитие своих исследований.
В настоящее время индуцированные стволовые клетки используются главным образом для моделирования болезней, скрининга (селективного отбора) лекарств, проверки токсичности различных препаратов. Однако в ближайшие годы начнётся их широкомасштабное использование для клеточной терапии и
выращивания органов и их «запчастей»
для трансплантации
.
↑
Tachibana M., Amato P., Sparman M., Gutierrez N. M., Tippner-Hedges R., Ma H., Kang E., Fulati A., Lee H. S., Sritanaudomchai H., Masterson K., Larson J., Eaton D., Sadler-Fredd K., Battaglia D., Lee D., Wu D., Jensen J., Patton P., Gokhale S., Stouffer R. L., Wolf D., Mitalipov S.
(англ.)
// Cell. — 2013. — Vol. 153,
no. 6
. — P. 1228—1238. —
doi
:
. —
.
[
]
↑
Takahashi K., Yamanaka S.
(англ.)
// Cell. — 2006. — Vol. 126,
no. 4
. — P. 663—676. —
doi
:
. —
.
[
]
↑
Yoshioka N., Gros E., Li H. R., Kumar S., Deacon D. C., Maron C., Muotri A. R., Chi N. C., Fu X. D., Yu B. D., Dowdy S. F.
(англ.)
// Cell stem cell. — 2013. — Vol. 13,
no. 2
. — P. 246—254. —
doi
:
. —
.
[
]
↑
Hou P., Li Y., Zhang X., Liu C., Guan J., Li H., Zhao T., Ye J., Yang W., Liu K., Ge J., Xu J., Zhang Q., Zhao Y., Deng H.
(англ.)
// Science (New York, N.Y.). — 2013. — Vol. 341,
no. 6146
. — P. 651—654. —
doi
:
. —
.
[
]
Lin J., Li M. R., Ti D. D., Chen M. X., Hao H. J., Zhao Y. L., Fu X. B., Han W. D.
(англ.)
// Ageing research reviews. — 2013. — Vol. 12,
no. 1
. — P. 29—38. —
doi
:
. —
.
[
]
Yamanaka S., Blau H. M.
(англ.)
// Nature. — 2010. — Vol. 465,
no. 7299
. — P. 704—712. —
doi
:
. —
.
[
]
Gurdon J. B., Wilmut I.
(англ.)
// Cold Spring Harbor perspectives in biology. — 2011. — Vol. 3,
no. 6
. —
doi
:
. —
.
[
]
Do J. T., Han D. W., Gentile L., Sobek-Klocke I., Stehling M., Lee H. T., Schöler H. R.
(англ.)
// Stem cells (Dayton, Ohio). — 2007. — Vol. 25,
no. 4
. — P. 1013—1020. —
doi
:
. —
.
[
]
Takahashi K., Tanabe K., Ohnuki M., Narita M., Ichisaka T., Tomoda K., Yamanaka S.
(англ.)
// Cell. — 2007. — Vol. 131,
no. 5
. — P. 861—872. —
doi
:
. —
.
[
]
↑
Wang W., Yang J., Liu H., Lu D., Chen X., Zenonos Z., Campos L. S., Rad R., Guo G., Zhang S., Bradley A., Liu P.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 2011. — Vol. 108,
no. 45
. — P. 18283—18288. —
doi
:
. —
.
[
]
↑
Lapasset L., Milhavet O., Prieur A., Besnard E., Babled A., Aït-Hamou N., Leschik J., Pellestor F., Ramirez J. M., De Vos J., Lehmann S., Lemaitre J. M.
(англ.)
// Genes & development. — 2011. — Vol. 25,
no. 21
. — P. 2248—2253. —
doi
:
. —
.
[
]
↑
Zhou H., Wu S., Joo J. Y., Zhu S., Han D. W., Lin T., Trauger S., Bien G., Yao S., Zhu Y., Siuzdak G., Schöler H. R., Duan L., Ding S.
(англ.)
// Cell stem cell. — 2009. — Vol. 4,
no. 5
. — P. 381—384. —
doi
:
. —
.
[
]
↑
Lee J., Sayed N., Hunter A., Au K. F., Wong W. H., Mocarski E. S., Pera R. R., Yakubov E., Cooke J. P.
(англ.)
// Cell. — 2012. — Vol. 151,
no. 3
. — P. 547—558. —
doi
:
. —
.
[
]
Li Z., Rana T. M.
(англ.)
// Current protocols in stem cell biology. — 2012. — Vol. Chapter 4. — P. 4. —
doi
:
. —
.
[
]
Anokye-Danso F., Trivedi C. M., Juhr D., Gupta M., Cui Z., Tian Y., Zhang Y., Yang W., Gruber P. J., Epstein J. A., Morrisey E. E.
(англ.)
// Cell stem cell. — 2011. — Vol. 8,
no. 4
. — P. 376—388. —
doi
:
. —
.
[
]
Miyoshi N., Ishii H., Nagano H., Haraguchi N., Dewi D. L., Kano Y., Nishikawa S., Tanemura M., Mimori K., Tanaka F., Saito T., Nishimura J., Takemasa I., Mizushima T., Ikeda M., Yamamoto H., Sekimoto M., Doki Y., Mori M.
(англ.)
// Cell stem cell. — 2011. — Vol. 8,
no. 6
. — P. 633—638. —
doi
:
. —
.
[
]
↑
Jayawardena T. M., Egemnazarov B., Finch E. A., Zhang L., Payne J. A., Pandya K., Zhang Z., Rosenberg P., Mirotsou M., Dzau V. J.
(англ.)
// Circulation research. — 2012. — Vol. 110,
no. 11
. — P. 1465—1473. —
doi
:
. —
.
[
]
Bao X., Zhu X., Liao B., Benda C., Zhuang Q., Pei D., Qin B., Esteban M. A.
(англ.)
// Current opinion in cell biology. — 2013. — Vol. 25,
no. 2
. — P. 208—214. —
doi
:
. —
.
[
]
Efe J. A., Ding S.
(англ.)
// Philosophical transactions of the Royal Society of London. Series B, Biological sciences. — 2011. — Vol. 366,
no. 1575
. — P. 2208—2221. —
doi
:
. —
.
[
]
Ladewig J., Mertens J., Kesavan J., Doerr J., Poppe D., Glaue F., Herms S., Wernet P., Kögler G., Müller F. J., Koch P., Brüstle O.
(англ.)
// Nature methods. — 2012. — Vol. 9,
no. 6
. — P. 575—578. —
doi
:
. —
.
[
]
Moschidou D., Mukherjee S., Blundell M. P., Drews K., Jones G. N., Abdulrazzak H., Nowakowska B., Phoolchund A., Lay K., Ramasamy T. S., Cananzi M., Nettersheim D., Sullivan M., Frost J., Moore G., Vermeesch J. R., Fisk N. M., Thrasher A. J., Atala A., Adjaye J., Schorle H., De Coppi P., Guillot P. V.
(англ.)
// Molecular therapy : the journal of the American Society of Gene Therapy. — 2012. — Vol. 20,
no. 10
. — P. 1953—1967. —
doi
:
. —
.
[
]
Pandian G. N., Sugiyama H.
(англ.)
// Biotechnology journal. — 2012. — Vol. 7,
no. 6
. — P. 798—809. —
doi
:
. —
.
[
]
Pandian G. N., Nakano Y., Sato S., Morinaga H., Bando T., Nagase H., Sugiyama H.
(англ.)
// Scientific reports. — 2012. — Vol. 2. — P. 544. —
doi
:
. —
.
[
]
от 19 августа 2014 на
Wayback Machine
FROM THE ARTICLE:
De Robertis E. M.
(англ.)
// Nature reviews. Molecular cell biology. — 2006. — Vol. 7,
no. 4
. — P. 296—302. —
doi
:
. —
.
[
]
Slack J. M.
(англ.)
// The Journal of pathology. — 2009. — Vol. 217,
no. 2
. — P. 161—168. —
doi
:
. —
.
[
]
Wei G., Schubiger G., Harder F., Müller A. M.
(англ.)
// Stem cells (Dayton, Ohio). — 2000. — Vol. 18,
no. 6
. — P. 409—414. —
doi
:
. —
.
[
]
Worley M. I., Setiawan L., Hariharan I. K.
(англ.)
// Annual review of genetics. — 2012. — Vol. 46. — P. 289—310. —
doi
:
. —
.
[
]
Xu P. F., Houssin N., Ferri-Lagneau K. F., Thisse B., Thisse C.
(англ.)
// Science (New York, N.Y.). — 2014. — Vol. 344,
no. 6179
. — P. 87—89. —
doi
:
. —
.
[
]
Yan K. S. et al.,& Kuo C. J. (2017).
. Cell Stem Cell, 21(1), 78-90.e6 DOI:
Stange D. E., Koo B. K., Huch M., Sibbel G., Basak O., Lyubimova A., Kujala P., Bartfeld S., Koster J., Geahlen J. H., Peters P. J., van Es J. H., van de Wetering M., Mills J. C., Clevers H.
(англ.)
// Cell. — 2013. — Vol. 155,
no. 2
. — P. 357—368. —
doi
:
. —
.
[
]
Tata P. R., Mou H., Pardo-Saganta A., Zhao R., Prabhu M., Law B. M., Vinarsky V., Cho J. L., Breton S., Sahay A., Medoff B. D., Rajagopal J.
(англ.)
// Nature. — 2013. — Vol. 503,
no. 7475
. — P. 218—223. —
doi
:
. —
.
[
]
Kusaba T., Lalli M., Kramann R., Kobayashi A., Humphreys B. D.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 2014. — Vol. 111,
no. 4
. — P. 1527—1532. —
doi
:
. —
.
[
]
Sieweke M. H., Allen J. E.
(англ.)
// Science (New York, N.Y.). — 2013. — Vol. 342,
no. 6161
. — P. 1242974. —
doi
:
. —
.
[
]
Soucie E. L., Weng Z., Geirsdóttir L., Molawi K., Maurizio J., Fenouil R., Mossadegh-Keller N., Gimenez G., VanHille L., Beniazza M., Favret J., Berruyer C., Perrin P., Hacohen N., Andrau J. C., Ferrier P., Dubreuil P., Sidow A., Sieweke M. H.
(англ.)
// Science (New York, N.Y.). — 2016. — Vol. 351,
no. 6274
. — P. 5510. —
doi
:
. —
.
[
]
Sandoval-Guzmán T., Wang H., Khattak S., Schuez M., Roensch K., Nacu E., Tazaki A., Joven A., Tanaka E. M., Simon A.
(англ.)
// Cell stem cell. — 2014. — Vol. 14,
no. 2
. — P. 174—187. —
doi
:
. —
.
[
]
Kuroda Y., Wakao S., Kitada M., Murakami T., Nojima M., Dezawa M.
(англ.)
// Nature protocols. — 2013. — Vol. 8,
no. 7
. — P. 1391—1415. —
doi
:
. —
.
[
]
Kuroda Y., Kitada M., Wakao S., Nishikawa K., Tanimura Y., Makinoshima H., Goda M., Akashi H., Inutsuka A., Niwa A., Shigemoto T., Nabeshima Y., Nakahata T., Nabeshima Y., Fujiyoshi Y., Dezawa M.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 2010. — Vol. 107,
no. 19
. — P. 8639—8643. —
doi
:
. —
.
[
]
Ogura F., Wakao S., Kuroda Y., Tsuchiyama K., Bagheri M., Heneidi S., Chazenbalk G., Aiba S., Dezawa M.
(англ.)
// Stem cells and development. — 2014. — Vol. 23,
no. 7
. — P. 717—728. —
doi
:
. —
.
[
]
Heneidi S., Simerman A. A., Keller E., Singh P., Li X., Dumesic D. A., Chazenbalk G.
(англ.)
// Public Library of Science ONE. — 2013. — Vol. 8,
no. 6
. — P. e64752. —
doi
:
. —
.
[
]
Simerman A. A., Dumesic D. A., Chazenbalk G. D.
(англ.)
// Clinical and translational medicine. — 2014. — Vol. 3. — P. 12. —
doi
:
. —
.
[
]
Wakao S., Kitada M., Dezawa M.
(англ.)
// Cytometry. Part A : the journal of the International Society for Analytical Cytology. — 2013. — Vol. 83,
no. 1
. — P. 18—26. —
doi
:
. —
.
[
]
Wakao S., Kitada M., Kuroda Y., Shigemoto T., Matsuse D., Akashi H., Tanimura Y., Tsuchiyama K., Kikuchi T., Goda M., Nakahata T., Fujiyoshi Y., Dezawa M.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 2011. — Vol. 108,
no. 24
. — P. 9875—9880. —
doi
:
. —
.
[
]
Sisakhtnezhad S., Matin M. M.
(англ.)
// Cell and tissue research. — 2012. — Vol. 348,
no. 3
. — P. 379—396. —
doi
:
. —
.
[
]
Merrell A. J., Stanger B. Z.
(англ.)
// Nature reviews. Molecular cell biology. — 2016. — Vol. 17,
no. 7
. — P. 413—425. —
doi
:
. —
.
[
]
Tokuzawa Y., Kaiho E., Maruyama M., Takahashi K., Mitsui K., Maeda M., Niwa H., Yamanaka S.
(англ.)
// Molecular and cellular biology. — 2003. — Vol. 23,
no. 8
. — P. 2699—2708. —
.
[
]
Andras Dinnyes, Xiuchun Cindy Tian, and Bj¨orn Oback.
Nuclear Transfer for Cloning Animals // Stem Cells. — Wiley-Blackwell, 2013. — P. 299—344. —
ISBN 978-3-527-32925-0
.
Ogura A., Inoue K., Wakayama T.
(англ.)
// Philosophical transactions of the Royal Society of London. Series B, Biological sciences. — 2013. — Vol. 368,
no. 1609
. — P. 20110329. —
doi
:
. —
.
[
]
Wilmut I., Schnieke A. E., McWhir J., Kind A. J., Campbell K. H.
(англ.)
// Nature. — 1997. — Vol. 385,
no. 6619
. — P. 810—813. —
doi
:
. —
.
[
]
Jullien J., Pasque V., Halley-Stott R. P., Miyamoto K., Gurdon J. B.
(англ.)
// Nature reviews. Molecular cell biology. — 2011. — Vol. 12,
no. 7
. — P. 453—459. —
doi
:
. —
.
[
]
Campbell K. H.
(англ.)
// Journal of anatomy. — 2002. — Vol. 200,
no. Pt 3
. — P. 267—275. —
.
[
]
Pan G., Wang T., Yao H., Pei D.
(англ.)
// BioEssays : news and reviews in molecular, cellular and developmental biology. — 2012. — Vol. 34,
no. 6
. — P. 472—476. —
doi
:
. —
.
[
]
Roh, et al. (Feb., 2014). Human embryonic stem cell line prepared by nuclear transfer of a human somatic cell into an enucleated human oocyte. United States Patent № 8,647,872
Hosseini S. M., Hajian M., Forouzanfar M., Moulavi F., Abedi P., Asgari V., Tanhaei S., Abbasi H., Jafarpour F., Ostadhosseini S., Karamali F., Karbaliaie K., Baharvand H., Nasr-Esfahani M. H.
(англ.)
// Cellular reprogramming. — 2012. — Vol. 14,
no. 2
. — P. 155—163. —
doi
:
. —
.
[
]
↑
Gupta M. K., Das Z. C., Heo Y. T., Joo J. Y., Chung H. J., Song H., Kim J. H., Kim N. H., Lee H. T., Ko D. H., Uhm S. J.
(англ.)
// Cellular reprogramming. — 2013. — Vol. 15,
no. 4
. — P. 322—328. —
doi
:
. —
.
[
]
De Bem T. H., Chiaratti M. R., Rochetti R., Bressan F. F., Sangalli J. R., Miranda M. S., Pires P. R., Schwartz K. R., Sampaio R. V., Fantinato-Neto P., Pimentel J. R., Perecin F., Smith L. C., Meirelles F. V., Adona P. R., Leal C. L.
(англ.)
// Cellular reprogramming. — 2011. — Vol. 13,
no. 5
. — P. 419—429. —
doi
:
. —
.
[
]
↑
Kishigami S., Mizutani E., Ohta H., Hikichi T., Thuan N. V., Wakayama S., Bui H. T., Wakayama T.
(англ.)
// Biochemical and biophysical research communications. — 2006. — Vol. 340,
no. 1
. — P. 183—189. —
doi
:
. —
.
[
]
Terashita Y., Wakayama S., Yamagata K., Li C., Sato E., Wakayama T.
(англ.)
// Biology of reproduction. — 2012. — Vol. 86,
no. 6
. — P. 180. —
doi
:
. —
.
[
]
Eva Hörmanseder, Angela Simeone, George E. Allen, Charles R. Bradshaw, Magdalena Figlmüller, John Gurdon, Jerome Jullien (2017).
. Cell Stem Cell,
doi
:
Qu P, Qing S, Liu R, Qin H, Wang W, Qiao F, et al. (2017) Effects of embryo-derived exosomes on the development of bovine cloned embryos. PLoS ONE12(3): e0174535.
Wakayama S., Kohda T., Obokata H., Tokoro M., Li C., Terashita Y., Mizutani E., Nguyen V. T., Kishigami S., Ishino F., Wakayama T.
(англ.)
// Cell stem cell. — 2013. — Vol. 12,
no. 3
. — P. 293—297. —
doi
:
. —
.
[
]
Kong Q., Ji G., Xie B., Li J., Mao J., Wang J., Liu S., Liu L., Liu Z.
(англ.)
// Stem cell reviews. — 2014. — Vol. 10,
no. 3
. — P. 399—407. —
doi
:
. —
.
[
]
Chung Y. G., Eum J. H., Lee J. E., Shim S. H., Sepilian V., Hong S. W., Lee Y., Treff N. R., Choi Y. H., Kimbrel E. A., Dittman R. E., Lanza R., Lee D. R.
(англ.)
// Cell stem cell. — 2014. — Vol. 14,
no. 6
. — P. 777—780. —
doi
:
. —
.
[
]
Monya Baker (April 2014)
от 29 апреля 2014 на
Wayback Machine
. Nature
Yang H., Shi L., Wang B. A., Liang D., Zhong C., Liu W., Nie Y., Liu J., Zhao J., Gao X., Li D., Xu G. L., Li J.
(англ.)
// Cell. — 2012. — Vol. 149,
no. 3
. — P. 605—617. —
doi
:
. —
.
[
]
Hayashi K., Ogushi S., Kurimoto K., Shimamoto S., Ohta H., Saitou M.
(англ.)
// Science (New York, N.Y.). — 2012. — Vol. 338,
no. 6109
. — P. 971—975. —
doi
:
. —
.
[
]
Tachibana M., Sparman M., Mitalipov S.
(англ.)
// Fertility and sterility. — 2012. — Vol. 97,
no. 5
. — P. e16. —
doi
:
. —
.
[
]
Paull D., Emmanuele V., Weiss K. A., Treff N., Stewart L., Hua H., Zimmer M., Kahler D. J., Goland R. S., Noggle S. A., Prosser R., Hirano M., Sauer M. V., Egli D.
(англ.)
// Nature. — 2013. — Vol. 493,
no. 7434
. — P. 632—637. —
doi
:
. —
.
[
]
Tachibana M., Amato P., Sparman M., Woodward J., Sanchis D. M., Ma H., Gutierrez N. M., Tippner-Hedges R., Kang E., Lee H. S., Ramsey C., Masterson K., Battaglia D., Lee D., Wu D., Jensen J., Patton P., Gokhale S., Stouffer R., Mitalipov S.
(англ.)
// Nature. — 2013. — Vol. 493,
no. 7434
. — P. 627—631. —
doi
:
. —
.
[
]
Hayden E. C.
(англ.)
// Nature. — 2013. — Vol. 502,
no. 7471
. — P. 284—285. —
doi
:
. —
.
[
]
Wang, X., Qu, J., Li, J., He, H., Liu, Z., & Huan, Y. (2020). Epigenetic Reprogramming During Somatic Cell Nuclear Transfer: Recent Progress and Future Directions. Frontiers in Genetics, 11, 205.
doi
:
PMC
Malin, K., Witkowska-Piłaszewicz, O., & Papis, K. (2022). The many problems of somatic cell nuclear transfer in reproductive cloning of mammals. Theriogenology, 189, 246-254.
PMID
doi
:
↑
Abad M., Mosteiro L., Pantoja C., Cañamero M., Rayon T., Ors I., Graña O., Megías D., Domínguez O., Martínez D., Manzanares M., Ortega S., Serrano M.
(англ.)
// Nature. — 2013. — Vol. 502,
no. 7471
. — P. 340—345. —
doi
:
. —
.
[
]
Shinagawa T., Takagi T., Tsukamoto D., Tomaru C., Huynh L. M., Sivaraman P., Kumarevel T., Inoue K., Nakato R., Katou Y., Sado T., Takahashi S., Ogura A., Shirahige K., Ishii S.
(англ.)
// Cell stem cell. — 2014. — Vol. 14,
no. 2
. — P. 217—227. —
doi
:
. —
.
[
]
Ishiuchi T., Enriquez-Gasca R., Mizutani E., Bošković A., Ziegler-Birling C., Rodriguez-Terrones D., Wakayama T., Vaquerizas J. M., Torres-Padilla M. E.
(англ.)
// Nature structural & molecular biology. — 2015. — Vol. 22,
no. 9
. — P. 662—671. —
doi
:
. —
.
[
]
Yong Jin Choi, Chao-Po Lin, Davide Risso, et al., & Lin He (2017).
от 10 ноября 2018 на
Wayback Machine
. Science :
doi
:
Vanderburg B.B. (2017).
от 16 января 2017 на
Wayback Machine
. ReliaWire
Hao, C., Chu, S., Quan, X., Zhou, T., Shi, J., Huang, X., ... & Pei, D. (2023). Establishing extended pluripotent stem cells from human urine cells. Cell & Bioscience, 13(1), 1-11.
PMID
PMC
doi
:
Hu, Y., Yang, Y., Tan, P. et al. (2022). Induction of mouse totipotent stem cells by a defined chemical cocktail. Nature
doi
:
Niu Z., Hu Y., Chu Z., Yu M., Bai Y., Wang L., Hua J.
(англ.)
// Cell biochemistry and function. — 2013. — Vol. 31,
no. 1
. — P. 12—19. —
doi
:
. —
.
[
]
Panula S., Medrano J. V., Kee K., Bergström R., Nguyen H. N., Byers B., Wilson K. D., Wu J. C., Simon C., Hovatta O., Reijo Pera R. A.
(англ.)
// Human molecular genetics. — 2011. — Vol. 20,
no. 4
. — P. 752—762. —
doi
:
. —
.
[
]
Yang S., Bo J., Hu H., Guo X., Tian R., Sun C., Zhu Y., Li P., Liu P., Zou S., Huang Y., Li Z.
(англ.)
// Cell proliferation. — 2012. — Vol. 45,
no. 2
. — P. 91—100. —
doi
:
. —
.
[
]
Irie N., Weinberger L., Tang W. W., Kobayashi T., Viukov S., Manor Y. S., Dietmann S., Hanna J. H., Surani M. A.
(англ.)
// Cell. — 2015. — Vol. 160,
no. 1-2
. — P. 253—268. —
doi
:
. —
.
[
]
Zhou, Q., Wang, M., Yuan, Y., Wang, X., Fu, R., Wan, H., … & Feng, G. (2016).
от 29 января 2017 на
Wayback Machine
. Cell stem cell, 18(3), 330—340. DOI:
Zhang, Y., & Liu, Y. (2016).
от 4 июня 2018 на
Wayback Machine
. Science China Life Sciences, 59(4), 436—437.
doi
:
PMID
Hou, J., Yang, S., Yang, H., Liu, Y., Liu, Y., Hai, Y., … & Li, Z. (2014).
от 5 июля 2017 на
Wayback Machine
. Reproduction, 147(6), R179-R188.
doi
:
Naoko Irie, Shinseog Kim, and M. Azim Surani (2016).
. Journal of Mammalian Ova Research, 33(2), 79-87
doi
:
Xu, H., Yang, M., Tian, R. et al. (2020). Derivation and propagation of spermatogonial stem cells from human pluripotent cells. Stem Cell Res Ther 11, 408
Orie Hikabe, Nobuhiko Hamazaki, Go Nagamatsu, Yayoi Obata, Yuji Hirao, Norio Hamada, So Shimamoto, Takuya Imamura, Kinichi Nakashima, Mitinori Saitou & Katsuhiko Hayashi (2016).
. Nature,
doi
:
Yamashiro, C., Sasaki, K., Yokobayashi, S. et al. (2020). Generation of human oogonia from induced pluripotent stem cells in culture. Nat Protoc 15, 1560—1583
Hamazaki, N., Kyogoku, H., Araki, H. et al. (2020). Reconstitution of the oocyte transcriptional network with transcription factors. Nature 589(7841):264-269
PMID
doi
:
Stevens L. C.
(англ.)
// Developmental biology. — 1970. — Vol. 21,
no. 3
. — P. 364—382. —
.
[
]
Mintz B., Cronmiller C., Custer R. P.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 1978. — Vol. 75,
no. 6
. — P. 2834—2838. —
.
[
]
Mintz B., Illmensee K.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 1975. — Vol. 72,
no. 9
. — P. 3585—3589. —
.
[
]
Martin G. R., Evans M. J.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 1975. — Vol. 72,
no. 4
. — P. 1441—1445. —
.
[
]
Illmensee K., Mintz B.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 1976. — Vol. 73,
no. 2
. — P. 549—553. —
.
[
]
Martin G. R.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 1981. — Vol. 78,
no. 12
. — P. 7634—7638. —
.
[
]
Альбертс Б. с соавтором И Дж. Уотсон (1987) Молекулярная биодогия клетки т.4 стр 72
↑
GRAHAM, C. F. (1977). Teratocarcinoma cells and normal mouse embryogenesis. In Concepts in Mammalian Embryogenesis (ed. M. I. Sherman), pp. 315—394. Cambridge: M.I.T. Press
↑
ILLMENSEE, K. (1978). Reversion of malignancy and normalized differentiation of teratocarcinoma cells in chimeric mice. In Genetic Mosaics and Chimeras in Mammals (ed. L. Russell), pp. 3-24. New York: Plenum
Martin G. R.
(англ.)
// Science (New York, N.Y.). — 1980. — Vol. 209,
no. 4458
. — P. 768—776. —
.
[
]
Papaioannou V. E., Gardner R. L., McBurney M. W., Babinet C., Evans M. J.
(англ.)
// Journal of embryology and experimental morphology. — 1978. — Vol. 44. — P. 93—104. —
.
[
]
Stewart C. L., Vanek M., Wagner E. F.
(англ.)
// The EMBO journal. — 1985. — Vol. 4,
no. 13B
. — P. 3701—3709. —
.
[
]
Jiho Choi, Soohyun Lee, William Mallard et al.,(2015).
. Nature Biotechnology,
doi
:
Phanstiel DH, Brumbaugh J, Wenger CD et al. & Coon JJ. (2011) Proteomic and phosphoproteomic comparison of human ES and iPS cells. Nat Methods.; 8(10): 821—827.
doi
:
↑
Linzhao Cheng, Nancy F. Hansen, Ling Zhao, et al & P. Paul Liu (2012) Low Incidence of DNA Sequence Variation in Human Induced Pluripotent Stem Cells Generated by Nonintegrating Plasmid Expression Cell Stem Cell, 2012; 10 (3), 337—344
doi
:
Zhao XY, Li W, Lv Z et al. (2009) iPS cells produce viable mice through tetraploid complementation. Nature ; 461: 86-90 doi:10.1038/nature08267
Choi, Jiho; Lee, Soohyun; Mallard, William; et al. (2015). A comparison of genetically matched cell lines reveals the equivalence of human iPSCs and ESCs. Nature Biotechnology 33: 1173—1181.
doi
:
Shutova, M. V., Surdina, A. V., Ischenko, D. S., Naumov, V. A., Bogomazova, A. N., Vassina, E. M., … & Kiselev, S. L. (2016). An integrative analysis of reprogramming in human isogenic system identified a clone selection criterion. Cell Cycle, 15(7), 986—997.
doi
:
↑
Kunitomi, A., Yuasa, S., Sugiyama, F., Saito, Y., Seki, T., Kusumoto, D., … & Egashira, T. (2016). H1foo Has a Pivotal Role in Qualifying Induced Pluripotent Stem Cells. Stem cell reports. 6(6), 825—833
doi
:
Nichols, J., Zevnik, B., Anastassiadis, K., Niwa, H., Klewe-Nebenius, D., Chambers, I., … & Smith, A. (1998). Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4. Cell, 95(3), 379—391.
Boyer, L. A., Lee, T. I., Cole, M. F., Johnstone, S. E., Levine, S. S., Zucker, J. P., … & Young, R. A. (2005). Core transcriptional regulatory circuitry in human embryonic stem cells. cell, 122(6), 947—956.
PMID
PMC
{{DOI: 10.1016/j.cell.2005.08.020}}
↑
Velychko, S., Adachi, K., Kim, K. P., Hou, Y., MacCarthy, C. M., Wu, G., & Schöler, H. R. (2019).
от 1 мая 2023 на
Wayback Machine
. Cell stem cell, 25(6), 737—753.
PMID
PMC
doi
:
от 8 ноября 2019 на
Wayback Machine
Oct4 is not only unnecessary but damaging during generation of mouse induced pluripotent stem cells (iPSCs)
An, Z., Liu, P., Zheng, J., Si, C., Li, T., Chen, Y., … & Ding, S. (2019).
. Cell reports, 29(7), 1986—2000.
PMID
doi
:
Yagi T, Kosakai A, Ito D, Okada Y, Akamatsu W, et al. (2012) Establishment of Induced Pluripotent Stem Cells from Centenarians for Neurodegenerative Disease Research. PLoS ONE 7(7): e41572
doi
:
Milhavet, O., & Lemaitre, J. M. (2014). Senescent-Derived Pluripotent Stem Cells Are Able to Redifferentiate into Fully Rejuvenated Cells. In Tumor Dormancy, Quiescence, and Senescence, Volume 2 (pp. 265—276). Springer Netherlands.
doi
:
Lee et al. (2020) Induced pluripotency and spontaneous reversal of cellular aging in supercentenarian donor cells. Biochemical and Biophysical Research Communications. DOI:
Yehezkel S, Rebibo-Sabbah A, Segev Y, Tzukerman M, Shaked R, Huber I, Gepstein L, Skorecki K, Selig S (2011) Reprogramming of telomeric regions during the generation of human induced pluripotent stem cells and subsequent differentiation into fibroblast-like derivatives. Epigenetics. 2011 Jan 1;6(1):63-75
West MD, Vaziri H. (2010) Back to immortality: the restoration of embryonic telomere length during induced pluripotency. Regen Med.;5(4):485-488
Gourronc FA, Klingelhutz AJ. (2011) Therapeutic opportunities: Telomere maintenance in inducible pluripotent stem cells. Mutat Res. 2011 May 13
Osamu Hashizume, Sakiko Ohnishi, Takayuki Mito, et al. & Jun-Ichi Hayashi (2015). Epigenetic regulation of the nuclear-coded GCAT and SHMT2 genes confers human age-associated mitochondrial respiration defects. Scientific Reports, 5, Article number: 10434
doi
:
Rohani, L., Johnson, A. A., Arnold, A. and Stolzing, A. (2014), The aging signature: a hallmark of induced pluripotent stem cells? Aging Cell, 13(1): 2-7.
doi
:
Shigeki Yagyu, Valentina Hoyos, Francesca Del Bufalo, Malcolm K Brenner. (2015). An Inducible Caspase-9 Suicide Gene to Improve the Safety of Therapy Using Human Induced Pluripotent Stem Cells. Molecular Therapy. 23, 1475-1485
Tongbiao Zhao,Zhen-Ning Zhang, Zhili Rong & Yang Xu (2011) Immunogenicity of induced pluripotent stem cells Nature 474, 212—215 doi:10.1038/nature10135
Dhodapkar MV, Dhodapkar KM.(2011) Spontaneous and therapy-induced immunity to pluripotency genes in humans: clinical implications, opportunities and challenges. Cancer Immunol Immunother.; 60(3):413-418
Ivan Gutierrez-Aranda, (2010) Human Induced Pluripotent Stem Cells Develop Teratoma More Efficiently and Faster than Human Embryonic Stem Cells Regardless of the Site of Injection. Stem Cells. 2010;28:1568-1570
Zhao T, Zhang ZN, Rong Z, Xu Y.(2011) Immunogenicity of induced pluripotent stem cells. Nature.;474:212-215
Paul J. Fairchild, Naoki Ichiryu (2013) Mitigating the Risk of Immunogenicity in the Pursuit of Induced Pluripotency. In «The Immunological Barriers to Regenerative Medicine» pp 77-94 Online
ISBN 978-1-4614-5480-9
Springer New York, DOI 10.1007/978-1-4614-5480-9_5
Jeremy I. Pearl, Joseph C. Wu (2013) The Immunogenicity of Embryonic Stem Cells and Their Differentiated Progeny. The Immunological Barriers to Regenerative Medicine Stem Cell Biology and Regenerative Medicine 2013, pp 37-48 Online
ISBN 978-1-4614-5480-9
Springer New York
Chan-Jung Chang, Koyel Mitra, Mariko Koya et al. & Eric E. Bouhassira (2011) Production of Embryonic and Fetal-Like Red Blood Cells from Human Induced Pluripotent Stem Cells. PLoS One.; 6(10): e25761. doi: 10.1371/journal.pone.0025761.
Lindgren AG, Natsuhara K, Tian E, Vincent JJ, Li X, et al. (2011) Loss of Pten Causes Tumor Initiation Following Differentiation of Murine Pluripotent Stem Cells Due to Failed Repression of Nanog. PLoS ONE
6(1): e16478. doi:10.1371/journal.pone.0016478
Grad, I., Hibaoui, Y., Jaconi,. et al. & Feki, A. (2011)
от 8 января 2014 на
Wayback Machine
. Eur. Cell Mater, 22, 258—274
Uri Ben-David, Qing-Fen Gan, Tamar Golan-Lev, et al & Nissim Benvenisty (2013)
от 24 сентября 2015 на
Wayback Machine
Cell Stem Cell, 12(2), 167—179
doi
:
Lou, K. J. (2013). Small molecules vs. teratomas. SciBX: Science-Business eXchange, 6(7). doi:10.1038/scibx.2013.158
Boheler, K. R., Bhattacharya, S., Kropp, E. M., Chuppa, S., Riordon, D. R., Bausch-Fluck, D., … & Gundry, R. L. (2014). A Human Pluripotent Stem Cell Surface N-Glycoproteome Resource Reveals Markers, Extracellular Epitopes, and Drug Targets. Stem Cell Reports.
doi
:
Chan, D. A., Sutphin, P. D., Nguyen, P., Turcotte, S., Lai, E. W., Banh, A., … & Giaccia, A. J. (2011). Targeting GLUT1 and the Warburg effect in renal cell carcinoma by chemical synthetic lethality. Science translational medicine, 3(94), 94ra70-94ra70.
Lee, M. O., Moon, S. H., Jeong, H. C. et al. and Cha, H. J. (2013). Inhibition of pluripotent stem cell-derived teratoma formation by small molecules. PNAS,110(35), E3281-E3290 doi:10.1073/pnas.1303669110
Chad Tang,
Irving L. Weissman
, and Micha Drukker (2012) The Safety of Embryonic Stem Cell Therapy Relies on Teratoma Removal. Oncotarget; 3(1): 7-8.
Julie Mathieu, Zhan Zhang, Angelique Nelson, et al. and Hannele Ruohola-Baker (2013) Hypoxia Induces Re-Entry of Committed Cells into Pluripotency. STEM CELLS
doi
:
Chaffer, C.L., Brueckmann, I., Scheel, C., Kaestli, A.J., Wiggins, P.A., Rodrigues, L.O., Brooks, M., Reinhardt, F., Su, Y., Polyak, K., et al. (2011). Normal and neoplastic nonstem cells can spontaneously convert to a stem-like state. Proc. Natl. Acad. Sci. 108, 7950-7955
Piyush B. Gupta, Christine M. Fillmore,Guozhi Jiang,Sagi D. Shapira,Kai Tao, Charlotte Kuperwasser,Eric S. Lander (2011) Stochastic State Transitions Give Rise to Phenotypic Equilibrium in Populations of Cancer Cells. Cell, 146 (4), 633—644
Fu W, Wang SJ, Zhou GD et al. and Zhang WJ. (2012) Residual undifferentiated cells during differentiation of induced pluripotent stem cells in vitro and in vivo. Stem Cells and Development, 21(4): 521—529. doi:10.1089/scd.2011.0131.
Arvind Ravi, Peter B. Rahl, et al. and Phillip A. Sharp (2013) Let-7 represses Nr6a1 and a mid-gestation developmental program in adult fibroblasts. Genes & Dev. 27(12): 941—954 doi:10.1101/gad.215376.113
Hongran Wang , Xiaohong Wang, Xueping Xu, Thomas P. Zwaka, Austin J. Cooney (2013) Epigenetic Re-programming of the Germ Cell Nuclear Factor Gene is Required for Proper Differentiation of Induced Pluripotent Cells. STEM CELLS DOI: 10.1002/stem.1367
Yijie Geng, Yongfeng Zhao, Lisa Corinna Schuster, et al., (2015). A Chemical Biology Study of Human Pluripotent Stem Cells Unveils HSPA8 as a Key Regulator of Pluripotency. Stem Cell Reports DOI:
Zhang, J., Tian, X., Peng, C., Yan, C., Li, Y., Sun, M., … & Han, Y. (2018). Transplantation of CREG modified embryonic stem cells improves cardiac function after myocardial infarction in mice. Biochemical and biophysical research communications, 503(2), 482—489.
doi
:
PMID
Justine J Cunningham, Thomas M Ulbright, Martin F Pera & Leendert H J Looijenga (2012) Lessons from human teratomas to guide development of safe stem cell therapies. Nature Biotechnology, 30, 849—857 doi:10.1038/nbt.2329
↑
Kazutoshi Takahashi and Shinya Yamanaka (2013)
от 5 марта 2016 на
Wayback Machine
. Development, 140, 2457—2461. DOI:10.1242/dev.092551
Park, H. S., Hwang, I., Choi, K. A., Jeong, H., Lee, J. Y., & Hong, S. (2015).
от 24 сентября 2015 на
Wayback Machine
. Biomaterials, 39, 47-58
doi
:
Sergio Ruiz, Andres J. Lopez-Contreras, Mathieu Gabut et al., & Oscar Fernandez-Capetillo (2015).
от 27 августа 2015 на
Wayback Machine
. Nature Communications 6, Article number: 8036
doi
:
Ryoko Araki, Masahiro Uda, Yuko Hoki, et al. & Masumi Abe (2013) Negligible immunogenicity of terminally differentiated cells derived from induced pluripotent or embryonic stem cells. Nature, (2013) doi:10.1038/nature11807
Monya Baker (2013) Safety of induced stem cells gets a boost. Fears of immune response have been overestimated. Nature 493, 145 doi:10.1038/493145a
M. Wahlestedt, G. L. Norddahl, G. Sten, et al. & D. Bryder (2013) An epigenetic component of hematopoietic stem cell aging amenable to reprogramming into a young state. Blood, DOI: 10.1182/blood-2012-11-469080
Ohnishi, K., Semi, K., Yamamoto, T., Shimizu, M., Tanaka, A., Mitsunaga, K., … & Yamada, Y. (2014). Premature Termination of Reprogramming In Vivo Leads to Cancer Development through Altered Epigenetic Regulation. Cell, 156(4), 663—677.
doi
:
Shibata, H., Komura, S., Yamada, Y., Sankoda, N., Tanaka, A., Ukai, T., … & Yamada, Y. (2018). In vivo reprogramming drives Kras-induced cancer development. Nature communications, 9(1), 1-16.
PMID
PMC
doi
:
Ocampo, A., Reddy, P., Martinez-Redondo, P., Platero-Luengo, A., Hatanaka, F., Hishida, T., … & Belmonte, J. C. I. (2016). In vivo amelioration of age-associated hallmarks by partial reprogramming. Cell, 167(7), 1719—1733.
PMID
PMC
doi
:
Singh, P. B., & Newman, A. G. (2018). Age reprogramming and epigenetic rejuvenation. Epigenetics & chromatin, 11(1), 1-7.
PMID
PMC
doi
:
Hishida, T., Yamamoto, M., Hishida-Nozaki, Y., Shao, C., Huang, L., Wang, C., … & Belmonte, J. C. I. (2022). In vivo partial cellular reprogramming enhances liver plasticity and regeneration. Cell Reports, 39(4), 110730.
doi
:
Grigorash B.B, van Essen D, Liang G, Grosse L, Emelyanov A, Kang Z, Korablev A, Kanzler B, Molina C, Lopez E, Demidov O.N, Garrido C, Liu F, Saccani S, Bulavin D.V. (2023). p16
High
senescence restricts cellular plasticity during somatic cell reprogramming. Nat Cell Biol.
PMID
doi
:
Olova, N., Simpson, D. J., Marioni, R. E., & Chandra, T. (2019). Partial reprogramming induces a steady decline in epigenetic age before loss of somatic identity. Aging cell, 18(1), e12877.
PMID
PMC
doi
:
Gill, D., Parry, A., Santos, F., Hernando-Herraez, I., Stubbs, T. M., Milagre, I., & Reik, W. (2021). Multi-omic rejuvenation of human cells by maturation phase transient reprogramming. bioRxiv.
doi
:
Browder KC, Reddy P, Yamamoto M, et al. (March 2022). "In vivo partial reprogramming alters age-associated molecular changes during physiological aging in mice".
Nat Aging
.
doi
:
.
Macip, C. C., Hasan, R., Hoznek, V., Kim, J., Metzger, L. E., Sethna, S., & Davidsohn, N. (2023).
от 12 января 2023 на
Wayback Machine
. bioRxiv.
doi
:
Melendez, E., Chondronasiou, D., Mosteiro, L., Martínez de Villarreal, J., Fernández-Alfara, M., Lynch, C. J., … & Serrano, M. (2022). Natural killer cells act as an extrinsic barrier for in vivo reprogramming. Development, 149(8), dev200361.
PMID
doi
:
Rackham O. J., Firas J., Fang H., Oates M. E., Holmes M. L., Knaupp A. S., Suzuki H., Nefzger C. M., Daub C. O., Shin J. W., Petretto E., Forrest A. R., Hayashizaki Y., Polo J. M., Gough J.
(англ.)
// Nature genetics. — 2016. — Vol. 48,
no. 3
. — P. 331—335. —
doi
:
. —
.
[
]
(неопр.)
. Дата обращения: 24 января 2016.
24 января 2016 года.
Ronquist, S., Patterson, G., Brown, M., Chen, H., Bloch, A., Muir, L., … & Rajapakse, I. (2017).
. PNAS.
doi
:
Moreno-Moral, A. (April 2021).
от 9 июля 2021 на
Wayback Machine
. European Biopharmaceutical Review. 43 — 45
от 9 июля 2021 на
Wayback Machine
. Duke-NUS Medical School Communications. MEDICUS. 2021(1)
Kamaraj, U. S., Chen, J., Katwadi, K., Ouyang, J. F., Sun, Y. B. Y., Lim, Y. M., … & Rackham, O. J. (2020). EpiMogrify models H3K4me3 data to identify signaling molecules that improve cell fate control and maintenance. Cell Systems, 11(5), 509—522.
PMID
doi
:
Rukhlenko, O. S., Halasz, M., Rauch, N., Zhernovkov, V., Prince, T., Wynne, K., … & Kholodenko, B. N. (2022). Control of cell state transitions. Nature, 1-11.
PMID
doi
:
Joung, J., Ma, S., Tay, T., Geiger-Schuller, K. R., Kirchgatterer, P. C., Verdine, V. K., ... & Zhang, F. (2023). A transcription factor atlas of directed differentiation. Cell, 186(1), 209-229.
PMID
doi
:
Baghbaderani B. A., Tian X., Neo B. H., Burkall A., Dimezzo T., Sierra G., Zeng X., Warren K., Kovarcik D. P., Fellner T., Rao M. S.
(англ.)
// Stem cell reports. — 2015. — Vol. 5,
no. 4
. — P. 647—659. —
doi
:
. —
.
[
]
Menasché P., Vanneaux V., Fabreguettes J. R., Bel A., Tosca L., Garcia S., Bellamy V., Farouz Y., Pouly J., Damour O., Périer M. C., Desnos M., Hagège A., Agbulut O., Bruneval P., Tachdjian G., Trouvin J. H., Larghero J.
(англ.)
// European heart journal. — 2015. — Vol. 36,
no. 12
. — P. 743—750. —
doi
:
. —
.
[
]
Babu, S. S., Duvvuru, H., Baker, J., Switalski, S., Shafa, M., Panchalingam, K. M., ... & Baghbaderani, B. A. (2023). Characterization of human induced pluripotent stems cells: Current approaches, challenges, and future solutions. Biotechnology Reports, e00784.
Suresh Babu S.
,
Duvvuru H.
,
Baker J.
,
Switalski S.
,
Shafa M.
,
Panchalingam K. M.
,
Dadgar S.
,
Beller J.
,
Ahmadian Baghbaderani B.
(англ.)
// Biotechnology Reports (Amsterdam, Netherlands). — 2023. — March (
vol. 37
). —
P. e00784—00784
. —
doi
:
. —
. —
PMC
.
[
]
↑
Koga, K., Wang, B., & Kaneko, S. (2020). Current status and future perspectives of HLA-edited induced pluripotent stem cells. Inflammation and Regeneration, 40(1), 23-29.
doi
:
PMC
PMID
Di Stasi A., Tey S. K., Dotti G., Fujita Y., Kennedy-Nasser A., Martinez C., Straathof K., Liu E., Durett A. G., Grilley B., Liu H., Cruz C. R., Savoldo B., Gee A. P., Schindler J., Krance R. A., Heslop H. E., Spencer D. M., Rooney C. M., Brenner M. K.
(англ.)
// The New England journal of medicine. — 2011. — Vol. 365,
no. 18
. — P. 1673—1683. —
doi
:
. —
.
[
]
Yagyu S., Hoyos V., Del Bufalo F., Brenner M. K.
(англ.)
// Molecular therapy : the journal of the American Society of Gene Therapy. — 2015. — Vol. 23,
no. 9
. — P. 1475—1485. —
doi
:
. —
.
[
]
Wu C., Hong S. G., Winkler T., Spencer D. M., Jares A., Ichwan B., Nicolae A., Guo V., Larochelle A., Dunbar C. E.
(англ.)
// Molecular therapy. Methods & clinical development. — 2014. — Vol. 1. — P. 14053. —
doi
:
. —
.
[
]
Ando M., Nishimura T., Yamazaki S., Yamaguchi T., Kawana-Tachikawa A., Hayama T., Nakauchi Y., Ando J., Ota Y., Takahashi S., Nishimura K., Ohtaka M., Nakanishi M., Miles J. J., Burrows S. R., Brenner M. K., Nakauchi H.
(англ.)
// Stem cell reports. — 2015. — Vol. 5,
no. 4
. — P. 597—608. —
doi
:
. —
.
[
]
Ivics Z.
(англ.)
// Molecular therapy : the journal of the American Society of Gene Therapy. — 2015. — Vol. 23,
no. 9
. — P. 1417—1420. —
doi
:
. —
.
[
]
Shi, Z. D., Tchao, J., Wu, L., & Carman, A. J. Precision installation of a highly efficient suicide gene safety switch in human induced pluripotent stem cells. Stem cells translational medicine.
PMID
doi
:
↑
Miyawaki S., Kawamura Y., Oiwa Y., Shimizu A., Hachiya T., Bono H., Koya I., Okada Y., Kimura T., Tsuchiya Y., Suzuki S., Onishi N., Kuzumaki N., Matsuzaki Y., Narita M., Ikeda E., Okanoya K., Seino K., Saya H., Okano H., Miura K.
(англ.)
// Nature communications. — 2016. — Vol. 7. — P. 11471. —
doi
:
. —
.
[
]
Takahashi K., Mitsui K., Yamanaka S.
(англ.)
// Nature. — 2003. — Vol. 423,
no. 6939
. — P. 541—545. —
doi
:
. —
.
[
]
Wu, L., Zhao, G., Xu, S., Kuang, J., Ming, J., Wu, G., … & Liu, J. (2021). The nuclear factor CECR2 promotes somatic cell reprogramming by reorganizing the chromatin structure. Journal of Biological Chemistry, 296, 100022.
PMID
PMC
doi
:
Tanabe K., Nakamura M., Narita M., Takahashi K., Yamanaka S.
// Proc Natl Acad Sci U S A. — 2013. —
doi
:
. —
.
[
]
Kumar R., DiMenna L., Schrode N., Liu T. C., Franck P., Muñoz-Descalzo S., Hadjantonakis A. K., Zarrin A. A., Chaudhuri J., Elemento O., Evans T.
(англ.)
// Nature. — 2013. — Vol. 500,
no. 7460
. — P. 89—92. —
doi
:
. —
.
[
]
Zhang H., Jiao W., Sun L., Fan J., Chen M., Wang H., Xu X., Shen A., Li T., Niu B., Ge S., Li W., Cui J., Wang G., Sun J., Fan X., Hu X., Mrsny R. J., Hoffman A. R., Hu J. F.
(англ.)
// Cell stem cell. — 2013. — Vol. 13,
no. 1
. — P. 30—35. —
doi
:
. —
.
[
]
Vidal, S. E., Polyzos, A., Chatterjee, K., Ee, L. S., Swanzey, E., Morales-Valencia, J., … & Stadtfeld, M. (2020). Context-dependent requirement of euchromatic histone methyltransferase activity during reprogramming to pluripotency. Stem cell reports, 15(6), 1233—1245.
PMID
PMC
doi
:
Deng, P., Yuan, Q., Cheng, Y., Li, J., Liu, Z., Liu, Y., … & Wang, C. Y. (2021). Loss of KDM4B exacerbates bone-fat imbalance and mesenchymal stromal cell exhaustion in skeletal aging. Cell Stem Cell. S1934-5909(21)00010-2
PMID
doi
:
Wei J, Antony J, Meng F, MacLean P, Rhind R, Laible G, Oback B (2017) KDM4B‐mediated reduction of H3K9me3 and H3K36me3 levels improves somatic cell reprogramming into pluripotency. Sci Rep 7: 7514
PMID
PMC
doi
:
Cheloufi S., Elling U., Hopfgartner B., Jung Y. L., Murn J., Ninova M., Hubmann M., Badeaux A. I., Euong Ang C., Tenen D., Wesche D. J., Abazova N., Hogue M., Tasdemir N., Brumbaugh J., Rathert P., Jude J., Ferrari F., Blanco A., Fellner M., Wenzel D., Zinner M., Vidal S. E., Bell O., Stadtfeld M., Chang H. Y., Almouzni G., Lowe S. W., Rinn J., Wernig M., Aravin A., Shi Y., Park P. J., Penninger J. M., Zuber J., Hochedlinger K.
(англ.)
// Nature. — 2015. — Vol. 528,
no. 7581
. — P. 218—224. —
doi
:
. —
.
[
]
Reynolds N., Salmon-Divon M., Dvinge H., Hynes-Allen A., Balasooriya G., Leaford D., Behrens A., Bertone P., Hendrich B.
(англ.)
// The EMBO journal. — 2012. — Vol. 31,
no. 3
. — P. 593—605. —
doi
:
. —
.
[
]
↑
Luo M., Ling T., Xie W., Sun H., Zhou Y., Zhu Q., Shen M., Zong L., Lyu G., Zhao Y., Ye T., Gu J., Tao W., Lu Z., Grummt I.
(англ.)
// Stem cells (Dayton, Ohio). — 2013. — Vol. 31,
no. 7
. — P. 1278—1286. —
doi
:
. —
.
[
]
Rais Y., Zviran A., Geula S., Gafni O., Chomsky E., Viukov S., Mansour A. A., Caspi I., Krupalnik V., Zerbib M., Maza I., Mor N., Baran D., Weinberger L., Jaitin D. A., Lara-Astiaso D., Blecher-Gonen R., Shipony Z., Mukamel Z., Hagai T., Gilad S., Amann-Zalcenstein D., Tanay A., Amit I., Novershtern N., Hanna J. H.
(англ.)
// Nature. — 2013. — Vol. 502,
no. 7469
. — P. 65—70. —
doi
:
. —
.
[
]
Shao Z., Zhang R., Khodadadi-Jamayran A., Chen B., Crowley M. R., Festok M. A., Crossman D. K., Townes T. M., Hu K.
(англ.)
// Nature communications. — 2016. — Vol. 7. — P. 10869. —
doi
:
. —
.
[
]
Soria-Valles C., Osorio F. G., López-Otín C.
(англ.)
// Cell cycle (Georgetown, Tex.). — 2015. — Vol. 14,
no. 21
. — P. 3345—3346. —
doi
:
. —
.
[
]
MacCarthy CM, Malik V, Wu G, et al., & Velychko S (September 2022).
от 25 сентября 2022 на
Wayback Machine
. bioRxiv.
doi
:
Alvarez‐Palomo, A. B., Requena‐Osete, J., Delgado‐Morales, R., Moreno‐Manzano, V., Grau‐Bove, C., Tejera, A. M., … & Edel, M. J. (2021).
от 1 марта 2022 на
Wayback Machine
. Stem Cells.
PMID
doi
:
Di Stefano B., Sardina J. L., van Oevelen C., Collombet S., Kallin E. M., Vicent G. P., Lu J., Thieffry D., Beato M., Graf T.
(англ.)
// Nature. — 2014. — Vol. 506,
no. 7487
. — P. 235—239. —
doi
:
. —
.
[
]
Di Stefano B., Collombet S., Jakobsen J. S., Wierer M., Sardina J. L., Lackner A., Stadhouders R., Segura-Morales C., Francesconi M., Limone F., Mann M., Porse B., Thieffry D., Graf T.
(англ.)
// Nature cell biology. — 2016. — Vol. 18,
no. 4
. — P. 371—381. —
doi
:
. —
.
[
]
Wang, H., Luo, G., Hu, X., Xu, G., Wang, T., Liu, M., ... & Li, J. (2023). Targeting C/EBPα overcomes primary resistance and improves the efficacy of FLT3 inhibitors in acute myeloid leukaemia. Nature Communications, 14(1), 1882.
PMID
PMC
doi
:
Paz-Priel, I., & Friedman, A. (2011). C/EBPα dysregulation in AML and ALL. Critical Reviews™ in Oncogenesis, 16(1-2).
PMID
PMC
doi
:
Hu, X., Wu, Q., Zhang, J., Kim, J., Chen, X., Hartman, A. A., … & Guo, S. (2021). Reprogramming progressive cells display low CAG promoter activity. STEM CELLS, 39(1), 43-54.
PMID
PMC
doi
:
Esteban, M. A., Wang, T., Qin, B., Yang, J., Qin, D., Cai, J., ... & Pei, D. (2010). Vitamin C enhances the generation of mouse and human induced pluripotent stem cells. Cell stem cell, 6(1), 71-79.
PMID
doi
:
Pavlovic, V., Ciric, M., Petkovic, M., & Golubovic, M. (2023). Vitamin C and epigenetics: A short physiological overview. Open Medicine, 18(1), 20230688. ((PMID|37359134}}
PMC
doi
:
Liu, X., Khan, A., Li, H., Wang, S., Chen, X., & Huang, H. (2022). Ascorbic acid in epigenetic reprogramming. Current Stem Cell Research & Therapy, 17(1), 13-25. PMID: 34264189 DOI: 10.2174/1574888X16666210714152730
Kovatcheva, M., Melendez, E., Chondronasiou, D. et al. (2023). Vitamin B12 is a limiting factor for induced cellular plasticity and tissue repair. Nat Metab
NAKAUCHI Hiromitsu, KAMIYA Akihide, SUZUKI Nao, ITO Keiichi, YAMAZAKI Satoshi (2011)
от 18 октября 2013 на
Wayback Machine
PATENT COOPERATION TREATY APPLICATION, patno: WO2011071085 (A1) ― 2011-06-16 (C12N5/07)
Chan, S., Arpke, R., Filareto, A., Xie, N., Pappas, M., & Penaloza, J. et al. (2018).
. Cell Stem Cell, 23(1), 74-85.e6.
doi
:
Amabile G., Welner R. S., Nombela-Arrieta C., D'Alise A. M., Di Ruscio A., Ebralidze A. K., Kraytsberg Y., Ye M., Kocher O., Neuberg D. S., Khrapko K., Silberstein L. E., Tenen D. G.
(англ.)
// Blood. — 2013. — Vol. 121,
no. 8
. — P. 1255—1264. —
doi
:
. —
.
[
]
Kaufman D. S.
(англ.)
// Blood. — 2013. — Vol. 121,
no. 8
. — P. 1245—1246. —
doi
:
. —
.
[
]
Masao Tsukada et al., (2017).
. Stem Cell Reports
doi
:
Tateno Hiroaki.
(англ.)
// Glycoscience: Biology and Medicine. — 2014. —
P. 1—8
. —
doi
:
.
[
]
Tateno H., Onuma Y., Ito Y., Minoshima F., Saito S., Shimizu M., Aiki Y., Asashima M., Hirabayashi J.
(англ.)
// Stem cell reports. — 2015. — Vol. 4,
no. 5
. — P. 811—820. —
doi
:
. —
.
[
]
Dabir D. V., Hasson S. A., Setoguchi K., Johnson M. E., Wongkongkathep P., Douglas C. J., Zimmerman J., Damoiseaux R., Teitell M. A., Koehler C. M.
(англ.)
// Developmental cell. — 2013. — Vol. 25,
no. 1
. — P. 81—92. —
doi
:
. —
.
[
]
Suzuki N., Yamazaki S., Yamaguchi T., Okabe M., Masaki H., Takaki S., Otsu M., Nakauchi H.
(англ.)
// Molecular therapy : the journal of the American Society of Gene Therapy. — 2013. — Vol. 21,
no. 7
. — P. 1424—1431. —
doi
:
. —
.
[
]
Marcus M. E., Leonard J. N.
(англ.)
// Pharmaceuticals (Basel, Switzerland). — 2013. — Vol. 6,
no. 5
. — P. 659—680. —
doi
:
. —
.
[
]
Forster R., Chiba K., Schaeffer L., Regalado S. G., Lai C. S., Gao Q., Kiani S., Farin H. F., Clevers H., Cost G. J., Chan A., Rebar E. J., Urnov F. D., Gregory P. D., Pachter L., Jaenisch R., Hockemeyer D.
(англ.)
// Stem cell reports. — 2014. — Vol. 2,
no. 6
. — P. 838—852. —
doi
:
. —
.
[
]
Goldstein R. S.
(англ.)
// Stem cell reviews. — 2010. — Vol. 6,
no. 3
. — P. 473—483. —
doi
:
. —
.
[
]
Joel M., Sandberg C. J., Boulland J. L., Vik-Mo E. O., Langmoen I. A., Glover J. C.
(англ.)
// Developmental dynamics : an official publication of the American Association of Anatomists. — 2013. — Vol. 242,
no. 9
. — P. 1078—1093. —
doi
:
. —
.
[
]
Gun-Sik Cho et al., (2017).
. Nature Protocols 12, 2097—2109
doi
:
(неопр.)
. Дата обращения: 14 сентября 2014.
14 сентября 2014 года.
Qiu X., Yang J., Liu T., Jiang Y., Le Q., Lu Y.
(англ.)
// Public Library of Science ONE. — 2012. — Vol. 7,
no. 3
. — P. e32612. —
doi
:
. —
.
[
]
Hirami Y., Osakada F., Takahashi K., Okita K., Yamanaka S., Ikeda H., Yoshimura N., Takahashi M.
(англ.)
// Neuroscience letters. — 2009. — Vol. 458,
no. 3
. — P. 126—131. —
doi
:
. —
.
[
]
Buchholz D. E., Hikita S. T., Rowland T. J., Friedrich A. M., Hinman C. R., Johnson L. V., Clegg D. O.
(англ.)
// Stem cells (Dayton, Ohio). — 2009. — Vol. 27,
no. 10
. — P. 2427—2434. —
doi
:
. —
.
[
]
Reichman S., Terray A., Slembrouck A., Nanteau C., Orieux G., Habeler W., Nandrot E. F., Sahel J. A., Monville C., Goureau O.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 2014. — Vol. 111,
no. 23
. — P. 8518—8523. —
doi
:
. —
.
[
]
Zhong X., Gutierrez C., Xue T., Hampton C., Vergara M. N., Cao L. H., Peters A., Park T. S., Zambidis E. T., Meyer J. S., Gamm D. M., Yau K. W., Canto-Soler M. V.
(англ.)
// Nature communications. — 2014. — Vol. 5. — P. 4047. —
doi
:
. —
.
[
]
Yang Jin, Nong Eva, Tsang Stephen H.
(англ.)
// Expert Review of Ophthalmology. — 2013. — February (
vol. 8
,
no. 1
). —
P. 5—8
. —
ISSN
. —
doi
:
.
[
]
Mark A. Fields, John Hwang, Jie Gong, Hui Cai, and Lucian V.
Chapter 1: The Eye as a Target Organ for Stem Cell Therapy
// Stem Cell Biology and Regenerative Medicine in Ophthalmology. — Springer, 2013. — P. 1—30. —
ISBN 9781461454939
.
Carr A. J., Vugler A. A., Hikita S. T., Lawrence J. M., Gias C., Chen L. L., Buchholz D. E., Ahmado A., Semo M., Smart M. J., Hasan S., da Cruz L., Johnson L. V., Clegg D. O., Coffey P. J.
(англ.)
// Public Library of Science ONE. — 2009. — Vol. 4,
no. 12
. — P. e8152. —
doi
:
. —
.
[
]
Li Y., Tsai Y. T., Hsu C. W., Erol D., Yang J., Wu W. H., Davis R. J., Egli D., Tsang S. H.
(англ.)
// Molecular medicine (Cambridge, Mass.). — 2012. — Vol. 18. — P. 1312—1319. —
doi
:
. —
.
[
]
Wagner D. E., Bonvillain R. W., Jensen T., Girard E. D., Bunnell B. A., Finck C. M., Hoffman A. M., Weiss D. J.
(англ.)
// Respirology (Carlton, Vic.). — 2013. — Vol. 18,
no. 6
. — P. 895—911. —
doi
:
. —
.
[
]
Firth A. L., Dargitz C. T., Qualls S. J., Menon T., Wright R., Singer O., Gage F. H., Khanna A., Verma I. M.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 2014. — Vol. 111,
no. 17
. — P. 1723—1730. —
doi
:
. —
.
[
]
Wong A. P., Rossant J.
(англ.)
// Current pathobiology reports. — 2013. — Vol. 1,
no. 2
. — P. 137—145. —
doi
:
. —
.
[
]
Mou H., Zhao R., Sherwood R., Ahfeldt T., Lapey A., Wain J., Sicilian L., Izvolsky K., Musunuru K., Cowan C., Rajagopal J.
(англ.)
// Cell stem cell. — 2012. — Vol. 10,
no. 4
. — P. 385—397. —
doi
:
. —
.
[
]
Ghaedi M., Calle E. A., Mendez J. J., Gard A. L., Balestrini J., Booth A., Bove P. F., Gui L., White E. S., Niklason L. E.
(англ.)
// The Journal of clinical investigation. — 2013. — Vol. 123,
no. 11
. — P. 4950—4962. —
doi
:
. —
.
[
]
Ghaedi M., Mendez J. J., Bove P. F., Sivarapatna A., Raredon M. S., Niklason L. E.
(англ.)
// Biomaterials. — 2014. — Vol. 35,
no. 2
. — P. 699—710. —
doi
:
. —
.
[
]
Huang S. X., Islam M. N., O'Neill J., Hu Z., Yang Y. G., Chen Y. W., Mumau M., Green M. D., Vunjak-Novakovic G., Bhattacharya J., Snoeck H. W.
(англ.)
// Nature biotechnology. — 2014. — Vol. 32,
no. 1
. — P. 84—91. —
doi
:
. —
.
[
]
Yuan T., Liao W., Feng N. H., Lou Y. L., Niu X., Zhang A. J., Wang Y., Deng Z. F.
(англ.)
// Stem cell research & therapy. — 2013. — Vol. 4,
no. 3
. — P. 73. —
doi
:
. —
.
[
]
Lam A. Q., Freedman B. S., Morizane R., Lerou P. H., Valerius M. T., Bonventre J. V.
(англ.)
// Journal of the American Society of Nephrology : JASN. — 2014. — Vol. 25,
no. 6
. — P. 1211—1225. —
doi
:
. —
.
[
]
Toyohara T., Mae S., Sueta S., Inoue T., Yamagishi Y., Kawamoto T., Kasahara T., Hoshina A., Toyoda T., Tanaka H., Araoka T., Sato-Otsubo A., Takahashi K., Sato Y., Yamaji N., Ogawa S., Yamanaka S., Osafune K.
(англ.)
// Stem cells translational medicine. — 2015. — Vol. 4,
no. 9
. — P. 980—992. —
doi
:
. —
.
[
]
Carroll, S. H., Wigner, N. A., Kulkarni, N., Johnston-Cox, H., Gerstenfeld, L. C., and Ravid, K. (2012). A2B adenosine receptor promotes mesenchymal stem cell differentiation to osteoblasts and bone formation in vivo. J. Biol. Chem. 287, 15718-15727.
Evans, B., & Ham, J. (2012). An emerging role for adenosine and its receptors in bone homeostasis. Frontiers in endocrinology, 3, 113.
Heemin Kang, Yu-Ru V. Shih, Manando Nakasaki, Harsha Kabra and Shyni Varghese (2016).
от 19 октября 2020 на
Wayback Machine
. Science Advances, 2(8), e1600691
doi
:
Rossant, J. (2015). Mouse and human blastocyst-derived stem cells: vive les differences. Development, 142(1), 9-12.
doi
:
Davidson, K. C., Mason, E. A., & Pera, M. F. (2015). The pluripotent state in mouse and human. Development, 142(18), 3090-3099.
doi
:
Pastor, W.A., Chen, D., Liu, W., Kim, R., Sahakyan, A., Lukianchikov, A., Plath, K., Jacobsen, S.E., and Clark, A.T. (2016).
. Cell Stem Cell, 18(3), 323—329 DOI:
Han Qin, Miroslav Hejna, Yanxia Liu, et al., & Miguel Ramalho-Santos (2016).
. Cell Reports. DOI:
Smagghe, B. J., Stewart, A. K., Carter, M. G., Shelton, L. M., Bernier, K. J., Hartman, E. J., … & DiNardo, B. A. (2013).
. PloS one, 8(3), e58601.
Carter, M.G., Smagghe, B.J., Stewart, A.K., Rapley, J.A., Lynch, E., Bernier, K.J., Keating, K.W., Hatziioannou, V.M., Hartman, E.J. and Bamdad, C. C. (2016), A Primitive Growth Factor, NME7AB, Is Sufficient to Induce Stable Naïve State Human Pluripotency; Reprogramming in This Novel Growth Factor Confers Superior Differentiation. STEM CELLS. doi: 10.1002/stem.2261
Zimmerlin, L., Park, T. S., Huo, J. S., Verma, K., Pather, S. R., Talbot, C. C., … & Guo, H. (2016). Tankyrase inhibition promotes a stable human naïve pluripotent state with improved functionality. Development, 143(23), 4368-4380.
doi
:
PMC
Park, T.S., Zimmerlin, L., Evans-Moses, R. et al. (2020). Vascular progenitors generated from tankyrase inhibitor-regulated naïve diabetic human iPSC potentiate efficient revascularization of ischemic retina. Nat Commun 11, 1195
Jun Wu, Daiji Okamura, Mo Li, et al., & Juan Carlos Izpisua Belmonte (2015). An alternative pluripotent state confers interspecies chimaeric competency. Nature,
doi
:
(неопр.)
. Дата обращения: 9 мая 2015.
8 мая 2015 года.
(неопр.)
. Дата обращения: 7 мая 2015.
10 мая 2015 года.
(неопр.)
. Дата обращения: 8 сентября 2020.
4 марта 2021 года.
Tonge P. D., Corso A. J., Monetti C., Hussein S. M., Puri M. C., Michael I. P., Li M., Lee D. S., Mar J. C., Cloonan N., Wood D. L., Gauthier M. E., Korn O., Clancy J. L., Preiss T., Grimmond S. M., Shin J. Y., Seo J. S., Wells C. A., Rogers I. M., Nagy A.
(англ.)
// Nature. — 2014. — Vol. 516,
no. 7530
. — P. 192—197. —
doi
:
. —
.
[
]
↑
Li Qian, Yu Huang, C. Ian Spencer, Amy Foley, Vasanth Vedantham, Lei Liu, Simon J. Conway, Ji-dong Fu & Deepak Srivastava. (2012) In vivo reprogramming of murine cardiac fibroblasts into induced cardiomyocytes. Nature, Nature; 485, 593—598.
doi
:
Eva Szabo, et al & Mickie Bhatia (2010) Direct conversion of human fibroblasts to multilineage blood progenitors. Nature 468, 521—526
PMID
doi
:
Jem A. Efe, et al & Sheng Ding (2011) Conversion of mouse fibroblasts into cardiomyocytes using a direct reprogramming strategy Nature Cell Biology 13, 215—222
PMID
doi
:
↑
Lujan E, Chanda S, Ahlenius H, Sudhof TC, Wernig M.(2012) Direct conversion of mouse fibroblasts to self-renewing, tripotent neural precursor cells. PNAS; 109(7), 2527—2532. doi: 10.1073/pnas.1121003109
↑
Thier M, Wörsdörfer P, Lakes YB, et al. Direct conversion of fibroblasts into stably expandable neural stem cells. Cell Stem Cell 2012; 10(4),473-479 doi: 10.1016/j.stem.2012.03.003
↑
Han DW, Tapia N., Hermann A., et al. & Schöler H.R. (2012) Direct Reprogramming of Fibroblasts into Neural Stem Cells by Defined Factors. Cell Stem Cell, 10(4), 465—472, doi: 10.1016/j.stem.2012.02.021
Taylor SM, Jones PA. (1979) Multiple new phenotypes induced in 10T1/2 and 3T3 cells treated with 5-azacytidine. Cell;17:771-779.
Lassar AB, Paterson BM, Weintraub H. (1986) Transfection of a DNA locus that mediates the conversion of 10T1/2 fibroblasts to myoblasts. Cell.;47(5):649-56.
Davis RL, Weintraub H, Lassar AB. Expression of a single transfected cDNA converts fibroblasts to myoblasts. Cell. 1987;51:987-1000.
Weintraub, H., Tapscott, S. J., Davis, R. L., Thayer, M. J., Adam, M. A., Lassar, A. B. and Miller, A. D. (1989) Activation of muscle-specific genes in pigment, nerve, fat, liver, and fibroblast cell-lines by forced expression of Myod. Proc. Natl. Acad. Sci. U.S.A. 86, 5434-5438.
Thomas Vierbuchen and Marius Wernig (2011) Direct Lineage Conversions: Unnatural but useful? Nat Biotechnol.; 29(10): 892—907.
doi
:
.
Han D.W., Tapia N., Hermann A., et al. & Schöler H.R. Direct Reprogramming of Fibroblasts into Neural Stem Cells by Defined Factors.Stem Cells Dev. 2012 Mar 1;21(4):521-9.
PMID
doi
:
Bar-Nur O., et al.,& Hochedlinger K. (2018).
. Stem Cell Reports, 10(5), 1505—1521
doi
:
Gatto, N., Dos Santos Souza, C., Shaw, A. C., Bell, S. M., Myszczynska, M. A., Powers, S., … & Azzouz, M. Directly converted astrocytes retain the ageing features of the donor fibroblasts and elucidate the astrocytic contribution to human CNS health and disease. Aging Cell, e13281.
Prasad A, Boon Loong Teh D, Shah Jahan FR, Manivannan J, Chua SM, and All AH (2016). Direct Conversion through Trans-differentiation: Efficacy and Safety. Stem Cells and Development.,
doi
:
.
Horisawa, K., & Suzuki, A. (2020). Direct cell-fate conversion of somatic cells: Toward regenerative medicine and industries. Proceedings of the Japan Academy. Series B, Physical and Biological Sciences, 96(4), 131—158.
doi
:
PMC
PMID
Vashe Chandrakanthan et al., (2016).
от 8 апреля 2016 на
Wayback Machine
. Proceedings of the National Academy of Sciences.
doi
:
Giurumescu, C. A., & Chisholm, A. D. (2011). Cell Identification and Cell Lineage Analysis. Caenorhabditis Elegans: Molecular Genetics and Development, 106, 323—341
McGhee, J. D. (2013), The Caenorhabditis elegans intestine. WIREs Dev Biol, 2: 347—367. doi: 10.1002/wdev.93
Riddle, M. R., Weintraub, A., Nguyen, K. C., Hall, D. H., & Rothman, J. H. (2013). Transdifferentiation and remodeling of post-embryonic C. elegans cells by a single transcription factor. Development, 140(24), 4844-4849 doi: 10.1242/dev.103010
Стасевич К. (2013)
от 12 декабря 2013 на
Wayback Machine
// Компьютерра
Konermann S., Brigham M. D., Trevino A. E., Joung J., Abudayyeh O. O., Barcena C., Hsu P. D., Habib N., Gootenberg J. S., Nishimasu H., Nureki O., Zhang F.
(англ.)
// Nature. — 2015. — Vol. 517,
no. 7536
. — P. 583—588. —
doi
:
. —
.
[
]
Liu, P., Chen, M., Liu, Y., Qi, L. S., & Ding, S. (2018).
от 1 июня 2019 на
Wayback Machine
. Cell Stem Cell.
doi
:
(неопр.)
. Дата обращения: 24 января 2018.
25 января 2018 года.
Wei Shu, Zou Qingjian, Lai Sisi, Zhang Quanjun, Li Li, Yan Quanmei, Zhou Xiaoqing , Zhong Huilin, Lai Liangxue.
// Scientific Reports. — 2016. — Vol. 6. — P. 19648. —
doi
:
. —
.
[
]
Black Joshua B., Adler Andrew F., Wang Hong-Gang, D’Ippolito Anthony M., Hutchinson Hunter A., Reddy Timothy E., Pitt Geoffrey S., Leong Kam W., Gersbach Charles A.
(англ.)
// Cell Stem Cell. — 2016. — September (
vol. 19
,
no. 3
). —
P. 406—414
. —
ISSN
. —
doi
:
.
[
]
Jung D. W., Williams D. R.
(англ.)
// ACS chemical biology. — 2011. — Vol. 6,
no. 6
. — P. 553—562. —
doi
:
. —
.
[
]
Kim W. H., Jung D. W., Kim J., Im S. H., Hwang S. Y., Williams D. R.
(англ.)
// ACS chemical biology. — 2012. — Vol. 7,
no. 4
. — P. 732—743. —
doi
:
. —
.
[
]
↑
Jung D. W., Williams D. R.
(англ.)
// ACS chemical biology. — 2012. — Vol. 7,
no. 11
. — P. 1773—1790. —
doi
:
. —
.
[
]
(неопр.)
. Дата обращения: 23 апреля 2013.
1 февраля 2014 года.
Xie J., Zhang H., Yea K., Lerner R. A.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 2013. — Vol. 110,
no. 20
. — P. 8099—8104. —
doi
:
. —
.
[
]
Zhang H., Wilson I. A., Lerner R. A.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 2012. — Vol. 109,
no. 39
. — P. 15728—15733. —
doi
:
. —
.
[
]
(неопр.)
. Дата обращения: 13 сентября 2017.
13 сентября 2017 года.
Blanchard et al., & Baldwin (2017).
. Nature Biotechnology
doi
:
Ito, N., & Ohta, K. (2015).
от 9 февраля 2018 на
Wayback Machine
. Development, growth & differentiation, 57(4), 305—312
PMID
doi
:
от 9 февраля 2018 на
Wayback Machine
. Technology Networks. NEWS Feb 07, 2018
Chapman S., Liu X., Meyers C., Schlegel R. and McBride A. A. (2010) Human keratinocytes are efficiently immortalized by a Rho kinase inhibitor. J Clin Invest.;120(7):2619-2626. doi:10.1172/JCI42297
Liu, X., Krawczyk, E., Suprynowicz, F. A., Palechor-Ceron, N., Yuan, H., Dakic, A., … & Lu, J. (2017).
от 2 февраля 2020 на
Wayback Machine
. Nature protocols, 12(2), 439—451
doi
:
от 23 октября 2013 на
Wayback Machine
Examining the biological consequences of DNA damage caused by irradiated J2-3T3 fibroblast feeder cells and HPV16: characterisation of the biological functions of Mll. Doctoral thesis, UCL (University College London)
Irena Szumiel (2012) Radiation hormesis: Autophagy and other cellular mechanisms International Journal of Radiation Biology. 88(9), 619—628 doi:10.3109/09553002.2012.699698
Hiroshi Kurosawa (2012) Application of Rho-associated protein kinase (ROCK) inhibitor to human pluripotent stem cells. Journal of Bioscience and Bioengineering, 114(6), 577—581
doi
:
Toshimasa Ishizaki, Masayoshi Uehata, Ichiro Tamechika, et al. and Shuh Narumiya (2000)
от 26 февраля 2021 на
Wayback Machine
. Molecular Pharmacology. 57(5), 976—998
So S, Lee Y, Choi J, Kang S, Lee J-Y, Hwang J, et al. (2020) The Rho-associated kinase inhibitor fasudil can replace Y-27632 for use in human pluripotent stem cell research. PLoS ONE 15(5): e0233057.
Terunuma A, Limgala RP, Park CJ, Choudhary I, Vogel JC. (2010) Efficient procurement of epithelial stem cells from human tissue specimens using a Rho-associated protein kinase inhibitor Y-27632. Tissue Eng Part A. ;16(4):1363-1368 doi: 10.1089/ten.tea.2009.0339
Suprynowicz F. A., Upadhyay G., Krawczyk E., et al. and Richard Schlegel. (2012) Conditionally reprogrammed cells represent a stem-like state of adult epithelial cells. PNAS, DOI: 10.1073/pnas.1213241109
Xuefeng Liu, Virginie Ory, Sandra Chapman, et al. & Richard Schlegel (2012) ROCK Inhibitor and Feeder Cells Induce the Conditional Reprogramming of Epithelial Cells. The American Journal of Pathology, 180(2), 599—607
doi
:
Seema Agarwal, David L. Rimm (2012) Making Every Cell Like HeLa: A Giant Step For Cell Culture. The American Journal of Pathology, 180(2), 443—445
doi
:
Palechor-Ceron N, Suprynowicz FA, Upadhyay G, et al. & Schlegel R, Liu X. (2013) Radiation Induces Diffusible Feeder Cell Factor(s) That Cooperate with ROCK Inhibitor to Conditionally Reprogram and Immortalize Epithelial Cells. Am J Pathol.; 183(6), 1862—1870. doi: 10.1016/j.ajpath.2013.08.009
Yann Barrandon, Nicolas Grasset, Andrea Zaffalon, et al. & Ariane Rochat (2012) Capturing epidermal stemness for regenerative medicine. Seminars in Cell & Developmental Biology. 23(8), 937—944
doi
:
Wu, X., Wang, S., Li, M., Li, J., Shen, J., Zhao, Y., … & Kaboli, P. J. (2020). Conditional reprogramming: next generation cell culture. Acta Pharmaceutica Sinica B. 10(8), 1360—1381
doi
:
Hang Yuan, Scott Myers, Jingang Wang, et al & Richard Schlegel. (2012) Use of Reprogrammed Cells to Identify Therapy for Respiratory Papillomatosis. New England Journal of Medicine; 367 (13): 1220—1227 DOI:10.1056/NEJMoa1203055
Sukhbir Kaur, David R. Soto-Pantoja, Erica V. Stein et al. & David D. Roberts.(2013)
от 17 июня 2013 на
Wayback Machine
. Scientific Reports; 3, Article number: 1673
doi
:
Soto-Pantoja, D. R., Ridnour, L. A., Wink, D. A. & Roberts, D. D. (2013) Blockade of CD47 increases survival of mice exposed to lethal total body irradiation. Sci Rep 3, 1038
doi
:
↑
Leo Kurian, Ignacio Sancho-Martinez, Emmanuel Nivet, et al. & Juan Carlos Izpisua Belmonte (2012)
. Nature Methods. doi:10.1038/nmeth.2255
Wang, Y. C., Nakagawa, M., Garitaonandia. et al. & Loring, J. F. (2011). Specific lectin biomarkers for isolation of human pluripotent stem cells identified through array-based glycomic analysis.Cell research, 21(11), 1551—1563. doi: 10.1038/cr.2011.148
Zhang, X., Stojkovic, P., Przyborski, S.,et al. and Stojkovic, M. (2006), Derivation of Human Embryonic Stem Cells from Developing and Arrested Embryos. STEM CELLS, 24: 2669—2676.
doi
:
Tateno, H., Matsushima, A., Hiemori, K., et al., & Hirabayashi, J. (2013). Podocalyxin is a glycoprotein ligand of the human pluripotent stem cell-specific probe rBC2LCN. Stem cells translational medicine, 2(4), 265—273.
doi
:
Suila Heli, Hirvonen Tia, Ritamo Ilja, et al. and Valmu Leena. (2014). Extracellular O-Linked N-Acetylglucosamine Is Enriched in Stem Cells Derived from Human Umbilical Cord Blood. BioResearch, 3(2): 39-44.
doi
:
Perdigoto, C. N., & Bardin, A. J. (2013). Sending the right signal: Notch and stem cells. Biochimica et Biophysica Acta (BBA)-General Subjects, 1830(2), 2307—2322.
Jafar-Nejad, H., Leonardi, J., & Fernandez-Valdivia, R. (2010). Role of glycans and glycosyltransferases in the regulation of Notch signaling. Glycobiology, 20(8), 931—949.
doi
:
Frederico Alisson-Silva, Deivid de Carvalho Rodrigues, Leandro Vairo, et al. and Adriane R Todeschini (2014). Evidences for the involvement of cell surface glycans in stem cell pluripotency and differentiation. Glycobiology 24 (5): 458—468.
doi
:
Hasehira, K., Tateno, H., Onuma, Y., Ito, Y., Asashima, M., & Hirabayashi, J. (2012). Structural and Quantitative Evidence for Dynamic Glycome Shift on Production of Induced Pluripotent Stem Cells. Molecular & Cellular Proteomics,11(12), 1913—1923.
doi
:
Becker-Kojic, Z. A., & Terness, P. (2002). A novel human erythrocyte GPI anchored glycoprotein ACA. Isolation, purification, primary structure determination, molecular parameters of its lipid structure. . Journal of Biological Chemistry, 277, 40472-40478.
doi
:
Z.A.Becker-Kojič, J.Ureña-Peralta, R.Saffrich et al. & M.Stojkovič (2013) Новый гликопротеин АСА — основной регулятор гемопоэза человека. КЛЕТОЧНЫЕ ТЕХНОЛОГИИ В БИОЛОГИИ И МЕДИЦИНЕ, 9(2), 69-84
Z.A.Becker-Kojič, J.R.Ureña-Peralta, I.Zipančić, et al. & M.Stojkovič (2013) Активация поверхностного гликопротеина АСА индуцирует плюрипотентность гемопоэтических клеток-предшественников. КЛЕТОЧНЫЕ ТЕХНОЛОГИИ В БИОЛОГИИ И МЕДИЦИНЕ, 9(2), 85-101
Mikkola, M. (2013)
от 17 октября 2013 на
Wayback Machine
.
19 июля 2011 года.
978-952-10-8444-7
Zheng, Z., Jian, J., Zhang, X., Zara, J. N., Yin, W., Chiang, M., … & Soo, C. (2012). Reprogramming of human fibroblasts into multipotent cells with a single ECM proteoglycan, fibromodulin. Biomaterials, 33(24), 5821-5831.
PMID
doi
:
Yang, P., Li, C., Lee, M., Marzvanyan, A., Zhao, Z. H., Ting, K., … & Zheng, Z. (2020). Photopolymerizable hydrogel encapsulated fibromodulin-reprogrammed cells for muscle regeneration. Tissue engineering. Part A.
Zheng, Z., Li, C., Ha, P., Chang, G. X., Yang, P., Zhang, X., … & Mills, Z. (2019). CDKN2B upregulation prevents teratoma formation in multipotent fibromodulin-reprogrammed cells. Journal of Clinical Investigation, 129(8), 3236-3251.
doi
:
PMC
PMID
Obokata, Haruko; et al. (2014).
20 февраля 2014 года.
. Nature 505(7485): 641—647.doi:10.1038/nature12968
(неопр.)
. Дата обращения: 3 июля 2014.
27 декабря 2020 года.
(рус.)
.
Lenta.ru
(5 августа 2014). Дата обращения: 19 декабря 2014.
19 декабря 2014 года.
(неопр.)
. THE JAPAN TIMES (19 декабря 2014). Дата обращения: 19 декабря 2014.
19 декабря 2014 года.
Torben Redmer, Sebastian Diecke, Tamara Grigoryan, Angel Quiroga-Negreira, Walter Birchmeier, Daniel Besser (2011) E-cadherin is crucial for embryonic stem cell pluripotency and can replace OCT4 during somatic cell reprogramming. EMBO reports , 12, 720—726, doi:10.1038/embor.2011.88
Bedzhov, I., Alotaibi, H., Basilicata, M. F. et al.., & Stemmler, M. P. (2013). Adhesion, but not a specific cadherin code, is indispensable for ES cell and induced pluripotency. Stem cell research, 11(3), 1250—1263
.
Guannan Su, Yannan Zhao, Jianshu Wei, et al. & Jianwu Dai (2013) Direct conversion of fibroblasts into neural progenitor-like cells by forced growth into 3D spheres on low attachment surfaces. Biomaterials, 34(24), 5897-5906
doi
:
Yongqing Liu, Brian Clem, Ewa K. Zuba-Surma, et al. & Douglas C. Dean (2009) Mouse Fibroblasts Lacking RB1 Function Form Spheres and Undergo Reprogramming to a Cancer Stem Cell Phenotype. Cell Stem Cell, 4(4), 336—347
Hein te Riele (2009) Recreating Stem Cells: A Novel Entrance to the Fountain of Youth. Cell Stem Cell, 4(4), 279—280
Nath S. C., Day B., Harper L., et al., & Rancourt D. E. (2021). Fluid Shear Stress Promotes Embryonic Stem Cell Pluripotency via Interplay between β‐catenin and Vinculin in Bioreactor Culture. Stem Cells,
doi
:
Timothy L. Downing, Jennifer Soto, Constant Morez, Timothee Houssin, Ashley Fritz, Falei Yuan, Julia Chu, Shyam Patel, David V. Schaffer, Song Li. Biophysical regulation of epigenetic state and cell reprogramming. Nature Materials, 2013;
doi
:
Yubing Sun, Koh Meng Aw Yong, Luis G. Villa-Diaz, et al. & Jianping Fu(2014). Hippo/YAP-mediated rigidity-dependent motor neuron differentiation of human pluripotent stem cells. Nature Materials
doi
:
Romero, L. O., Massey, A. E., Mata-Daboin, A. D., Sierra-Valdez, F. J., Chauhan, S. C., Cordero-Morales, J. F., & Vásquez, V. (2019).
. Nature communications, 10(1), 1200.
doi
:
PMC
Guilak, F., Cohen, D. M., Estes, B. T., et al. & Chen, C. S. (2009)
. Cell stem cell, 5(1), 17-26. doi: 10.1016/j.stem.2009.06.016
Worley, K., Certo, A., & Wan, L. Q. (2013). Geometry-Force Control of Stem Cell Fate. BioNanoScience, 3(1), 43-51.
doi
:
Aminuddin, N. I., Ahmad, R., Akbar, S. A., & Murphy, B. P. (2016).
. Science and Technology of Advanced Materials, 17(1), 1-43,
doi
:
Massimiliano Caiazzo, Yuya Okawa, Adrian Ranga, Alessandra Piersigilli, Yoji Tabata, Matthias P. Lutolf (2016). Defined three-dimensional microenvironments boost induction of pluripotency. Nature Materials,
doi
:
Roy, B., Yuan, L., Lee, Y., Bharti, A., Mitra, A., & Shivashankar, G. V. (2020). Fibroblast rejuvenation by mechanical reprogramming and redifferentiation. Proceedings of the National Academy of Sciences, 117(19), 10131-10141
doi
:
PMC
PMID
Ankur Singh, Shalu Suri, Ted Lee, et al. & Andrés J García (2013). Adhesion strength-based, label-free isolation of human pluripotent stem cells. Nature Methods, 10, 438—444
doi
:
Sheng C, Zheng Q, Wu J, et al. and Qi Zhou (2012) Generation of dopaminergic neurons directly from mouse fibroblasts and fibroblast-derived neural progenitors. Cell Res; 22:769-772. doi:10.1038/cr.2012.32
Mingliang Zhang , Yuan-Hung Lin , Yujiao Jennifer Sun , Saiyong Zhu10 , Jiashun Zheng , Kai Liu , Nan Cao , Ke Li , Yadong Huang , Sheng Ding (2016).
. Cell Stem Cell, DOI:
Lin Cheng, Wenxiang Hu, Binlong Qiu et al. and Gang Pei (2014).
от 28 марта 2014 на
Wayback Machine
. Cell Research
doi
:
Eva C. Thomaemail, Claudia Merkl, Tobias Heckel, Rachel Haab, Frederic Knoflach, Corinne Nowaczyk, Nicholas Flint, Ravi Jagasia, Sannah Jensen Zoffmann, Hoa Hue Truong, Pascal Petitjean, Sebastian Jessberger, Martin Graf, Roberto Iacone(2014). Chemical Conversion of Human Fibroblasts into Functional Schwann Cells. Stem Cell Reports, 3(4), 539—547, DOI:
↑
Olof Torper, Ulrich Pfisterer, Daniel A. Wolf, et al. and Malin Parmar (2013) Generation of induced neurons via direct conversion in vivo. PNAS, DOI:10.1073/pnas.1303829110
Wenze Niu, Tong Zang, Yuhua Zou, Sanhua Fang, Derek K. Smith, Robert Bachoo, Chun-Li Zhang. In vivo reprogramming of astrocytes to neuroblasts in the adult brain. Nature Cell Biology, 2013; 15 (10): 1164
doi
:
Zhida Su, Wenze Niu, Meng-Lu Liu, Yuhua Zou, Chun-Li Zhang. In vivo conversion of astrocytes to neurons in the injured adult spinal cord. Nature Communications, 2014; 5
doi
:
Paul Luemai, Grace Woodruff, Yaozhi Wang, et al. & Mark H. Tuszynski. (2014).
. Neuron, DOI:
Caiazzo, M., Giannelli, S., Valente, P., Lignani, G., Carissimo, A., Sessa, A., … & Broccoli, V. (2015).
. Stem Cell Reports. 4(1), 25-36, DOI:
Liu G-H, Yi F, Suzuki K, Qu J. and Izpisua Belmonte J C. (2012) Induced neural stem cells: a new tool for studying neural development and neurological disorders. Cell Research 22, 1087—1091.
doi
:
Fadi J Najm, Angela M Lager, Anita Zaremba, et al. & Paul J Tesar (2013) Transcription factor-mediated reprogramming of fibroblasts to expandable, myelinogenic oligodendrocyte progenitor cells. Nature Biotechnology, doi:10.1038/nbt.2561
Nan Yang, J Bradley Zuchero, Henrik Ahlenius, et al. & Marius Wernig (2013) Generation of oligodendroglial cells by direct lineage conversion. Nature Biotechnology, doi:10.1038/nbt.2564
Panagiotis Douvaras, Jing Wang, Matthew Zimmer, et al. & Valentina Fossatiemail (2014).
. Stem Cell Reports. 3(2), 250—259,
DOI:
Chunhui (2012) Turning cardiac fibroblasts into cardiomyocytes in vivo Trends in Molecular Medicine, doi:10.1016/j.molmed.2012.06.009
Chen J X., Krane M, Deutsch M-A, et al. and Sean M. Wu (2012)Inefficient Reprogramming of Fibroblasts into Cardiomyocytes Using Gata4, Mef2c, and Tbx5. Circulation Research.;111: 50-55, doi:10.1161/CIRCRESAHA.112.270264
Ji-Dong Fu, Nicole R. Stone, Lei Liu, et al. & Deepak Srivastava (2013) Direct Reprogramming of Human Fibroblasts toward a Cardiomyocyte-like State. Stem Cell Reports, doi: 10.1016/j.stemcr.2013.07.005
Miyamoto et al., & Ieda (2017)
. Cell Stem Cell,
doi
:
Liu, L., Guo, Y., Tian, S., Lei, I., Gao, W., Li, Z., ... & Wang, Z. (2024). Transient BRD4 degradation improves cardiac reprogramming by inhibiting macrophage/Oncostatin M induced JAK/STAT pathway. bioRxiv, 2023-12.
doi
:
Paul W. Burridge, Gordon Keller, Joseph D. Gold, Joseph C. Wu (2012) Production of De Novo Cardiomyocytes: Human Pluripotent Stem Cell Differentiation and Direct Reprogramming Review Article Cell Stem Cell, 10(1), 16-28
Zhang, Y., Cao, N., Huang, Y., Spencer, C.I., et al., & Srivastava D., Ding S. (2016).
. Cell Stem Cell, 18(3), 368—381, DOI:
Lalit, P.A., Salick, M.R., Nelson, D.O., et al. & Kamp T.J.(2016).
. Cell Stem Cell, 18(3), 354—367 DOI:
Carpenter L. et al. and Watt S. M.(2012) Efficient Differentiation of Human Induced Pluripotent Stem Cells Generates Cardiac Cells That Provide Protection Following Myocardial Infarction in the Rat. Stem Cells and Development. 21(6): 977—986. doi:10.1089/scd.2011.0075
Xiaojun Lian, Cheston Hsiao, Gisela Wilson, Et al and Sean P. Palecek (2012) Robust cardiomyocyte differentiation from human pluripotent stem cells via temporal modulation of canonical Wnt signaling. PNAS 2012 109 (27) E1848-E1857,doi:10.1073/pnas.1200250109.
(неопр.)
. Дата обращения: 24 ноября 2012. Архивировано из
15 ноября 2012 года.
Satsuki Yamada, Timothy J. Nelson, Garvan C. Kane et al. & Andre Terzic (2013) iPS Cell Intervention Rescues Wall Motion Disparity Achieving Biological Cardiac Resynchronization Post-Infarction.The Journal of Physiology, 591, 4335-4349.; DOI:10.1113/jphysiol.2013.252288
Y-F. Hu, J. F. Dawkins, H. C. Cho, E. Marbán, E. Cingolani, (2014).
от 12 июня 2020 на
Wayback Machine
. Sci. Transl. Med. 6, 245ra94
Haixia Wang, Nan Cao, C. Ian Spencer, Baoming Nie, Tianhua Ma, Tao Xu, Yu Zhang, Xiaojing Wang, Deepak Srivastava, ShengDing (20 February 2014).Small Molecules Enable Cardiac Reprogramming of Mouse Fibroblasts with a Single Factor, Oct4. Cell Reports, doi: 10.1016/j.celrep.2014.01.038
Nan Cao, Yu Huang, Jiashun Zheng et al & Deepak Srivastava, Sheng Ding (2016). Conversion of human fibroblasts into functional cardiomyocytes by small molecules. Science,
doi
:
от 11 февраля 2018 на
Wayback Machine
. GLADSTONE INSTITUTES. News Center
Huang, C., Tu, W., Fu, Y., Wang, J., & Xie, X. (2018).
от 7 апреля 2020 на
Wayback Machine
. Cell research,
doi
:
Tung-Ying Lu, Bo Lin, Jong Kim, et al. & Lei Yang (2013) Repopulation of decellularized mouse heart with human induced pluripotent stem cell-derived cardiovascular progenitor cells. Nature Communications, 4, Article number: 2307 doi:10.1038/ncomms3307
Подробный обзор: Budniatzky, I., & Gepstein, L. (2014).
7 апреля 2014 года.
. Stem cells translational medicine, 3(4), 448—457.
doi
:
Pushp, P., Nogueira, D. E., Rodrigues, C. A., Ferreira, F. C., Cabral, J. M., & Gupta, M. K. (2020). A Concise Review on Induced Pluripotent Stem Cell-Derived Cardiomyocytes for Personalized Regenerative Medicine. Stem Cell Reviews and Reports, 1-29.
PMID
doi
:
Funakoshi, S., Yoshida, Y. (2021).
. Arch Toxicol 95, 3633-3650
doi
:
Gun-Sik Cho, Dong I. Lee, et al., & Daniel P. Judge, David A. Kass, Chulan Kwon.(2017).
. Cell Reports, 18(2): 571—582
doi
:
Benjamin D Cosgrove, Penney M Gilbert, Ermelinda Porpiglia et al. & Helen M Blau (Feb. 2014). Rejuvenation of the muscle stem cell population restores strength to injured aged muscles. Nature Medicine,
doi
:
Sousa-Victor, P., Gutarra, S., García-Prat, L., et al. & Muñoz-Cánoves, P. (2014).
. Nature, 506(7488), 316—321
doi
:
Hosoyama, et al. and Masatoshi Suzuki (March, 2014). Derivation of Myogenic Progenitors Directly From Human Pluripotent Stem Cells Using a Sphere-Based Culture. Stem Cells Trans Med.
doi
:
Castell JV, Gomez-Lechon MJ. Liver cell culture techniques. Methods Mol Biol 2009; 481: 35-46.
David C. Hay. (2013)
Stem Cells and Development. doi:10.1089/scd.2013.1500.
Sgodda, M.; Mobus, S.; Hoepfner, J et al. & Cantz, T. (2013) Improved Hepatic Differentiation Strategies for Human Induced Pluripotent Stem Cells. Current Molecular Medicine, 13(5), 842—855
Chen, Y.-F., Tseng, C.-Y., Wang, H.-W., Kuo, H.-C., Yang, V. W. and Lee, O. K. (2012), Rapid generation of mature hepatocyte-like cells from human induced pluripotent stem cells by an efficient three-step protocol. Hepatology, 55: 1193—1203. doi: 10.1002/hep.24790
Massoud Vosough, Eskandar Omidinia, Mahdi Kadivar et al. and Hossein Baharvand (2013) Generation of Functional Hepatocyte-like Cells from Human Pluripotent Stem Cells in a Scalable Suspension Culture. Stem Cells and Development. doi:10.1089/scd.2013.0088
Si-Tayeb, K., Noto, F. K., Nagaoka, M., et al. and Duncan, S. A. (2010), Highly efficient generation of human hepatocyte-like cells from induced pluripotent stem cells. Hepatology, 51: 297—305. doi: 10.1002/hep.23354
Sullivan, G. J., Hay, D. C., Park, I.-H., et al. and Wilmut, I. (2010), Generation of functional human hepatic endoderm from human induced pluripotent stem cells. Hepatology, 51: 329—335. doi: 10.1002/hep.23335
Liu H, Ye Z, Kim Y, Sharkis S, Jang YY. Generation of endoderm-derived human induced pluripotent stem cells from primary hepatocytes. Hepatology 2010; 51: 1810-9.
Sekiya S, Suzuki A. Direct conversion of mouse fibroblasts to hepatocyte-like cells by defined factors. Nature 2011; 475: 390—393
Huang P, He Z, Ji S, et al. Induction of functional hepatocyte-like cells from mouse fibroblasts by defined factors. Nature 2011;475: 386-9
Jing Shan, Robert E Schwartz, Nathan T Ross, et al. & Sangeeta N Bhatia (2013) Identification of small molecules for human hepatocyte expansion and iPS differentiation. Nature Chemical Biology doi:10.1038/nchembio.1270
Takayama K., Nagamoto Y., Mimura N., et al. & Mizuguchi H. (2013) Long-Term Self-Renewal of Human ES/iPS-Derived Hepatoblast-like Cells on Human Laminin 111-Coated Dishes. Stem Cell Reports, doi: 10.1016/j.stemcr.2013.08.006
Cameron K., Tan R., Schmidt-Heck W., et al & Hay D.C. (2015).
. Stem Cell Reports DOI:
Ruiz J. C., Ludlow J. W., Sherwood S., et al. and Gimble J. M. (2010) Differentiated human adipose-derived stem cells exhibit hepatogenic capability in vitro and in vivo. J. Cell. Physiol., 225(2), 429—436 DOI: 10.1002/jcp.22216 22216
Lis, V. M., & Castell, J. V. (2013)
от 24 февраля 2017 на
Wayback Machine
. Biomaterials for Stem Cell Therapy: State of Art and Vision for the Future, 249—278
Xu, Dan ; Nishimura, Toshihiko ; Zheng, Ming et al. & Peltz, Gary (2013) Enabling Autologous Human Liver Regeneration With Differentiated Adipocyte Stem Cells. Cell Transplantation
Ngan F. Huang (2013) Tissue Engineering and Regenerative Medicine: Role of Extracellular Matrix Microenvironment. Stem Cells and Cancer Stem Cells, 9, 313—323 DOI 10.1007/978-94-007-5645-8_30
Maher JJ, Bissell DM. (1993) Cell-matrix interactions in liver. Semin Cell Biol ; 4(3): 189—201
doi
:
Takanori Takebe, Keisuke Sekine, Masahiro Enomura, et al. & Hideki Taniguchi (2013) Vascularized and functional human liver from an iPSC-derived organ bud transplant. Nature doi:10.1038/nature12271
(неопр.)
. Дата обращения: 6 июля 2013. Архивировано из
6 октября 2014 года.
(неопр.)
. Дата обращения: 28 февраля 2014.
28 февраля 2014 года.
Saiyong Zhu, Milad Rezvani, Jack Harbell, et al. & Sheng Ding (2014). Mouse liver repopulation with hepatocytes generated from human fibroblasts. Nature, doi:10.1038/nature13020
Huang, P., Zhang, L., Gao, Y., He, Z., Yao, D., Wu, Z., … & Hui, L. (2014). Direct Reprogramming of Human Fibroblasts to Functional and Expandable Hepatocytes. Cell stem cell, 14(3), 370—384.
doi
:
Dean Yimlamai, Constantina Christodoulou, Giorgio G. Galli, et al., & Fernando D. Camargoemai (2014).
от 22 августа 2017 на
Wayback Machine
. Cell, 157(6), 1324—1338 DOI:
Katsuda T, Kawamata M, Hagiwara K, Takahashi R, Yamamoto Y, Camargo FD, Ochiya T ().
. Cell Stem Cell,
doi
:
Miyajima, A., Tanaka, M., & Itoh, T. (2014). Stem/Progenitor Cells in Liver Development, Homeostasis, Regeneration, and Reprogramming. Cell Stem Cell, 14(5), 561—574. DOI:
Saiyong Zhu, Holger A. Russ, Xiaojing Wang, Mingliang Zhang, Tianhua Ma, Tao Xu, Shibing Tang, Matthias Hebrok, Sheng Ding. Human pancreatic beta-like cells converted from fibroblasts. Nature Communications, 2016; 7: 10080
doi
:
Abdelalim, E. M., Bonnefond, A., Bennaceur-Griscelli, A., & Froguel, P. (2014). Pluripotent Stem Cells as a Potential Tool for Disease Modelling and Cell Therapy in Diabetes. Stem Cell Reviews and Reports, 1-11.
doi
:
Hrvatin, S., O’Donnell, C. W., Deng, F., et al. & Melton, D. A. (2014). Differentiated human stem cells resemble fetal, not adult, β cells. Proceedings of the National Academy of Sciences, 111(8), 3038-3043.
doi
:
Akinci E, Banga A, Tungatt K, et al. and Slack, J.M. (2013).
от 14 марта 2014 на
Wayback Machine
. PLoS ONE 8(11): e82424.
doi
:
Chen, Y. J., Finkbeiner, S. R., Weinblatt, D., et al. & Stanger, B. Z. (2014). De Novo Formation of Insulin-Producing «Neo-β Cell Islets» from Intestinal Crypts. Cell Reports.,
doi
:
Yin L, Ohanyan V, Pung Y F, and Chilian W M. (2012) Induction of Vascular Progenitor Cells From Endothelial Cells Stimulates Coronary Collateral Growth. Circulation Research.;110:241-252, doi:10.1161/CIRCRESAHA.111.250126
American Heart Association (2012, July 25). Adult stem cells from liposuction used to create blood vessels in the lab. ScienceDaily.
Rekha Samuel, Laurence Daheron, Shan Liao, et al. and Rakesh K. Jain (2013) Generation of functionally competent and durable engineered blood vessels from human induced pluripotent stem cells. PNAS doi:10.1073/pnas.1310675110
Lior Zangi, Kathy O Lui, Alexander von Gise, et al. & Kenneth R Chien.(2013) Modified mRNA directs the fate of heart progenitor cells and induces vascular regeneration after myocardial infarction. Nature Biotechnology,; DOI:10.1038/nbt.2682
Nutan Prasain, Man Ryul Lee, Sasidhar Vemula et al., & Mervin C Yoder (2014)/
. Nature Biotechnology.
doi
:
Caroline E. Hendry, Jessica M. Vanslambrouck, Jessica Ineson, et al. and Melissa H. Little (2013) Direct Transcriptional Reprogramming of Adult Cells to Embryonic Nephron Progenitors. JASN ASN.2012121143; ,doi:10.1681/ASN.2012121143
Xinaris C, Benedetti V, Rizzo P, et al. and Giuseppe Remuzzi (2012)
(недоступная ссылка)
. J Am Soc Nephrol 23: 1857—1868, doi: 10.1681/ASN.2012050505
Pereira, C. F., Chang, B., Qiu, J., Niu, X., Papatsenko, D., Hendry, C. E., … & Moore, K. (2013). Induction of a hemogenic program in mouse fibroblasts. Cell stem cell, 13(2), 205—218. DOI:
Jonah Riddell, Roi Gazit, Brian S. Garrison, et al., & Derrick J. Rossi (2014). Reprogramming Committed Murine Blood Cells to Induced Hematopoietic Stem Cells with Defined Factors. Cell, 157(30, 549—564, DOI:
Е. С. Филоненко, М. А. Лагарькова, С. Л. Киселёв (2013)
от 27 марта 2014 на
Wayback Machine
. КТТИ, 8(2), 6-12 PDF
Focosi, D., Amabile, G., Di Ruscio, A., Quaranta, P., Tenen, D. G., & Pistello, M. (2014).
от 20 мая 2014 на
Wayback Machine
. Blood Cancer Journal (2014) 4, e211; doi:10.1038/bcj.2014.30
Zeuner, A., Martelli, F., et al. and Migliaccio, A. R. (2012), Concise Review: Stem Cell-Derived Erythrocytes as Upcoming Players in Blood Transfusion. STEM CELLS, 30: 1587—1596. doi: 10.1002/stem.1136
Rousseau, G. F., Mazurier, C. and Douay, L. (2016), Culturing red blood cells from stem cells: a solution to present and future challenges of transfusion medicine?. ISBT Science Series, 11: 111—117.
doi
:
Giarratana MC, Rouard H, Dumont A, et al & Luc Douay (2011) Proof of principle for transfusion of in vitro generated red blood cells. Blood; 118(19): 5071-5079. doi: 10.1182/blood-2011-06-362038.
Hirose Sho-ichi, Takayama Naoya, Nakamura Sou, et al. & Eto Koji (2013) Immortalization of Erythroblasts by c-MYC and BCL-XL Enables Large-Scale Erythrocyte Production from Human Pluripotent Stem Cells. Stem Cell Reports, doi: 10.1016/j.stemcr.2013.10.010
Ladan Kobari, Frank Yates, Noufissa Oudrhiri et al. and Luc Douay (2012) Human induced pluripotent stem cells can reach complete terminal maturation: in vivo and in vitro evidence in the erythropoietic differentiation model. Haematologica. 2012; 97:xxx DOI: 10.3324/haematol.2011.055566
Keerthivasan Ganesan , Wickrema A, and Crispino J D (2011) Erythroblast Enucleation Stem Cells Int.; 2011: 139851. doi: 10.4061/2011/139851
Emmanuel Olivier, Caihong Qiu, Eric E. Bouhassira (2012) Protocols and Manufacturing for Cell-Based Therapies Novel, High-Yield Red Blood Cell Production Methods from CD34-Positive Cells Derived from Human Embryonic Stem, Yolk Sac, Fetal Liver, Cord Blood, and Peripheral Blood. Stem Cells Trans Med first published on August 2, 2012;doi:10.5966/sctm.2012-0059
См. также: Migliaccio AR, Whitsett C, Papayannopoulou T, Sadelain M. (2012)
от 11 июля 2012 на
Wayback Machine
Review. Cell Stem Cell.;10(2):115-9
Giani, F. C., Fiorini, C., Wakabayashi, A., Ludwig, L. S., Salem, R. M., Jobaliya, C. D., … & Guo, M. H. (2016). Targeted Application of Human Genetic Variation Can Improve Red Blood Cell Production from Stem Cells. Cell stem cell, 18(1), 73-78
doi
:
Stanford, E. A., Wang, Z., Novikov, O., Mulas, F., Landesman-Bollag, E., Monti, S., … & Sherr, D. H. (2016). The role of the aryl hydrocarbon receptor in the development of cells with the molecular and functional characteristics of cancer stem-like cells. BMC biology, 14(1), 1.
doi
:
Brenden W. Smith, Sarah S. Rozelle, Amy Leung, et al and George J. Murphy (2013) The aryl hydrocarbon receptor directs hematopoietic progenitor cell expansion and differentiation. Blood, blood-2012-11-466722,
doi
:
Sivalingam J., at al., & Oh S.K.W. (2020).
. Stem Cell Reports (in press)
doi
:
Rousseau, G. F., Mazurier, C., & Douay, L. (2016).
от 5 мая 2016 на
Wayback Machine
. ISBT Science Series, 11(S1), 111—117.
doi
:
Mao, B., Lu, X., Huang, S., Yu, J., Lai, M., Tsuji, K., … & Ma, F. (2015). Derivation of Mature Erythrocytes from Human Pluripotent Stem Cells by Coculture with Murine Fetal Stromal Cells. In Hematopoietic Differentiation of Human Pluripotent Stem Cells (pp. 15-39). Springer Netherlands.
doi
:
Fujita, A., Uchida, N., Haro-Mora, J. J., Winkler, T. and Tisdale, J. (2016), β-Globin-Expressing Definitive Erythroid Progenitor Cells Generated from Embryonic and Induced Pluripotent Stem Cell-Derived Sacs. STEM CELLS.
doi
:
Olivier, E., Marenah, L., McCahill, A., Condie, A., Cowan, S., & Mountford, J. C. (2016).
(недоступная ссылка)
. Stem Cells Translational Medicine.
doi
:
Figueiredo C, Goudeva L., Horn P. A., et al and Seltsam A. (2010) Generation of HLA-deficient platelets from hematopoietic progenitor cells. Transfusion.; 50(8): 1690—701. doi: 10.1111/j.1537-2995.2010.02644.x.
Suzuki, D., Flahou, C., Yoshikawa, N., Stirblyte, I., Hayashi, Y., Sawaguchi, A., … & Matsumoto, T. (2020). «iPSC-Derived Platelets Depleted of HLA Class I Are Inert to Anti-HLA Class I and Natural Killer Cell Immunity». Stem cell reports, 14(1), 49-59.
doi
:
PMC
PMID
Sou Nakamura, Naoya Takayama, Shinji Hirata, et al. & Koji Eto. (2014). Expandable Megakaryocyte Cell Lines Enable Clinically Applicable Generation of Platelets from Human Induced Pluripotent Stem Cells. Cell Stem Cell; DOI: 10.1016/j.stem.2014.01.011
Moreau, T., Evans, A. L., Vasquez, L., Tijssen, M. R., Yan, Y., Trotter, M. W., … & Dalby, A. (2016).
от 15 апреля 2016 на
Wayback Machine
. Nature Communications, 7, Article number: 11208
doi
:
Thon JN, Medvetz DA, Karlsson SM, Italiano Jr JE.
от 5 мая 2016 на
Wayback Machine
. J Thromb Haemost 2015; 13 (Suppl. 1): S55-S62.
doi
:
Nurhayati, R. W., Ojima, Y., & Taya, M. (2016). Recent developments in ex vivo platelet production. Cytotechnology, 1-11.
doi
:
Riddell, S.R. & Greenberg, P.D. (1995) Principles for adoptive T cell therapy of human viral diseases. Annu. Rev. Immunol. 13, 545—586 DOI: 10.1146/annurev.iy.13.040195.002553
Toshinobu Nishimura, Shin Kaneko, Ai Kawana-Tachikawa et al. & Hiromitsu Nakauchi (2013) Generation of rejuvenated antigen-specific T cells by pluripotency reprogramming and redifferentiation. Cell Stem Cell, 12(1), 114—126 DOI: 10.1016/j.stem.2012.11.002
Raul Vizcardo, Kyoko Masuda, Daisuke Yamada, et al. & Hiroshi Kawamoto (2013) Regeneration of Human Tumor Antigen-Specific T Cells from iPSCs Derived from Mature CD8+ T Cells . Cell Stem Cell, 12(1), 31-36 DOI:
doi
:
↑
Joseph G. Crompton, Mahendra Rao, Nicholas P. Restifo (2013) Memoirs of a Reincarnated T Cell. Cell Stem Cell, 12(1), 6-8 DOI: 10.1016/j.stem.2012.12.009
Lei F, Haque R, Xiong X, Song J. (2012) Directed differentiation of induced pluripotent stem cells towards T lymphocytes. J Vis Exp. ;(63): e3986. doi: 10.3791/3986
Sadelain, M., Brentjens, R. & Riviere, I. (2013). The basic principles of chimeric antigen receptor design. Cancer Discov. 3, 388—398 doi: 10.1158/2159-8290.CD-12-054
Maria Themeli, Christopher C Kloss, Giovanni Ciriello, et al. & Michel Sadelain (2013) Generation of tumor-targeted human T lymphocytes from induced pluripotent stem cells for cancer therapy. Nature Biotechnology, doi:10.1038/nbt.2678
Karsten A. Pilones, Joseph Aryankalayil, and Sandra Demaria (2012) Invariant NKT Cells as Novel Targets for Immunotherapy in Solid Tumors. Clinical and Developmental Immunology, 2012 , Article ID 720803, doi:10.1155/2012/720803
Watarai H, Yamada D, Fujii S, Taniguchi M, Koseki H. (2012) Induced pluripotency as a potential path towards iNKT cell-mediated cancer immunotherapy. Int J Hematol. ;95(6):624-631. doi: 10.1007/s12185-012-1091-0
Woan K.V., Kim H., Bjordahl R., et al. (2021). Harnessing features of adaptive NK cells to generate iPSC-derived NK cells for enhanced immunotherapy. Cell Stem Cell, In Press,
doi
:
M Haruta, Y Tomita, A Yuno, et al. and S Senju (2012) TAP-deficient human iPS cell-derived myeloid cell lines as unlimited cell source for dendritic cell-like antigen-presenting cells. Gene Therapy, doi:10.1038/gt.2012.59
Fábio F. Rosa, Cristiana F. Pires, Ilia Kurochkin, et al., (2018).
от 31 июля 2021 на
Wayback Machine
. Science Immunology, 3(30), eaau4292
doi
:
Xie, H., Ye, M., Feng, R. & Graf, T (2004)
от 24 сентября 2015 на
Wayback Machine
. Cell 117(5), 663—676 .doi: 10.1016/S0092-8674(04)00419-2
Bussmann, L.H., Schubert, A., Vu Manh, T.P et al. and Graf, T. (2009). A robust and highly efficient immune cell reprogramming system. Cell Stem Cell, 5(5), 554—566. doi: 10.1016/j.stem.2009.10.004
Bruno Di Stefano, Jose Luis Sardina, Chris van Oevelen, et al. & Thomas Graf. (2013) C/EBPα poises B cells for rapid reprogramming into induced pluripotent stem cells. Nature,; DOI:10.1038/nature12885
Rapino F., et al., & Graf T. (2013) C/EBPaInduces Highly Efficient Macrophage Transdifferentiation of B Lymphoma and Leukemia Cell Lines and Impairs Their Tumorigenicity, Cell Reports
Guo, J., Feng, Y., Barnes, P., Huang, F. F., Idell, S., Su, D. M., & Shams, H. (2012). Deletion of FoxN1 in the thymic medullary epithelium reduces peripheral T cell responses to infection and mimics changes of aging. PloS one, 7(4), e34681.
doi
:
Sun, L., Guo, J., Brown, R., Amagai, T., Zhao, Y. and Su, D.-M. (2010),
от 25 мая 2014 на
Wayback Machine
. Aging Cell, 9: 347—357.
doi
:
Nicholas Bredenkamp, Craig S. Nowell and C. Clare Blackburn (April 2014).
от 12 апреля 2014 на
Wayback Machine
. Development, 141, 1627—1637
doi
:
Bredenkamp N., Ulyanchenko S., O’Neill K. E., Manley N. R., Vaidya H. J. & Blackburn C. C. (2014).
. Nature Cell Biology,
doi
:
↑
Peng Y, Huang S, Cheng B, et al. and Fu X. (2012) Mesenchymal stem cells: A revolution in therapeutic strategies of age-related diseases Review Article.
Ageing Research Reviews
, , Available online 30 April 2012, .doi.org/10.1016/j.arr.2012.04.005
Bieback K, Kern S, Kocaomer A et al. (2008) Comparing mesenchymal stromal cells from different human tissues: Bone marrow, adipose tissue and umbilical cord blood. Biomed Mater Eng; 18:S71-S76
Medet Jumabay, Raushan Abdmaulen, Albert Ly, et al. and Kristina I. Boström (January 2014). Pluripotent Stem Cells Derived From Mouse and Human White Mature Adipocytes. Stem Cells Trans Med. sctm.2013-0107 doi:10.5966/sctm.2013-0107
Poloni A, Maurizi G, Leoni P, et al. & Cinti S (2012) Human Dedifferentiated Adipocytes Show Similar Properties to Bone Marrow-Derived Mesenchymal Stem Cells. Stem Cells. ;30(5):965-74.
doi
:
.
Sara M. Melief, Jaap Jan Zwaginga, Willem E. Fibbe and Helene Roelofs (2013) Adipose Tissue-Derived Multipotent Stromal Cells Have a Higher Immunomodulatory Capacity Than Their Bone Marrow-Derived Counterparts. Stem Cells Trans Med May 2013 sctm.2012-0184 doi:10.5966/sctm.2012-0184
Shen JF, Sugawara A, Yamashita J, Ogura H, Sato S. (2011) Dedifferentiated fat cells: an alternative source of adult multipotent cells from the adipose tissues. Int J Oral Sci.;3(3):117-24
Shah, M., George, R. L., Evancho-Chapman, M. M., & Zhang, G. (2016). Current Challenges in Dedifferentiated Fat Cells Research. Organogenesis,
doi
:
(неопр.)
. Дата обращения: 8 сентября 2020.
24 февраля 2021 года.
Anastasia Efimenko, Nina Dzhoyashvili, Natalia Kalinina, Tatiana Kochegura, Renat Akchurin, Vsevolod Tkachuk, Yelena Parfyonova.
Adipose-Derived Mesenchymal Stromal Cells From Aged Patients With Coronary Artery Disease Keep Mesenchymal Stromal Cell Properties but Exhibit Characteristics of Aging and Have Impaired Angiogenic Potential
(англ.)
// Stem Cells Trans Med. — 2014. —
Vol. 3184
. —
P. 32—41
. —
doi
:
.
Stolzing A, Jones E, McGonagle D et al. (2008) Age-related changes in human bone marrow derived mesenchymal stem cells: Consequences for cell therapies. Mech Ageing Dev;129:163-173
Duscher, D., Rennert, R. C., Januszyk, M et al., & Gurtner, G. C. (2014).
от 24 февраля 2015 на
Wayback Machine
. Scientific reports, 4. Article number: 7144
doi
:
Bloor, A.J.C., Patel, A., Griffin, J.E. et al. (2020). Production, safety and efficacy of iPSC-derived mesenchymal stromal cells in acute steroid-resistant graft versus host disease: a phase I, multicenter, open-label, dose-escalation study. Nat Med.
Luzzani, C., Neiman, G., Garate, X., Questa, M., Solari, C., Espinosa, D. F., … & Miriuka, S. G. (2015).
от 1 мая 2018 на
Wayback Machine
. Stem Cell Research & Therapy, 6(1), 1-13.
doi
:
Joana Frobe, Hatim Hemeda, Michael Lenz, et al., & Wolfgang Wagneremai (Sept. 2014). Epigenetic Rejuvenation of Mesenchymal Stromal Cells Derived from Induced Pluripotent Stem Cells. Stem Cell Reports, 3(3), 414—422, doi:
Irina Eberle, Mohsen Moslem, Reinhard Henschler, Tobias Cantz (2012)
от 1 мая 2018 на
Wayback Machine
. Advances in Biochemical Engineering Biotechnology
Diederichs Solvig and TuanRocky S. (April, 2014). Functional Comparison of Human-Induced Pluripotent Stem Cell-Derived Mesenchymal Cells and Bone Marrow-Derived Mesenchymal Stromal Cells from the Same Donor Stem Cells and Development.
doi
:
PMID
PMC
Rogers, R. E., Haskell, A., White, B. P., Dalal, S., Lopez, M., Tahan, D., … & Kaunas, R. A scalable system for generation of mesenchymal stem cells derived from induced pluripotent cells employing bioreactors and degradable microcarriers. Stem cells translational medicine.
PMID
doi
:
Soontararak, S., Chow, L., Johnson, V., Coy, J., Wheat, W., Regan, D., & Dow, S. (2018). Mesenchymal Stem Cells (MSC) Derived from Induced Pluripotent Stem Cells (iPSC) Equivalent to Adipose‐Derived MSC in Promoting Intestinal Healing and Microbiome Normalization in Mouse Inflammatory Bowel Disease Model. Stem cells translational medicine.
Chen Y S, Pelekanos R A., Ellis R L., et al and Nicholas M. Fisk (2012) Small Molecule Mesengenic Induction of Human Induced Pluripotent Stem Cells to Generate Mesenchymal Stem/Stromal Cells Stem Cells Trans Med published online February 7, 2012
doi
:
Millard, S. M. and Fisk, N. M. (2012), Mesenchymal stem cells for systemic therapy: Shotgun approach or magic bullets?. Bioessays.
doi
:
.
Hynes, K., Menicanin, D., Han, J., et al. & Bartold, P. M. (2013). Mesenchymal stem cells from iPS cells facilitate periodontal regeneration. Journal of dental research, 92(9), 833—839.
doi
:
(неопр.)
. Дата обращения: 3 октября 2017.
4 марта 2016 года.
Zou, L., Luo, Y., Chen, M., Wang, G., Ding, M., Petersen, C. C., … & Bünger, C. (2013).
от 7 апреля 2014 на
Wayback Machine
. Scientific reports, 3.
doi
:
↑
Kondo, T. (2021). Selective eradication of pluripotent stem cells by inhibiting DHODH activity. Stem cells, 39(1), 33-42.
PMID
doi
:
↑
Al-Akashi, Z., Zujur, D., Kamiya, D., Kato Jr, T., Kondo, T., & Ikeya, M. (2023). Selective vulnerability of
human
-induced pluripotent stem cells to dihydroorotate dehydrogenase inhibition during mesenchymal stem/stromal cell purification. Frontiers in Cell and Developmental Biology, 11. 11, 1089945
PMID
PMC
doi
:
↑
Pellegrini, S., Zamarian, V., & Sordi, V. (2022). Strategies to Improve the Safety of iPSC-Derived β Cells for β Cell Replacement in Diabetes. Transplant International, 169.
PMID
PMC
doi
:
Zhang, L., Wang, H., Liu, C., Wu, Q., Su, P., Wu, D., … & Zhou, J. (2018).
. Stem Cell Reports. DOI:
Pei-Lun Lai, Hsuan Lin, Shang-Fu Chen, et al., & Jean Lu (2017).
от 19 марта 2017 на
Wayback Machine
. Scientific Reports 7, Article number: 44534
doi
:
Ruenn Chai Lai, Ronne Wee Yeh Yeo, Soon Sim Tan, Bin Zhang, et al. and Sai Kiang Lim (2013) Mesenchymal Stem Cell Exosomes: The Future MSC-Based Therapy? In: Mesenchymal Stem Cell Therapy. Chase, Lucas G.; Vemuri, Mohan C. (Eds.). 39-61 DOI 10.1007/978-1-62703-200-1_3
Ruenn Chai Lai, Ronne Wee Yeh Yeo, Kok Hian Tan, Sai Kiang Lim (2013) Exosomes for drug delivery — a novel application for the mesenchymal stem cell. Biotechnology Advances.
doi
:
Ronne Wee Yeh Yeoa, b, 1, Ruenn Chai Laia, 1, Bin Zhanga, et al. & Sai Kiang Lim (2012)Mesenchymal stem cell: An efficient mass producer of exosomes for drug delivery. Advanced Drug Delivery Reviews
doi
:
Nobuyoshi Kosaka, Fumitaka Takeshita, Yusuke Yoshioka, et al. & Takahiro Ochiya (2012) Exosomal tumor-suppressive microRNAs as novel cancer therapy: «Exocure» is another choice for cancer treatment. Advanced Drug Delivery Reviews
doi
:
Mangeot, Philippe Lotteau, Vincent Peschanski, Marc Girard, Mathilde (Evry Cedex, FR) (2013)
от 17 октября 2013 на
Wayback Machine
United States Patent Application 20130034900
Dominici, M. L. B. K., Le Blanc, K., Mueller, I., Slaper-Cortenbach, I., Marini, F. C., Krause, D. S., … & Horwitz, E. M. (2006). Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy, 8(4), 315—317.
PMID
doi
:
↑
Myret Ghabriel, Ahmed El Hosseiny, Ahmed Moustafa, Asma Amleh (2021).
от 27 мая 2021 на
Wayback Machine
. bioRxiv 2021.05.25.445659; doi:
doi
:
Kaya, H. E. K., & Radhakrishnan, S. K. (2020). Trash Talk: Mammalian Proteasome Regulation at the Transcriptional Level. Trends in Genetics. 37(2), 160—173
PMID
PMC
doi
:
Cheng, A., Hardingham, T. E., & Kimber, S. J. (2013). Generating Cartilage Repair from Pluripotent Stem Cells. Tissue Engineering Part B: Reviews. doi:10.1089/ten.teb.2012.0757
Tsumaki, N. (2015).
от 18 июня 2018 на
Wayback Machine
. In A Tissue Regeneration Approach to Bone and Cartilage Repair (pp. 85-98). Springer International Publishing.
doi
:
Outani H, Okada M, Yamashita A, Nakagawa K, Yoshikawa H, et al. (2013) Direct Induction of Chondrogenic Cells from Human Dermal Fibroblast Culture by Defined Factors. PLoS ONE 8(10): e77365. doi:10.1371/journal.pone.0077365
K. Miyoshi, D. Tsuji, K. Kudoh, et al.& Takafumi Noma (2010) Generation of human induced pluripotent stem cells from oral mucosa J Biosci Bioeng, 110(3), 345—350
doi
:
Katsuhiro Yoshikawa, Motoko Naitoh, Hiroshi Kubota, et al. (2013) Multipotent stem cells are effectively collected from adult human cheek skin. Biochemical and Biophysical Research Communications, 431(1), 104—110
doi
:
Hong-Kee Tana, Cheng-Xu Delon Toha, Dongrui Mab, et al. and Yuin-Han Loh (2014). Human Finger-Prick Induced Pluripotent Stem Cells Facilitate the Development of Stem Cell Banking. Stem Cells Trans Med.
doi
:
↑
Okita, K., Yamakawa T., Matsumura, Y., et al. and Shinya Yamanaka (2012) An Efficient Non-viral Method to Generate Integration-Free Human iPS Cells from Cord Blood and Peripheral Blood Cells. STEM CELLS DOI: 10.1002/stem.1293
Imbisaat Geti, Mark L. Ormiston, Foad Rouhani, et al & Nicholas W. Morrell (2012) A Practical and Efficient Cellular Substrate for the Generation of Induced Pluripotent Stem Cells from Adults: Blood-Derived Endothelial Progenitor Cells. Stem Cells Trans Med. sctm.2012-0093 doi:10.5966/sctm.2012-0093
Judith Staerk, Meelad M. Dawlaty, Qing Gaoet al. and
Rudolf Jaenisch
(2010) Reprogramming of Human Peripheral Blood Cells to Induced Pluripotent Stem Cells. Cell Stem Cell, 7(1), 20-24 doi:10.1016/j.stem.2010.06.002
Park TS, Huo JS, Peters A, Talbot CC Jr, Verma K, et al. (2012) Growth Factor-Activated Stem Cell Circuits and Stromal Signals Cooperatively Accelerate Non-Integrated iPSC Reprogramming of Human Myeloid Progenitors. PLoS ONE 7(8): e42838. doi:10.1371/journal.pone.0042838
Zhou T, Benda C, Duzinger S, Et al & Esteban MA(2011) Generation of induced pluripotent stem cells from urine. J Am Soc Nephrol 22: 1221—1228
Ting Zhou, Christina Benda, Sarah Dunzinger, et al. & Miguel A Esteban (2012) Generation of human induced pluripotent stem cells from urine samples. Nature Protocols. 7(12), 2080—2089
doi
:
Lihui Wang, Linli Wang, Wenhao Huang, & Duanqing Pei (2012) Generation of integration-free neural progenitor cells from cells in human urine. Nature Methods, doi:10.1038/nmeth.2283
. Cai J, Zhang Y, Liu P, Chen S, Wu X, Sun Y, Li A, Huang K et al (2013)
от 26 апреля 2015 на
Wayback Machine
. .Cell Regeneration , 2:6 doi:10.1186/2045-9769-2-6
Sun, W., Hu, X., Wang, L., Ma, Y., Zhang, X., Zhang, R., … & Wang, G. (2022).
от 7 июня 2022 на
Wayback Machine
. Stem Cell Research, 102682.
doi
:
Shantaram Bharadwaj, Guihua Liu, Yingai Shi, et al. & Yuanyuan Zhang (2013) Multi-Potential Differentiation of Human Urine-Derived Stem Cells: Potential for Therapeutic Applications in Urology. STEM CELLS,31(9), 1840—1856
doi
:
Huang, Y. Z., He, T., Cui, J., Jiang, Y. L., Zeng, J. F., Zhang, W. Q., & Xie, H. Q. (2022). Urine-Derived Stem Cells for Regenerative Medicine: Basic Biology, Applications, and Challenges. Tissue Engineering Part B: Reviews.
PMID
doi
:
Culenova, M., Nicodemou, A., Novakova, Z. V., Debreova, M., Smolinská, V., Bernatova, S., … & Danisovic, L. (2021). Isolation, Culture and Comprehensive Characterization of Biological Properties of Human Urine-Derived Stem Cells. International Journal of Molecular Sciences, 22(22), 12503.
PMID
PMC
doi
:
Yimei Wang1, Jinyu Liu1, Xiaohua Tan1, et al. and Yulin Li (2012) Induced Pluripotent Stem Cells from Human Hair Follicle Mesenchymal Stem Cells. Stem Cell Reviews and Reports,.doi:10.1007/s12015-012-9420-5
Raab, S., Klingenstein, M., Liebau, S., & Linta, L. (2014). A comparative view on human somatic cell sources for iPSC generation. Stem Cells International, 2014(2014), Article ID 768391,
Schnabel L. V, Abratte C. M., Schimenti J. C, et al. and Fortier L. A. (2012) Genetic background affects induced pluripotent stem cell generation. Stem Cell Research & Therapy 2012, 3:30 doi:10.1186/scrt121
Panopoulos AD, Ruiz S, Yi F, Herrerías A, Batchelder EM, Izpisua Belmonte JC.(2011) Rapid and highly efficient generation of induced pluripotent stem cells from human umbilical vein endothelial cells. PLoS One;6:e19743
↑
J.M. Polo, S. Liu, M.E. Figueroa, et al. & Konrad Hochedlinger (2010) Cell type of origin influences the molecular and functional properties of mouse induced pluripotent stem cells. Nat Biotechnol, 28, 848—855 doi:10.1038/nbt.1667
Miura K, Okada Y, Aoi T, Okada A, et al & Yamanaka S.(2009) Variation in the safety of induced pluripotent stem cell lines. Nat Biotechnol.;27:743-745
↑
K. Kim, A. Doi, B. Wen, K. Ng, R. Zhao, P. Cahan, J. Kim, M.J. Aryee, H. Ji, L.I. Ehrlich et al. (2010) Epigenetic memory in induced pluripotent stem cells. Nature, 467, 285—290 doi:10.1038/nature09342
↑
K. Kim, R. Zhao, A. Doi, K. Ng, J. Unternaehrer, P. Cahan, H. Huo, Y.H. Loh, M.J. Aryee, M.W. Lensch et al. (2011) Donor cell type can influence the epigenome and differentiation potential of human induced pluripotent stem cells. Nat Biotechnol, 29, pp. 1117—1119
↑
O. Bar-Nur, H.A. Russ, S. Efrat, N. Benvenisty (2011)
от 24 сентября 2015 на
Wayback Machine
in induced pluripotent stem cells derived from human pancreatic islet beta cells. Cell Stem Cell, 9 , 17-23
doi
:
↑
Denker H-W.(2012) Time to Reconsider Stem Cell Induction Strategies. Cells.; 1(4):1293-1312. doi:10.3390/cells1041293
Jong-Hee Lee, Jung Bok Lee, Zoya Shapovalova, Aline Fiebig-Comyn, Ryan R. Mitchell, Sarah Laronde, Eva Szabo, Yannick D. Benoit & Mickie Bhatia (2014).
от 19 января 2016 на
Wayback Machine
Nature Communications 5, Article number: 5605 doi:10.1038/ncomms6605
Васькова, Е. А., Стекленева, А. Е., Медведев, С. П., & Закиян, С. М. (2013).
от 25 февраля 2021 на
Wayback Machine
. Acta Naturae (русскоязычная версия), 5(4 (19)).
Aija Kyttälä, Roksana Moraghebi, Cristina Valensisi, Johannes Kettunen, Colin Andrus, Kalyan Kumar Pasumarthy, Mahito Nakanishi, Ken Nishimura, Manami Ohtaka, Jere Weltner, Ben Van Handel, Olavi Parkkonen, Juha Sinisalo, Anu Jalanko, R. David Hawkins, Niels-Bjarne Woods, Timo Otonkoski, Ras Trokovic (2016). Genetic Variability Overrides the Impact of Parental Cell Type and Determines iPSC Differentiation Potential. Stem Cell Reports,
doi
:
Р. Фрешни (2010) Культура животных клеток. Издательство: Бином. Лаборатория знаний.
ISBN 978-5-94774-596-2
Zhang Y, Wei C, Zhang P, Li X, Liu T, et al. (2014).
от 22 февраля 2014 на
Wayback Machine
. PLoS ONE 9(1): e85089. doi:10.1371/journal.pone.0085089
↑
Masato Nakagawa, Yukimasa Taniguchi, Sho Senda, et al. & Shinya Yamanaka (2014).
от 12 февраля 2014 на
Wayback Machine
. Scientific Reports, 4, Article number: 3594 doi:10.1038/srep03594
Chen, K. G., Mallon, B. S., McKay, R. D., & Robey, P. G. (2014). Human Pluripotent Stem Cell Culture: Considerations for Maintenance, Expansion, and Therapeutics. Cell stem cell, 14(1), 13-26.
Dolley-Sonneville PJ, Romeo LE, Melkoumian ZK (2013) Synthetic Surface for Expansion of Human Mesenchymal Stem Cells in Xeno-Free, Chemically Defined Culture Conditions. PLoS ONE 8(8): e70263. doi:10.1371/journal.pone.0070263
Aumailley M et al. (2005). A simplified laminin nomenclature. Matrix Biol. 24 (5): 326-32.doi:10.1016/j.matbio.2005.05.006.
Bergstrom, R., Strom, S., Holm, F., Feki, A. & Hovatta, O. (2011). Xeno-free culture of human pluripotent stem cells. Methods Mol Biol 767, 125—136
Sergey Rodin, Liselotte Antonsson, Colin Niaudet et al. & Karl Tryggvason (January 2014)
от 2 февраля 2014 на
Wayback Machine
. Nature Communications 5, Article number: 3195 doi:10.1038/ncomms4195
Eric W. Brunner, Izabela Jurewicz, Elena Heister, et al. and Alan B. Dalton (2014). Growth and Proliferation of Human Embryonic Stem Cells on Fully Synthetic Scaffolds Based on Carbon Nanotubes. ACS Applied Materials & Interfaces,; 140123104241006 DOI:10.1021/am405097w
Dixon, J. E., Shah, D. A., Rogers, et al. & Shakesheff, K. M. (2014).
. Proceedings of the National Academy of Sciences,
doi
:
(неопр.)
. Дата обращения: 1 июля 2021.
1 марта 2022 года.
(неопр.)
. Дата обращения: 1 июля 2021.
9 июля 2021 года.
(неопр.)
. Дата обращения: 1 июля 2021.
9 июля 2021 года.
Chen, Y., Tristan, C. A., Chen, L., Jovanovic, V. M., Malley, C., Chu, P. H., … & Singeç, I. (2021). A versatile polypharmacology platform promotes cytoprotection and viability of human pluripotent and differentiated cells. Nature Methods, 18(5), 528—541.
PMID
doi
:
Kejin Hu.(March, 2014). Vectorology and factor delivery in induced pluripotent stem cell reprogramming. Stem Cells and Development.
doi
:
Emilie Bayart and Odile Cohen-Haguenaue (2013)Technological Overview of iPS Induction from Human Adult Somatic Cells Current Gene Therapy,13(2),73-92
Zhang Z., Gao Y., Gordon A., Wang Z. Z., Qian Z., Wu W. S.
(англ.)
// Public Library of Science ONE. — 2011. — Vol. 6,
no. 10
. — P. e26592. —
doi
:
. —
.
[
]
Imamura M., Okuno H., Tomioka I., Kawamura Y., Lin Z. Y., Nakajima R., Akamatsu W., Okano H. J., Matsuzaki Y., Sasaki E., Okano H.
(англ.)
// Methods in molecular biology (Clifton, N.J.). — 2012. — Vol. 925. — P. 21—48. —
doi
:
. —
.
[
]
Nethercott H. E., Brick D. J., Schwartz P. H.
(англ.)
// Methods in molecular biology (Clifton, N.J.). — 2011. — Vol. 767. — P. 67—85. —
doi
:
. —
.
[
]
Shutova Maria V., Chestkov Ilya V., Bogomazova Alexandra N., Lagarkova Maria A., Kiselev Sergey L.
(англ.)
// Springer Protocols Handbooks. — 2011. —
P. 133—149
. —
ISSN
. —
doi
:
.
[
]
от 29 октября 2013 на
Wayback Machine
Min Lu, Cristina Moore, Vi Chu (2011) Enhanced Reprogramming of Human Somatic Cells using Human STEMCCA Polycistronic Lentivirus and Human iPS Cell Boost Supplement
Awe J. P., Lee P. C., Ramathal C., Vega-Crespo A., Durruthy-Durruthy J., Cooper A., Karumbayaram S., Lowry W. E., Clark A. T., Zack J. A., Sebastiano V., Kohn D. B., Pyle A. D., Martin M. G., Lipshutz G. S., Phelps P. E., Pera R. A., Byrne J. A.
(англ.)
// Stem cell research & therapy. — 2013. — Vol. 4,
no. 4
. — P. 87. —
doi
:
. —
.
[
]
Nakanishi M., Otsu M.
(англ.)
// Current gene therapy. — 2012. — Vol. 12,
no. 5
. — P. 410—416. —
.
[
]
Macarthur C. C., Fontes A., Ravinder N., Kuninger D., Kaur J., Bailey M., Taliana A., Vemuri M. C., Lieu P. T.
(англ.)
// Stem cells international. — 2012. — Vol. 2012. — P. 564612. —
doi
:
. —
.
[
]
Fusaki Noemi.
(англ.)
// Stem Cells and Cancer Stem Cells, Volume 9. — 2012. — 21 November. —
P. 59—69
. —
doi
:
.
[
]
(неопр.)
. Дата обращения: 9 ноября 2013.
9 ноября 2013 года.
Koyanagi-Aoi M., Ohnuki M., Takahashi K., Okita K., Noma H., Sawamura Y., Teramoto I., Narita M., Sato Y., Ichisaka T., Amano N., Watanabe A., Morizane A., Yamada Y., Sato T., Takahashi J., Yamanaka S.
(англ.)
// Proceedings of the National Academy of Sciences of the United States of America. — 2013. — Vol. 110,
no. 51
. — P. 20569—20574. —
doi
:
. —
.
[
]
Choi I. Y., Lim H., Lee G.
(англ.)
// Journal of visualized experiments : JoVE. — 2014. —
No. 86
. —
doi
:
. —
.
[
]
Zhou W., Freed C. R.
(англ.)
// Stem cells (Dayton, Ohio). — 2009. — Vol. 27,
no. 11
. — P. 2667—2674. —
doi
:
. —
.
[
]
Loh Y. H., Yang J. C., De Los Angeles A., Guo C., Cherry A., Rossi D. J., Park I. H., Daley G. Q.
(англ.)
// Current protocols in stem cell biology. — 2012. — Vol. Chapter 4. — P. 4—5. —
doi
:
. —
.
[
]
Hudry, E., Martin, C., Gandhi, S., György, B., Scheffer, D. I., Mu, D., … & Masek, M. (2016).
от 23 марта 2017 на
Wayback Machine
. Gene therapy, 23(4), 380—392.
doi
:
György, B., Sage, C., Indzhykulian, A. A., Scheffer, D. I., Brisson, A. R., Tan, S., … & Li, Y. (2017). Rescue of Hearing by Gene Delivery to Inner-Ear Hair Cells Using Exosome-Associated AAV. Molecular Therapy. 25(2), 379—391
doi
:
Warren L., Manos P. D., Ahfeldt T., Loh Y. H., Li H., Lau F., Ebina W., Mandal P. K., Smith Z. D., Meissner A., Daley G. Q., Brack A. S., Collins J. J., Cowan C., Schlaeger T. M., Rossi D. J.
(англ.)
// Cell stem cell. — 2010. — Vol. 7,
no. 5
. — P. 618—630. —
doi
:
. —
.
[
]
Warren L., Ni Y., Wang J., Guo X.
(англ.)
// Scientific reports. — 2012. — Vol. 2. — P. 657. —
doi
:
. —
.
[
]
Luni, C., Giulitti, S., Serena, E., Ferrari, L., Zambon, A., Gagliano, O., … & Elvassore, N. (2016).
от 24 октября 2016 на
Wayback Machine
. Nature methods, 13(5), 446—452.
doi
:
Raimes, W., Rubi, M., Super, A., Marques, M. P., Veraitch, F., & Szita, N. (2016).
. Process Biochemistry.
doi
:
McKinlay, C. J., Vargas, J. R., Blake, T. R., Hardy, J. W., Kanada, M., Contag, C. H., … & Waymouth, R. M. (2017).
от 1 февраля 2017 на
Wayback Machine
. Proceedings of the National Academy of Sciences, 114(4), E448-E456,
doi
:
Vivien C., Scerbo P., Girardot F., Le Blay K., Demeneix B. A., Coen L.
(англ.)
// The Journal of biological chemistry. — 2012. — Vol. 287,
no. 10
. — P. 7427—7435. —
doi
:
. —
.
[
]
Woltjen K., Hämäläinen R., Kibschull M., Mileikovsky M., Nagy A.
(англ.)
// Methods in molecular biology (Clifton, N.J.). — 2011. — Vol. 767. — P. 87—103. —
doi
:
. —
.
[
]
Tsukiyama T., Kato-Itoh M., Nakauchi H., Ohinata Y.
(англ.)
// Public Library of Science ONE. — 2014. — Vol. 9,
no. 3
. — P. e92973. —
doi
:
. —
.
[
]
Kime C., Rand T. A., Ivey K. N., Srivastava D., Yamanaka S., Tomoda K.
(англ.)
// Current protocols in human genetics / editorial board, Jonathan L. Haines ... [et al.]. — 2015. — Vol. 87. — P. 21—22. —
doi
:
. —
.
[
]
Zhang W., Guan D., Qu J., Zhang W., Liu G. H.
(англ.)
// Protein & cell. — 2012. — Vol. 3,
no. 4
. — P. 241—245. —
doi
:
. —
.
[
]
Kim J. H., Lee S. R., Li L. H., Park H. J., Park J. H., Lee K. Y., Kim M. K., Shin B. A., Choi S. Y.
(англ.)
// Public Library of Science ONE. — 2011. — Vol. 6,
no. 4
. — P. e18556. —
doi
:
. —
.
[
]
Gao S. Y., Jack M. M., O'Neill C.
(англ.)
// Public Library of Science ONE. — 2012. — Vol. 7,
no. 11
. — P. e48668. —
doi
:
. —
.
[
]
Qu X., Liu T., Song K., Li X., Ge D.
(англ.)
// Public Library of Science ONE. — 2012. — Vol. 7,
no. 10
. — P. e48161. —
doi
:
. —
.
[
]
Koren E., Torchilin V. P.
(англ.)
// Trends in molecular medicine. — 2012. — Vol. 18,
no. 7
. — P. 385—393. —
doi
:
. —
.
[
]
BR Liu, YW Huang, HJ Chiang, HJ Lee.
Primary effectors in the mechanisms of transmembrane delivery of arginine-rich cell-penetrating peptides // Advanced Studies in Biology. — 2013. — Vol. 5, no. 1—4. — P. 11—25. —
ISSN
.
De Los Angeles A., Daley G. Q.
(англ.)
// Cell research. — 2013. — Vol. 23,
no. 12
. — P. 1337—1338. —
doi
:
. —
.
[
]
Göttlicher M., Minucci S., Zhu P., Krämer O. H., Schimpf A., Giavara S., Sleeman J. P., Lo Coco F., Nervi C., Pelicci P. G., Heinzel T.
(англ.)
// The EMBO journal. — 2001. — Vol. 20,
no. 24
. — P. 6969—6978. —
doi
:
. —
.
[
]
Huangfu D., Maehr R., Guo W., Eijkelenboom A., Snitow M., Chen A. E., Melton D. A.
(англ.)
// Nature biotechnology. — 2008. — Vol. 26,
no. 7
. — P. 795—797. —
doi
:
. —
.
[
]
Sanchez-Ripoll Y., Bone H. K., Owen T., Guedes A. M., Abranches E., Kumpfmueller B., Spriggs R. V., Henrique D., Welham M. J.
(англ.)
// Public Library of Science ONE. — 2013. — Vol. 8,
no. 4
. — P. e60148. —
doi
:
. —
.
[
]
W. Bradley, F. Kevin, R. James.
(англ.)
// Embryonic Stem Cells - Basic Biology to Bioengineering. — 2011. — 15 September. —
doi
:
.
[
]
Hoffmeyer K., Raggioli A., Rudloff S., Anton R., Hierholzer A., Del Valle I., Hein K., Vogt R., Kemler R.
(англ.)
// Science (New York, N.Y.). — 2012. — Vol. 336,
no. 6088
. — P. 1549—1554. —
doi
:
. —
.
[
]
Ichida J. K., Blanchard J., Lam K., Son E. Y., Chung J. E., Egli D., Loh K. M., Carter A. C., Di Giorgio F. P., Koszka K., Huangfu D., Akutsu H., Liu D. R., Rubin L. L., Eggan K.
(англ.)
// Cell stem cell. — 2009. — Vol. 5,
no. 5
. — P. 491—503. —
doi
:
. —
.
[
]
Maherali N., Hochedlinger K.
(англ.)
// Current biology : CB. — 2009. — Vol. 19,
no. 20
. — P. 1718—1723. —
doi
:
. —
.
[
]
Lee M. G., Wynder C., Schmidt D. M., McCafferty D. G., Shiekhattar R.
(англ.)
// Chemistry & biology. — 2006. — Vol. 13,
no. 6
. — P. 563—567. —
doi
:
. —
.
[
]
Li K. K., Luo C., Wang D., Jiang H., Zheng Y. G.
(англ.)
// Medicinal research reviews. — 2012. — Vol. 32,
no. 4
. — P. 815—867. —
doi
:
. —
.
[
]
Lu J., Kong X., Luo C., Li K. K.
(англ.)
// Medicinal research reviews. — 2013. — Vol. 33,
no. 4
. — P. 790—822. —
doi
:
. —
.
[
]
Suzuki T., Ozasa H., Itoh Y., Zhan P., Sawada H., Mino K., Walport L., Ohkubo R., Kawamura A., Yonezawa M., Tsukada Y., Tumber A., Nakagawa H., Hasegawa M., Sasaki R., Mizukami T., Schofield C. J., Miyata N.
(англ.)
// Journal of medicinal chemistry. — 2013. — Vol. 56,
no. 18
. — P. 7222—7231. —
doi
:
. —
.
[
]
Miranda T. B., Cortez C. C., Yoo C. B., Liang G., Abe M., Kelly T. K., Marquez V. E., Jones P. A.
(англ.)
// Molecular cancer therapeutics. — 2009. — Vol. 8,
no. 6
. — P. 1579—1588. —
doi
:
. —
.
[
]
Barrett S. D., Bridges A. J., Dudley D. T., Saltiel A. R., Fergus J. H., Flamme C. M., Delaney A. M., Kaufman M., LePage S., Leopold W. R., Przybranowski S. A., Sebolt-Leopold J., Van Becelaere K., Doherty A. M., Kennedy R. M., Marston D., Howard W. A. Jr., Smith Y., Warmus J. S., Tecle H.
(англ.)
// Bioorganic & medicinal chemistry letters. — 2008. — Vol. 18,
no. 24
. — P. 6501—6504. —
doi
:
. —
.
[
]
Akinleye A., Furqan M., Mukhi N., Ravella P., Liu D.
(англ.)
// Journal of hematology & oncology. — 2013. — Vol. 6. — P. 27. —
doi
:
. —
.
[
]
Kim, Y., Jeong, J. & Choi, D. (2020). Small-molecule-mediated reprogramming: a silver lining for regenerative medicine. Exp Mol Med.
Xie Xin
,
Fu Yanbin
,
Liu Jian.
(англ.)
// Current Opinion in Genetics & Development. — 2017. — October (
vol. 46
). —
P. 104—113
. —
ISSN
. —
doi
:
.
[
]
Zhao Y.
,
Zhao T.
,
Guan J.
,
Zhang X.
,
Fu Y.
,
Ye J.
,
Zhu J.
,
Meng G.
,
Ge J.
,
Yang S.
,
Cheng L.
,
Du Y.
,
Zhao C.
,
Wang T.
,
Su L.
,
Yang W.
,
Deng H.
(англ.)
// Cell. — 2015. — Vol. 163,
no. 7
. — P. 1678—1691. —
doi
:
. —
.
[
]
Xiang Li et al., Zhen Chai, Hongkui Deng (2017).
. Cell Stem Cell
doi
:
Guan, J., Wang, G., Wang, J., Zhang, Z., Fu, Y., Cheng, L., … & Deng, H. (2022).
от 12 мая 2022 на
Wayback Machine
. Nature, 1-7.
PMID
doi
:
Gan, T., Fan, L., Zhao, L., Misra, M., Liu, M., Zhang, M., & Su, Y. (2021). JNK Signaling in Drosophila aging and longevity. International Journal of Molecular Sciences, 22(17), 9649.
PMID
PMC
doi
:
(неопр.)
. Дата обращения: 5 мая 2015. Архивировано из
14 сентября 2015 года.
Hirata N.
,
Nakagawa M.
,
Fujibayashi Y.
,
Yamauchi K.
,
Murata A.
,
Minami I.
,
Tomioka M.
,
Kondo T.
,
Kuo T. F.
,
Endo H.
,
Inoue H.
,
Sato S.
,
Ando S.
,
Kawazoe Y.
,
Aiba K.
,
Nagata K.
,
Kawase E.
,
Chang Y. T.
,
Suemori H.
,
Eto K.
,
Nakauchi H.
,
Yamanaka S.
,
Nakatsuji N.
,
Ueda K.
,
Uesugi M.
(англ.)
// Cell reports. — 2014. — Vol. 6,
no. 6
. — P. 1165—1174. —
doi
:
. —
.
[
]
Seung-Ju Cho, So-Yeon Kim, Soon-Jung Park, Naree Song, Haw-Young Kwon, Nam-Young Kang, Sung-Hwan Moon, Young-Tae Chang, and Hyuk-Jin Cha (2016).
от 7 июня 2019 на
Wayback Machine
. ACS Cent. Sci., Article ASAP
doi
:
(неопр.)
. Дата обращения: 21 января 2017.
20 января 2017 года.
Mao, D., Ando, S., Sato, S.-i., et al. (2017).
от 2 февраля 2017 на
Wayback Machine
, Angewandte Chemie International Edition
doi
:
Kuang, Y., Miki, K., Parr, C. J., Hayashi, K., Takei, I., Li, J., … & Saito, H. (2017).
. Cell Chemical Biology.
doi
:
Tanosaki, S., Tohyama, S., Fujita, J., Someya, S., Hishiki, T., Matsuura, T., ... & Fukuda, K. (2020). Fatty acid synthesis is indispensable for survival of human pluripotent stem cells. Iscience, 23(9), 101535.
PMID
PMC
doi
:
Nakashima, Y., Miyagi-Shiohira, C., Noguchi, H., & Omasa, T. (2018). Atorvastatin inhibits the HIF1α-PPAR axis, which is essential for maintaining the function of human induced pluripotent stem cells. Molecular Therapy, 26(7), 1715-1734.
PMID
PMC
doi
:
Burkert, K., Taheri, H., Hamad, S. et al. (2021). Salicylic diamines selectively eliminate residual undifferentiated cells from pluripotent stem cell-derived cardiomyocyte preparations. Sci Rep 11, 2391
PMC
doi
:
Takahashi K.
,
Yamanaka S.
(англ.)
// Development (Cambridge, England). — 2013. — Vol. 140,
no. 12
. — P. 2457—2461. —
doi
:
. —
.
[
]
Svendsen C. N.
(англ.)
// Human molecular genetics. — 2013. — Vol. 22,
no. R1
. — P. 32—38. —
doi
:
. —
.
[
]
Madrid M.
,
Sumen C.
,
Aivio S.
,
Saklayen N.
(англ.)
// Current Protocols. — 2021. — March (
vol. 1
,
no. 3
). —
P. e88—88
. —
doi
:
. —
.
[
]
Glaser R.
,
Rice J.
,
Speicher C. E.
,
Stout J. C.
,
Kiecolt-Glaser J. K.
(англ.)
// Behavioral Neuroscience. — 1986. — October (
vol. 100
,
no. 5
). —
P. 675—678
. —
doi
:
. —
.
[
]
Pareja-Galeano H.
,
Sanchis-Gomar F.
,
Pérez L. M.
,
Emanuele E.
,
Lucia A.
,
Gálvez B. G.
,
Gallardo M. E.
(англ.)
// Ageing research reviews. — 2016. — Vol. 27. — P. 37—41. —
doi
:
. —
.
[
]
Литература
Медведев С. П. и др.
Индуцированные плюрипотентные стволовые клетки. — 2-е изд. — Новосибирск: Издательство СО РАН, 2014. — 376 с. —
ISBN 978-5-7692-1384-7
.
Knyazer, A., Bunu, G., Toren, D., Mracica, T. B., Segev, Y., Wolfson, M., … & Fraifeld, V. E. (2021). Small molecules for cell reprogramming: a systems biology analysis. Aging (Albany NY), 13(24), 25739.
PMID
PMC
doi
:
Morris S. A. (2019),
. Nature. 574(7778). News & Views
doi
:
История репрограммирования клеток
Cai, Y., Belmonte, J. C. I., Qu, J., Liu, G. H., & Zhang, W. (2022). Opening up the black box of human cell plasticity. The Innovation, 3(5), 100276.
PMID
PMC
doi
:
Qin, H., Zhao, A. & Fu, X. (2017).
Cell. Mol. Life Sci. 1-23.
doi
:
методов перепрограммирования и анализа полученных ИПСК
Luni, C., Gagliano, O., & Elvassore, N. (2022). Derivation and Differentiation of Human Pluripotent Stem Cells in Microfluidic Devices. Annual Review of Biomedical Engineering, 24, 231—248.
PMID
doi
:
.
Testa, G., Di Benedetto, G., & Passaro, F. (2021). Advanced Technologies to Target Cardiac Cell Fate Plasticity for Heart Regeneration. International Journal of Molecular Sciences, 22(17), 9517.
doi
:
Aguirre, M., Escobar, M., Forero Amézquita, S., Cubillos, D., Rincón, C., Vanegas, P., ... & Celis, L. G. (2023). Application of the Yamanaka Transcription Factors Oct4, Sox2, Klf4, and c-Myc from the Laboratory to the Clinic. Genes, 14(9), 1697.
doi
: